PHI X 174
| Вирус Escherichia φx174 | |
|---|---|
| |
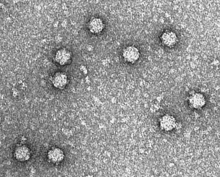
| Электронная микрофотография фага φx174 | |
| Классификация вирусов | |
| (не вмешательство): | Вирус |
| Область : | Моноднавирия |
| Королевство: | Sangervirae |
| Филум: | Фиксвирикота |
| Сорт: | Малграндавирицеты |
| Заказ: | Пятый |
| Семья: | Microviridae |
| Род: | Вирус Синшаймера |
| Разновидность: | Вирус Escherichia φx174
|

Бактериофаг Phi x 174 (или φx174 ) ) , представляет собой одноцепочечный вирус ДНК ( SSDNA который заражает Escherichia coli . Этот вирус был выделен в 1935 году Николя Бульгаковым [ 1 ] В Феликса Д'Эрелле лаборатории в Институте Пастера из образцов, собранных в парижской канализации. Его характеристика и изучение механизма репликации были выполнены с 1950 -х годов. Это был первый геном на основе ДНК , который был секвенирован. Эта работа была завершена Фредом Сэнгером и его командой в 1977 году. [ 2 ] В 1962 году Уолтер Фирс и Роберт Синшаймер уже продемонстрировали физическую, ковалентно закрытую кругуральность ДНК φx174. [ 3 ] Победитель Нобелевской премии Артур Корнберг использовал φx174 в качестве модели, чтобы сначала доказать, что ДНК, синтезированная в пробирке с помощью очищенных ферментов, может вызвать все признаки естественного вируса, открывая эпоху синтетической биологии . [ 4 ] [ 5 ] В 1972–1974 годах Джерард Хервиц , Сью Уикнер и Рид Викнер с соавтором идентифицировали гены, необходимые для производства ферментов для катализации преобразования однократной трансляции вируса в двойную репликативную форму. [ 6 ] сообщила В 2003 году группа Крейга Вентера , что геном φx174 был первым, кто был полностью собран in vitro из синтезированных олигонуклеотидов. [ 7 ] Частица вируса φx174 также была успешно собрана in vitro . [ 8 ] В 2012 году было показано, как его сильно перекрывающийся геном может быть полностью декомпрессирован и при этом оставаться функциональными. [ 9 ]
Геном
[ редактировать ]
Этот бактериофаг имеет геном одноцепочечной ДНК [+] Циркулярный диапазон 5386 нуклеотидов . [ 10 ] Геном GC-контент составляет 44%, а 95% нуклеотидов принадлежат к кодирующим генам. Из -за базовой базовой паттерны генома он используется в качестве контрольной ДНК для секвенсоров Illumina. [ Цитация необходима ]
Гены
[ редактировать ]Φx174 кодирует 11 генов, названных последовательными буквами алфавита в порядке, который они были обнаружены, за исключением*, который является альтернативным начальным кодоном в больших генах. Считается, что только гены a* и k являются неосведомленными, хотя есть некоторые сомнения в*, потому что его начальный кодон может быть изменен на ATT, но не любую другую последовательность. [ 11 ] Теперь известно, что ATT по -прежнему способен продуцировать белок [ 12 ] Внутри кишечной палочки и, следовательно, этот ген может быть важным.
Первая половина генома φx174 отличается высоким уровнем перекрытия генов [ 13 ] с восемью из 11 генов, перекрывающихся как минимум одним нуклеотидом. [ 2 ] Было показано, что эти совпадения не являются необходимыми [ 9 ] Хотя рефакторированный фаг со всеми удаленными генами перекрывался, снизился приспособленность от дикого типа. [ 14 ]
Phage φx174 был использован для попытки установить отсутствие не обнаруженной генетической информации с помощью подхода «доказательства путем синтеза». [ 15 ]
Транскриптом
[ редактировать ]В 2020 году был сгенерирован транскриптом φx174. [ 16 ] Примечательные особенности транскриптома φx174 представляют собой серию из четырех относительно слабых промоторов последовательно с четырьмя Rh-независимыми (внутренними) терминаторами и одним Rho-зависимым терминатором. [ Цитация необходима ]
Белки
[ редактировать ]Φx174 кодирует 11 белков .
| Белок | Копии | Функция [ 17 ] |
|---|---|---|
| А | — | Nicks RF ДНК, чтобы инициировать репликацию Rolling Circle ; Лигат кончики линейной фаговой ДНК с образованием одноцепочечной круглой ДНК |
| A* | — | Ингибирует репликацию ДНК клеток -хозяев; блокируют суперинффицируют фаг; не обязательно |
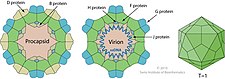
| Беременный | 60 в Прокапсид | Внутренний белок лесов, участвующий в сборке ProCAPSID |
| В | — | Упаковка ДНК |
| Дюймовый | 240 в ProcapsId | Белок внешнего леса, участвуя в сборке ProCAPSID |
| И | — | клеток -хозяина Лизис |
| Фон | 60 в вирионе | Основной капсидный белок |
| Глин | 60 в вирионе | Основной белок шипа |
| ЧАС | 12 в вирионе | Пилотный белок ДНК (или незначительный белок всплеска) |
| Дж | 60 в вирионе | Связывается с новой одноцепочечной фаж-ДНК; сопровождает фаговую ДНК в ProCAPSID |
| K | — | Оптимизирует размер взрыва; не обязательно |
Протеом
[ редактировать ]Недавно сообщалось о идентификации всех белков φx174 с использованием масс -спектрометрии. [ 14 ]
Инфекционный цикл
[ редактировать ]Инфекция начинается, когда G -белок связывается с липополисахаридами на поверхности клетки бактериальной клетки -хозяина. H белок (или пилот -белок ДНК) вирусный геном через бактериальную мембрану E.coli бактерий [ 18 ] скорее всего через прогнозируемую спираль трансмембранного домена . [ 19 ] Тем не менее, стало очевидно, что H белок является многофункциональным белком. [ 20 ] Это единственный вирусный капсидный белок φx174, которым не хватает кристаллической структуры по нескольким причинам. Он имеет низкое ароматическое содержание и высокое содержание глицина , что делает структуру белка очень гибкой, а также, кроме того, отдельные атомы водорода (группа R для глицинов) трудно обнаружить при кристаллографии белка. Кроме того, H белок индуцирует лизис бактериального хозяина при высоких концентрациях, поскольку предсказанная N-концевая трансмембранная спираль легко проталкивает отверстия через бактериальную стенку. По биоинформатике этот белок содержит четыре прогнозируемых домена с спиральной катушкой , которые имеют значительную гомологию для известных факторов транскрипции. Кроме того, было установлено, что белок de novo H был необходим для оптимального синтеза других вирусных белков. [ 21 ] Мутации в белках, которые предотвращают включение вируса, могут быть преодолены, когда поставляются избыточные количества белка B, внутреннего белка для каркасов. [ Цитация необходима ]
ДНК выбрасывается через гидрофильный канал в 5-кратной вершине. [ 22 ] Понятно, что H белок находится в этой области, но экспериментальные данные не подтвердили его точное местоположение. Оказавшись внутри бактерии -хозяина, репликация генома [+] ssDNA протекает посредством промежуточного звена ДНК отрицательного смысла . Это делается как суперклеки генома фага, а вторичная структура, образованная таким суперклейным, привлекает примасому белкового комплекса. Это транслоцирует один раз вокруг генома и синтезирует [ -] ssDNA из положительного исходного генома. [+] геномы SSDNA для упаковки в вирусы создаются из этого механизмом катящегося круга. Это механизм, с помощью которого двойная мельчайшая суперсшитая геном порезан положительной цепью с помощью вируса, кодируемого белком, также притягивая бактериальную ДНК-полимеразу (DNAP) в место расщепления. DNAP использует негативную цепь в качестве шаблона, чтобы сделать ДНК положительного смысла. Когда он транслоцируется вокруг генома, он вытесняет внешнюю цепь уже синтезированной ДНК, которая сразу же покрывается белками SSBP . Белок A расщепляет полный геном каждый раз, когда он распознает последовательность происхождения. [ Цитация необходима ]
Поскольку D -белок является наиболее распространенным геном транскрипта, он является наиболее распространенным белком в вирусной прокапсиде. Точно так же транскрипты генов для F, J и G более распространены, чем для H, поскольку стехиометрия для этих структурных белков составляет 5: 5: 5: 1. Первомасомы представляют собой белковые комплексы, которые прикрепляют/связывают фермент -геликазу на шаблоне. Primosomes дает РНК праймеры для синтеза ДНК для цепей. [ Цитация необходима ]
Скорость мутации
[ редактировать ]Скорость мутации PHIX174 оценивается как 1,0 x 10 -6 Замена на базу на раунд копирования, значение, которое согласуется с правилом Дрейка (0,003 мутации на геном на раунд копирования в микроорганизмах на основе ДНК). [ 23 ]
Рекомбинация
[ редактировать ]PHIX174 способен пройти генетическую рекомбинацию . Основываясь на частотах рекомбинации, полученных в генетических крестах, была построена генетическая карта. [ 24 ] Рекомбинация в PHI X174 связана с высокой негативной интерференцией, то есть положительной корреляцией (негативное помехи) рекомбинационных событий (см. Вмешательство Википедии кроссовер ). [ 24 ]
Филогенетика и разнообразие
[ редактировать ]Φx174 тесно связан с другими Microviridae , особенно фагом NC (например, NC1, NC7, NC11, NC16, NC37, NC5, NC41, NC56, NC51 и т. Д.) И более дистанционно связаны с G4-подобными и даже более дистанционными связанными к α3-подобному фагу. Rokyta et al. 2006 представил филогенетическое дерево их отношений. [ 25 ]
Использование
[ редактировать ]Экспериментальная эволюция
[ редактировать ]Φx174 использовался в качестве модельного организма во многих экспериментах по эволюции. [ 26 ]
Биотехнология
[ редактировать ]Φx174 регулярно используется в качестве положительного контроля при секвенировании ДНК из -за его относительно небольшого размера генома по сравнению с другими организмами, его относительно сбалансированное содержание нуклеотидов - около 23% г, 22% c, 24% a и 31% t, т.е. 45% G+C и 55% A+T, см. Вступление NC_001422.1 [ 10 ] для его 5386 длинных нуклеотидных последовательности. Инструменты секвенирования Illumina используют φx174 в качестве положительного контроля, [ 27 ] И один запуск секвенирования Illumina может охватить геном φx174 несколько миллионов раз, что делает этот очень вероятный геном в истории. [ Цитация необходима ]
Φx174 также используется для проверки устойчивости оборудования для личного защиты к вирусам кровопролития. [ 28 ]
Φx174 также был модифицирован, чтобы включить пептидный дисплей (фаговый дисплей) из вирусного капсида G -белка. [ 29 ]
Синтетическая биология
[ редактировать ]Геном φx174 был первым фагом, который был клонирован в дрожжах, [ 9 ] который обеспечивает удобный сухой док для модификаций генома. [ 30 ] Φx174 был также первым геномом, который был полностью разместил, с удаленными генов. [ 13 ] Эффект этих изменений привел к значительному снижению прикрепления хозяина, диспрессируемой экспрессии белка и чувствительности тепла. [ 14 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Лаксович, Здравко; Толжан, Карло (20 декабря 2020 г.). «Владимир Сертич: забытый пионер вирусологии и бактериофагов» . Примечания и записи: Журнал Королевского общества истории науки . 74 (4): 567–578. doi : 10.1098/rsnr.2019.0010 . ISSN 0035-9149 . PMC 7653334 . PMID 33177747 .
- ^ Jump up to: а беременный Sanger F, Air GM, Barrell BG, Brown NL, Coulson AR, Fiddes CA, et al. (Февраль 1977 г.). «Нуклеотидная последовательность бактериофага ДНК PHI X174». Природа . 265 (5596): 687–95. Bibcode : 1977natur.265..687s . doi : 10.1038/265687a0 . PMID 870828 . S2CID 4206886 .
- ^ Fiers W, Sinsheimer RL (октябрь 1962 г.). «Структура ДНК бактериофага PHI-X174. III. Ультрацентрифугальные доказательства кольцевой структуры». Журнал молекулярной биологии . 5 (4): 424–34. doi : 10.1016/s0022-2836 (62) 80031-x . PMID 13945085 .
- ^ Национальная библиотека медицины в науке. Артур Корнберг Бумаги. «Создание жизни в пробирке», 1959-1970. связь [ Необходимый источник необходимы ]]
- ^ Гулиан М., Корнберг А., Синшаймер Р.Л. (декабрь 1967 г.). «Ферментативный синтез ДНК, XXIV. Синтез инфекционного фага ДНК Phi-X174» . Труды Национальной академии наук Соединенных Штатов Америки . 58 (6): 2321–8. Bibcode : 1967pnas ... 58.2321g . doi : 10.1073/pnas.58.6.2321 . JSTOR 58720 . PMC 223838 . PMID 4873588 .
- ^ Wickner S, Hurwitz J (октябрь 1974 г.). «Превращение вирусной ДНК PHIX174 в двухцепочечную форму очищенными белками Escherichia coli» . Труды Национальной академии наук Соединенных Штатов Америки . 71 (10): 4120–4. doi : 10.1073/pnas.71.10.4120 . PMC 434340 . PMID 4610569 .
- ^ Смит Хо, Хатчисон К.А., Пфаннкоч С., Вентер Дж.С. (декабрь 2003 г.). «Генерирование синтетического генома с помощью сборки цельного генома: бактериофаг PHIX174 из синтетических олигонуклеотидов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (26): 15440–5. Bibcode : 2003pnas..10015440S . doi : 10.1073/pnas.2237126100 . JSTOR 3149024 . PMC 307586 . PMID 14657399 .
- ^ Шерва Дж., Органтини Л.Дж., Эшли Р.Е., Хафенштейн С.Л., Фейн Б.А. (сентябрь 2011 г.). «In vitro сборка Øx174 ProcapsId из белковых олигомеров внешних каркасов и ранних промежуточных продуктов пентамерной сборки». Журнал молекулярной биологии . 412 (3): 387–96. doi : 10.1016/j.jmb.2011.07.070 . PMID 21840317 .
- ^ Jump up to: а беременный в Джашке П.Р., Либерман Эк, Родригес Дж., Сьерра А., Энди Д (декабрь 2012 г.). «Полностью декомпрессированный синтетический бактериофаг ØX174, собранное и архивируемое в дрожжах» . Вирусология . 434 (2): 278–84. doi : 10.1016/j.virol.2012.09.020 . PMID 23079106 .
- ^ Jump up to: а беременный в Enterobacteria phage phix174 Sensu Lato , полный геном. «Полный геном: вступление NC_001422» , Национальный центр информации о биотехнологии . Получено 30 января 2016 года.
- ^ Baas PD, Liewerink H, Van Teeffelen HA, Van Mansfeld AD, Van Boom JH, Jansz HS (июнь 1987 г.). «Изменение начального кодона ATG белка A бактериофага Phi X174 в кодон ATT дает жизнеспособный фаг, указывающий на то, что белок не является необходимым для воспроизведения PHI X174» . Письма Febs . 218 (1): 119–25. doi : 10.1016/0014-5793 (87) 81030-x . PMID 2954853 . S2CID 24174007 .
- ^ Hecht A, Glasgow J, Jaschke PR, Bawazer LA, Munson MS, Cochran JR, et al. (Апрель 2017). «Измерения инициации перевода от всех 64 кодонов в кишечной палочке» . Исследование нуклеиновых кислот . 45 (7): 3615–3626. doi : 10.1093/nar/gkx070 . PMC 5397182 . PMID 28334756 .
- ^ Jump up to: а беременный Райт, Брэдли В.; Моллой, Марк П.; Джашке, Пол Р. (5 октября 2021 г.). «Перекрывающиеся гены в натуральных и инженерных геномах» . Nature Reviews Genetics . 23 (3): 154–168. doi : 10.1038/s41576-021-00417-w . ISSN 1471-0064 . PMC 8490965 . PMID 34611352 .
- ^ Jump up to: а беременный в Райт Б.В., Руан Дж., Моллой М.П., Джашке П.Р. (ноябрь 2020 г.). «Модуляризация генома выявляет перекрывающуюся топологию генов необходима для эффективного вирусного размножения». ACS Синтетическая биология . 9 (11): 3079–3090. doi : 10.1021/acssynbio.0c00323 . PMID 33044064 . S2CID 222300240 .
- ^ Jaschke PR, Dotson GA, Hung KS, Liu D, Endy D (ноябрь 2019). «Окончательная демонстрация путем синтеза полноты аннотации генома» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (48): 24206–24213. doi : 10.1073/pnas.1905990116 . PMC 6883844 . PMID 31719208 .
- ^ Logel Dy, Jaschke PR (август 2020 г.). «Карта высокого разрешения бактериофага ϕx174» . Вирусология . 547 : 47–56. doi : 10.1016/j.virol.2020.05.008 . PMID 32560904 . S2CID 219459208 .
- ^ Фейн Б.А., Брентлингер К.Л., Берч А.Д., Чен М., Хафенштейн С., Мур Е., Новак К.Р., Учияма А. (2006). "ɸx174 и др. Microviridae , В календере R (ред.). Бактериофафы (2 -е изд.). Нью -Йорк: Оксфордский университет. Нажимать. П. 130. ISBN 978-0195148503 .
- ^ Джазвински С.М., Линдберг А.А., Корнберг А (июль 1975 г.). «Рецептор липополисахаридов для бактериофага PHIX174 и S13». Вирусология . 66 (1): 268–82. doi : 10.1016/0042-6822 (75) 90197-X . PMID 1094681 .
- ^ Tusnády GE, Саймон I (сентябрь 2001 г.). «Сервер прогнологии топологии трансмембранной топологии HMMTOP» . Биоинформатика . 17 (9): 849–50. doi : 10.1093/bioinformatics/17.9.849 . PMID 11590105 .
- ^ Шерва JE, Young LN, Fane BA (март 2011 г.). «Необвисывание функций многофункционального белка: выделение мутанта белка -пилота ДНК, который влияет на морфогенез частиц» . Вирусология . 411 (1): 9–14. doi : 10.1016/j.virol.2010.12.026 . PMID 21227478 .
- ^ Ruboyianes MV, Chen M, Dubrava MS, Cherwa Je, Fane Ba (октябрь 2009 г.). «Экспрессия N-концевых делеционных пилотных белков ДНК ингибирует ранние стадии репликации PHIX174» . Журнал вирусологии . 83 (19): 9952–6. doi : 10.1128/jvi.01077-09 . PMC 2748053 . PMID 19640994 .
- ^ McKenna R, Xia D, Willowmann P, Ilag LL, Krishnaswamy S, Rossmann MG, et al. (Январь 1992). «Атомная структура одноцепочечного ДНК-бактериофага PHI X174 и его функциональные последствия» . Природа . 355 (6356): 137–43. Bibcode : 1992natur.355..137M . doi : 10.1038/355137A0 . PMC 4167681 . PMID 1370343 .
- ^ Cuevas JM, Duffy S, Sanjuán R (октябрь 2009 г.). «Скорость точечной мутации бактериофага PHIX174» . Генетика . 183 (2): 747–9. doi : 10.1534/Genetics.109.106005 . PMC 2766332 . PMID 19652180 .
- ^ Jump up to: а беременный Бенбоу Р.М., Хатчисон К.А., Фабритант Д.Д., Синшаймер Р.Л. (май 1971). «Генетическая карта бактериофага Phix174» . J Virol . 7 (5): 549–58. doi : 10.1128/jvi.7.5.549-558.1971 . PMC 356162 . PMID 16789129 .
- ^ Rokyta DR, Burch CL, Caudle SB, Wichman HA (февраль 2006 г.). «Горизонтальный перенос генов и эволюция геномов колифхажа микровиридов» . Журнал бактериологии . 188 (3): 1134–42. doi : 10.1128/jb.188.3.1134-1142.2006 . PMC 1347346 . PMID 16428417 .
- ^ Wichman HA, Brown CJ (август 2010 г.). «Экспериментальная эволюция вирусов: Microviridae как модельная система» . Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 365 (1552): 2495–501. doi : 10.1098/rstb.2010.0053 . PMC 2935103 . PMID 20643739 .
- ^ «Использование управления PHIX для секвенирования HISEQ®» . Illumina. Архивировано с оригинала 9 января 2019 года . Получено 8 января 2019 года .
- ^ «Ст . wwwn.cdc.gov . Получено 8 февраля 2019 года .
- ^ Christakos KJ, Chapman JA, Fane Ba, Campos SK (январь 2016 г.). «Phixing-It, отображая инородные пептиды на бактериофаге φx174» . Вирусология . 488 : 242–8. doi : 10.1016/j.virol.2015.11.021 . PMC 6191337 . PMID 26655242 .
- ^ Ando H, Lemire S, Pires DP, Lu TK (сентябрь 2015 г.). «Инженерные модульные вирусные каркасы для редактирования бактериальной популяции» . Клеточные системы . 1 (3): 187–196. doi : 10.1016/j.cels.2015.08.013 . PMC 4785837 . PMID 26973885 .
Внешние ссылки
[ редактировать ]- Goodsell D (февраль 2000 г.). «Бактериофаг PHIX174» . Молекула месяца . RCSB-PDB.