Антенна (биология)

Усики ( sg.: антенна ) , иногда называемые «щупальцами», представляют собой парные придатки, используемые для зондирования у членистоногих .
Усики прикрепляются к первым одному или двум сегментам головы членистоногих. Они сильно различаются по форме, но всегда состоят из одного или нескольких сочлененных сегментов. Хотя они обычно являются органами чувств , точная природа того, что они ощущают и как они это ощущают, не одинакова во всех группах. Функции могут включать в себя прикосновение , движение воздуха, тепло, вибрацию (звук) и особенно запах или вкус . [ 1 ] [ 2 ] Усики иногда модифицируются для других целей, таких как спаривание, высиживание, плавание и даже прикрепление членистоногого к субстрату . [ 2 ] У личинок членистоногих усики отличаются от таковых у взрослых особей. Например, у многих ракообразных есть свободноплавающие личинки , которые для плавания используют свои усики. Усики также могут определять местонахождение других членов группы, если насекомое живет в группе, как муравей . Общий предок всех членистоногих, вероятно, имел одну пару одноветвистых (неразветвленных) антенноподобных структур, за которыми следовали одна или несколько пар двуветвистых (имеющих две основные ветви) ногообразных структур, как это наблюдается у некоторых современных ракообразных и ископаемых трилобитов . [ 3 ] За исключением хелицератов и протуранов , у которых их нет, все членистоногие, не относящиеся к ракообразным, имеют одну пару усиков. [ 4 ]
Ракообразные
[ редактировать ]
Ракообразные несут две пары усиков. Пара, прикрепленная к первому сегменту головы, называется первичными усиками или антеннулами . Эта пара обычно одноветвистая, но двуветвистая у крабов, омаров и ремипедов . Пара, прикрепленная ко второму сегменту, называется вторичными усиками или просто усиками . Вторые усики плезиоморфно двуветвистые, но позже у многих видов появились одноветвистые пары. [ 2 ] Вторые усики могут быть значительно редуцированы (например, ремипеды) или явно отсутствовать (например, ракушки ).
Подразделения усиков ракообразных имеют множество названий, включая жгутики (общий термин для насекомых), кольца, статьи и сегменты. Концевые концы усиков ракообразных делятся на две основные категории: сегментированные и жгутиковые. Антенна считается сегментированной, если каждое из колец отделено от окружающих и имеет отдельные мышечные крепления. С другой стороны, жгутиковые усики имеют мышечные прикрепления только вокруг основания, действуя как шарнир для жгутика — гибкой цепочки колец без прикрепления мышц. [ 5 ]
У ракообразных есть несколько примечательных несенсорных способов использования усиков. У многих ракообразных есть подвижная личиночная стадия, называемая науплиусом , которая характеризуется использованием усиков для плавания. Ракушки , сильно модифицированные ракообразные, используют свои усики для прикрепления к камням и другим поверхностям. [ 2 ] Вторые усики роющих Hippoidea и Corystidae имеют щетинки, которые сцепляются, образуя трубку или «трубку», по которой фильтрованная вода течет через жабры. [ 6 ]





| Копепода | изопода | Амфипода | Десятиногие | Десятиногие | Ремипедия | Чиррепедия | |
|---|---|---|---|---|---|---|---|
Первичные усики
|

|

|

|

|
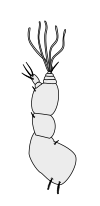
|

|
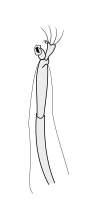
|
Вторичные усики
|

|

|

|
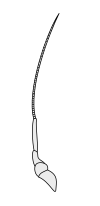
|
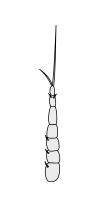
|

|
Насекомые
[ редактировать ]
Некоторые утверждают, что насекомые произошли от доисторических ракообразных и что у них есть вторичные усики, как у ракообразных, но нет первичных усиков. Усики — основные обонятельные сенсоры насекомых. [ 7 ] и, соответственно, хорошо оснащены разнообразными сенсиллами (единственное число: сенсиллы ). Парные, подвижные и сегментированные, они расположены между глазами на лбу. Эмбриологически они представляют собой придатки второго сегмента головы. [ 8 ]
Усики есть у всех насекомых, однако у личиночных форм они могут быть значительно редуцированы. Среди ненасекомых классов Hexapoda у Collembola и Diplura есть антенны, а у Protura их нет. [ 9 ]
Фибриллы усиков играют важную роль в Culex pipiens спаривании . Возведение этих фибрилл считается первым этапом размножения. Эти фибриллы выполняют разные функции у разных полов. Поскольку фибриллы усиков используются самками C. pipiens для поиска хозяев и кормления, самцы C. pipiens используют их для поиска самок. [ нужна ссылка ]
Структура
[ редактировать ]
Три основных сегмента типичной антенны насекомых — это скапус или лопатка (основание), ножка или ножка (стебель) и, наконец, жгутик , который часто состоит из множества единиц, известных как жгутики . [ 10 ] Цветоножка (второй сегмент) содержит орган Джонстона , представляющий собой скопление чувствительных клеток. [ 11 ]
Чешуя прикреплена к гнезду в более или менее кольцеобразной склеротизованной области, называемой торулусом , часто являющейся приподнятой частью головной капсулы насекомого. Розетка закрыта мембраной, в которую встроено основание черешка. Однако антенна не висит на мембране свободно, а поворачивается на жестко подпружиненном выступе от края тора. Тот выступ, на котором вращается антенна, называется усиком . Вся конструкция позволяет насекомому перемещать антенну целиком за счет внутренних мышц, связанных со скапусом. Цветоножка гибко связана с дистальным концом черенка, а его движениями, в свою очередь, можно управлять с помощью мышечных связей между черешком и ножкой. Количество жгутиков может сильно различаться у разных видов насекомых и часто имеет диагностическое значение. [ нужна ссылка ]
Настоящие жгутики соединены мембранной связью , которая обеспечивает движение, хотя жгутик «настоящих» насекомых не имеет внутренних мышц. Однако у некоторых других членистоногих имеются внутренние мышцы по всему жгутику. К таким группам относятся Symphyla , Collembola и Diplura . У многих настоящих насекомых, особенно у более примитивных групп, таких как Thysanura и Blattodea , жгутик частично или полностью состоит из гибко связанной цепочки небольших кольцеобразных колец . Кольца не являются настоящими флагелломерами, и у данного вида насекомых количество колец обычно не так постоянно, как количество жгутиков у большинства видов. [ 10 ]
У многих жуков и у ос апикальные хальцидоидных жгутики образуют булавовидную форму, называемую клавой . Общий термин для сегментов между булавой и основанием усика — жгутик ; Традиционно при описании анатомии жука термин «пучок» относится к сегментам между булавой и скапусом . Однако традиционно при работе с осами за жгутик принимают сегменты между булавой и ножкой. [ 10 ]
, пучок за ножкой довольно сложный Довольно часто у Endopterygota, таких как жуки, мотыльки и перепончатокрылые , и одной из распространенных адаптаций является способность складывать антенну посередине, в месте соединения ножки и жгутика. Это дает эффект типа «сгибания колена», и такую антенну называют коленчатой . Коленообразные усики обычны у жесткокрылых и перепончатокрылых. Они важны для насекомых, таких как муравьи, которые следуют по запаховым следам, для пчел и ос, которым необходимо «нюхать» цветы, которые они посещают, а также для жуков, таких как Scarabaeidae и Curculionidae , которым необходимо откидывать усики, когда они складываются в целях самозащиты. все их конечности в оборонительной позиции. [ нужна ссылка ]
Поскольку канатик не имеет собственных мышц, он обычно должен двигаться как единое целое, несмотря на то, что он сочленен. Однако некоторые канатики сложны и очень подвижны. Например, у Scarabaeidae есть пластинчатые усики, которые можно плотно сложить в целях безопасности или раскрыть открыто для обнаружения запахов или феромонов . Насекомое управляет такими действиями за счет изменения кровяного давления, за счет чего оно использует эластичность стенок и мембран канатиков, которые, по сути, обладают эрекцией. [ 12 ]
В группах с более однородными усиками (например: многоножки ) все членики называются антенномерами . Некоторые группы имеют простую или различным образом видоизмененную апикальную или субапикальную щетину, называемую аристой (она может быть особенно хорошо развита у различных Diptera ). [ 13 ]
Функции
[ редактировать ]
Обонятельные рецепторы на антеннах связываются со свободно плавающими молекулами, такими как водяной пар , и запахами, включая феромоны . Нейроны потенциалы , обладающие этими рецепторами, сигнализируют об этом связывании, посылая действия своим аксонам в антеннальную долю мозга . по Отсюда нейроны в долях усиков соединяются с грибовидными телами , которые распознают запах. Сумма электрических потенциалов усиков к данному запаху может быть измерена с помощью электроантеннограммы . [ 14 ]
У бабочки-монарха усики необходимы для правильной ориентации по солнечному компасу с компенсацией времени во время миграции. У монархов существуют антенные часы, и они, вероятно, обеспечивают основной механизм синхронизации для ориентации по солнечному компасу. [ 15 ]
У африканского хлопкового листовертки усики выполняют важную функцию сигнализации ухаживания. В частности, усики необходимы самцам, чтобы ответить на брачный зов самки. Хотя самкам для спаривания усики не требуются, спаривание самки без усиков было ненормальным. [ 16 ]
У ромбовидной моли усики служат для сбора информации о вкусе и запахе растения-хозяина. После того, как желаемый вкус и запах будут определены, самка бабочки откладывает яйца на растение. [ 17 ] Гигантские бабочки- парусники также полагаются на чувствительность антенн к летучим соединениям при идентификации растений-хозяев. Было обнаружено, что самки на самом деле более чувствительны к распознаванию антенн, скорее всего, потому, что они отвечают за откладку яиц на правильном растении. [ 18 ]
У сумеречного бражника ( Manduca sexta ) усики помогают стабилизировать полет. Подобно жужжальцам двукрылых насекомых, усики передают силы Кориолиса через орган Джонстона, которые затем можно использовать для корректирующего поведения. Серия исследований стабильности полета при слабом освещении, в которых бабочки с ампутированными жгутиками возле ножки, показали значительно более низкую стабильность полета по сравнению с бабочками с неповрежденными усиками. [ 19 ] Чтобы определить, могут ли существовать другие сенсорные входы от антенн, второй группе бабочек ампутировали усики, а затем снова прикрепили их, прежде чем пройти тестирование в том же исследовании стабильности. Эти мотыльки показали немного худшие характеристики по сравнению с интактными бабочками, что указывает на то, что, возможно, для стабилизации полета используются другие сенсорные сигналы. Повторная ампутация усиков привела к резкому снижению устойчивости полета до уровня первой ампутированной группы. [ нужна ссылка ]
Ссылки
[ редактировать ]- ^ Чепмен, РФ (1998). Насекомые: структура и функции (4-е изд.). Издательство Кембриджского университета. стр. 8–11 . ISBN 978-0521570480 .
- ^ Перейти обратно: а б с д Боксшолл, Джефф; Жауме, Д. (2013). Функциональная морфология и разнообразие: антеннулы и усики ракообразных . Издательство Оксфордского университета. стр. 199–236. doi : 10.1093/acprof:osobl/9780195398038.003.0007 .
- ^ Форти, Ричард А.; Томас, Ричард Х. (1998). Отношения членистоногих: филогенетический анализ (1-е изд.). Ассоциация систематиков. п. 117. ИСБН 978-94-011-4904-4 .
- ^ Коттон, Тревор Дж.; Брэдди, Саймон Дж. (2004). «Филогения арахноморфных членистоногих и происхождение Chelicerata». Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 94 (3): 169–193. дои : 10.1017/S0263593300000596 . S2CID 54841296 .
- ^ Боксхолл, Джорджия (2004). «Эволюция конечностей членистоногих». Биологические обзоры Кембриджского философского общества . 79 (2): 253–300. дои : 10.1017/s1464793103006274 . ПМИД 15191225 . S2CID 25339728 .
- ^ Люке, Дж.; Фельдманн, Р.М.; Вернигора, О.; Швейцер, CE; Кэмерон, CB; Керр, штат Калифорния; Вега, ФДж; Дуке, А.; Странно, М.; Палмер, Арканзас; Харамилло, К. (2019). «Исключительная сохранность морских членистоногих среднего мела и эволюция новых форм посредством гетерохронии» . Достижения науки . 5 (4): eaav3875. Бибкод : 2019SciA....5.3875L . дои : 10.1126/sciadv.aav3875 . ПМК 6482010 . ПМИД 31032408 .
- ^ Дарби, Джин (1958). Что такое бабочка? . Чикаго: Бенефик Пресс . п. 8. ОСЛК 1391997 .
- ^ Галлан, Пенни Дж.; Крэнстон, Питер С. (2005). Насекомые: Очерк энтомологии (3-е изд.). Оксфорд, Великобритания: Blackwell Publishing . п. 38 . ISBN 978-1-4051-1113-3 .
- ^ Чепмен, Реджинальд Фредерик (1998). Насекомые: структура и функции (4-е изд.). Нью-Йорк: Издательство Кембриджского университета . п. 8 . ISBN 978-0-521-57890-5 .
- ^ Перейти обратно: а б с Томас А. Кейл (1999). «Морфология и развитие периферических органов обоняния». В Ханссоне, Билл С. (ред.). Обоняние насекомых (1-е изд.). Спрингер . стр. 5–48. ISBN 978-3-540-65034-8 .
- ^ Насекомые: Очерк энтомологии, четвертое издание
- ^ Пасс, Гюнтер. Анатомия и ультраструктура антеннальных органов кровообращения жука-хук Melolontha melolontha L. (Coleoptera, Scarabaeidae) Зооморфология 01/1980; 96(1):77-89. два : 10.1007/BF00310078
- ^ Лоуренс, Элеонора, изд. (2005). Словарь биологических терминов Хендерсона (13-е изд.). Образование Пирсона . п. 51 . ISBN 978-0-13-127384-9 .
- ^ «Электроантеннография (ЭАГ)» . Университет Георга Августа Геттингена . Проверено 27 марта 2010 г.
- ^ Мерлин, Кристина; Гегер, Роберт Дж.; Репперт, Стивен М. (сентябрь 2009 г.). «Усиковые циркадные часы координируют ориентацию по солнечному компасу у мигрирующих бабочек-монархов» . Наука . 325 (5948): 1700–1704. Бибкод : 2009Sci...325.1700M . дои : 10.1126/science.1176221 . ПМЦ 2754321 . ПМИД 19779201 .
- ^ Эллис, Пегги; Бримакомб, Линда (1980). «Брачное поведение египетской хлопковой листовертки Spodopteralittoralis (Boisd.)». Поведение животных . 28 (4): 1239–1248. дои : 10.1016/s0003-3472(80)80112-6 . S2CID 53188823 .
- ^ Юстус, штат Калифорния; Митчелл, БК (ноябрь 1996 г.). «Выбор места откладки яиц бабочкой Plutella xylostella (L.) (Lepidoptera: Plutellidae)». Журнал поведения насекомых . 9 (6): 887–898. Бибкод : 1996JIBeh...9..887J . дои : 10.1007/BF02208976 . S2CID 28455636 .
- ^ М.И.), Скрибер, Дж.М. (Университет штата Мичиган, Ист-Лансинг; Р.В., Доуэлл (1991). «Пригодность растения-хозяина и проверка гипотезы пищевой специализации с использованием Papilio cresphontes (Lepidoptera: Papilionidae)» . Энтомолог Великих озер (США) .ИССН 0090-0222 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сане, Санджай П.; Дьедонне, Александр; Уиллис, Марк А.; Дэниел, Томас Л. (9 февраля 2007 г.). «Антеннальные механосенсоры обеспечивают управление полетом бабочек». Наука . 315 (5813): 863–866. Бибкод : 2007Sci...315..863S . CiteSeerX 10.1.1.205.7318 . дои : 10.1126/science.1133598 . ISSN 0036-8075 . ПМИД 17290001 . S2CID 2429129 .
| Базы данных органов управления : Национальные |
|---|