Антенна (биология)

Антенны ( SG . : Antenna ), иногда называемая «щупальцами», являются парными придатками, используемыми для зондирования в членистоногих .
Антенны связаны с первым одним или двумя сегментами головы членистоногих. Они широко различаются по форме, но всегда сделаны из одного или нескольких соединенных сегментов. Хотя они обычно являются сенсорными органами , точная природа того, что они чувствуют и как они чувствуют, это не одно и то же во всех группах. Функции могут по -разному включать в себя чувствительный прикосновение , движение воздуха, тепло, вибрацию (звук) и особенно запах или вкус . [ 1 ] [ 2 ] Антенны иногда модифицируются для других целей, таких как спаривание, задумчивость, плавание и даже привязка членистоногих на подложку . [ 2 ] Членистоногие из личинок имеют антенны, которые отличаются от антенных от взрослого. Например, у многих ракообразных есть личинки , которые используют свои антенны для плавания. Антенны также могут найти других членов группы, если насекомое живет в группе, как муравей . Общий предок всех членистоногих, вероятно, имел одну пару однорамных (неразветвленных) антенных структур, за которыми следуют одна или несколько пар бириамных (имеющих две основные ветви), похожие на ноги, как видно у некоторых современных ракообразных и ископаемых трилобитов . [ 3 ] За исключением хелицератов и протранцев , которые не имеют ни одного, все некстачевые членистоногих имеют одну пару антенн. [ 4 ]
Ракообразные
[ редактировать ]
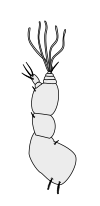
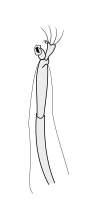
Раковые ракообразные несут две пары антенн. Пара, прикрепленная к первому сегменту головы, называется первичными антеннами или антеннами . Эта пара, как правило, однорамна, но является бирамным в крабах, омарах и переосмыслениях . Пара, прикрепленная ко второму сегменту, называются вторичными антеннами или просто антеннами . Вторые антенны являются плезиоморфическими , но многие виды позже развили однорамные пары. [ 2 ] Вторые антенны могут быть значительно снижены (например, переоборудование) или, по -видимому, отсутствуют (например, саралы ).
В подразделениях ракообразных антенн имеет много имен, в том числе жгутиковые, общие термины с насекомыми), аннули, статьи и сегменты. Терминальные концы ракообразных антенн имеют две основные категоризации: сегментированные и жгутиковые. Антенна считается сегментированной, если каждая из аннули отделена от окружающих и имеет отдельные мышечные прикрепления. Плаговые антенны, с другой стороны, имеют мышечные прикрепления только к основанию, действуя как шарнир для жгутика - гибкая цепочка аннули без прикрепления мышц. [ 5 ]
Есть несколько заметных незенсорных применений антенн у ракообразных. Многие ракообразные имеют подвижную личиночную стадию под названием Nauplius , которая характеризуется использованием антенн для плавания. Barnacles , высоко модифицированное ракообразные, используют свои антенны для прикрепления к камням и другим поверхностям. [ 2 ] Вторые антенны в Burrroving Hippoidea и Corystidae имеют щетинки, которые переключаются, образуя трубку или «сноркелу», которая прокинула воду над жабрами. [ 6 ]





| МОПЕПОДА | Изопода | Амфипода | Decapoda | Decapoda | Remipedia | Cirretia | |
|---|---|---|---|---|---|---|---|
Первичные антенны
|

|

|

|

|
|

|
|
Вторичные антенны
|

|

|

|
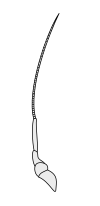
|
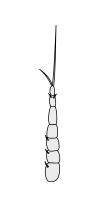
|

|
Насекомые
[ редактировать ]
Некоторые утверждают, что насекомые развивались из доисторических ракообразных, и у них есть вторичные антенны, такие как ракообразные, но не первичные антенны. Антенны являются основными обонятельными датчиками насекомых [ 7 ] и соответственно хорошо оборудованы с широким спектром сенсиллы ( Sensillum ). Парные, мобильные и сегментированные, они расположены между глазами на лбу. Эмбриологически они представляют придатки второго сегмента головы. [ 8 ]
Все насекомые имеют антенны, однако они могут быть значительно уменьшены в личиночных формах. Среди неинзаконных классов гексаподы , как Коллембола , так и диплора имеют антенну, а протура нет. [ 9 ]
Антенные фибриллы играют важную роль в практике спаривания Culex Pipiens . Эффекция этих фибрилсов считается первой стадией в воспроизведении. Эти фибриллы служат разным функциям во всем полое. Поскольку антенные фибриллы используются самками C. Pipiens для размещения хозяев для питания, мужские Pipiens используют их для размещения женщин -партнеров. [ Цитация необходима ]
Структура
[ редактировать ]
Три основных сегмента типичной антенны насекомых - это Scape или Scapus (основание), цветоножка или педицеллус (STEM) и, наконец, жгутичок , который часто включает в себя множество единиц, известных как флагелломеры . [ 10 ] Песедовая (второй сегмент) содержит орган Джонстона , который представляет собой набор сенсорных ячеек. [ 11 ]
Scape монтируется в гнездо в более или менее склеротизированной области в форме кольца, называемой Torulus , часто поднятой части капсулы головы насекомых. Гребень закрыт мембраной, в которую устанавливается основание от Scape. Тем не менее, антенна не висит на мембране, а поворачивается на жестко выросшей проекции от обода турулуса. Эта проекция, на которой антенна называется антеннифером . Вся структура позволяет насекомому перемещать антенну в целом, нанося внутренние мышцы, соединенные с Scape. Цибковая цветочка гибко соединена с дистальным концом Scape, и ее движения, в свою очередь, могут контролироваться мышечными соединениями между Scape и Pedicel. Количество жгутиков может сильно различаться между видами насекомых и часто имеет диагностическое значение. [ Цитация необходима ]
Истинные флагелломеры связаны мембранной связью , которая позволяет двигаться, хотя жгутичок «истинных» насекомых не имеет каких -либо внутренних мышц. Однако некоторые другие члпания имеют внутренние мышцы по всему жгутике. Такие группы включают Симфила , Коллемболу и Диплуру . У многих истинных насекомых, особенно более примитивных групп, таких как тисанура и блаттодиа , флагелм частично или полностью состоит из гибко соединенной строки маленьких кольцевых аннули . Аннулы не являются настоящими жгутиками, а у данных видов насекомых число аннули, как правило, не так последовательно, как количество жгутовщиков у большинства видов. [ 10 ]
Во многих жуках и в хальцидоидных ос апикальных называемую жгутиков образуют клубную форму, клаву . Коллективный термин для сегментов между клубом и антенной базой является Funicle ; Традиционно при описании анатомии жуков термин «funicle» относится к сегментам между клубом и Scape . Тем не менее, традиционно при работе над осами принимается, чтобы составлять сегменты между клубом и цветоночной кости. [ 10 ]
Вполне часто Funicle за рамки цветоножки в эндоптериготе довольно сложна , таких как жуки, мотыльки и гименоптера , и одной общей адаптацией является способность сложить антенну в середину, в суставе между цветоночной и жгутией. Это дает эффект, как «изгиб колена», и такая антенна, как говорят, является геникулентной . Геникулятные антенны распространены в Coleoptera и Hymenoptera. Они важны для насекомых, таких как муравьи, которые следуют за ароматическими тропами, для пчел и ошей, которым необходимо «нюхать» цветы, которые они посещают, и для жуков, таких как Scrabaeidae и Curculionidae , которые должны сложить свои антенны, когда они самопомощивыво складываются Все их конечности в оборонительных отношениях. [ Цитация необходима ]
Поскольку Funicle не имеет внутренних мышц, он обычно должен двигаться как единое целое, несмотря на то, что их сформулировали. Тем не менее, некоторые любые сложны и очень мобильны. Например, Scarabaeidae имеют пластинчатые антенны, которые можно плотно сложить для безопасности или разбросано для обнаружения запахов или феромонов . Насекомое управляет такими действиями путем изменений в артериальном давлении, с помощью которых он использует эластичность в стенах и мембранах в любых веществах, которые действительно являются эректильными. [ 12 ]
В группах с более однородными антеннами (например: Millipedes ) все сегменты называются антенномерами . Некоторые группы имеют простую или по-разному модифицированной апикальной или субапикальной щетины, называемой Arista (это может быть особенно хорошо разработано в различных Diptera ). [ 13 ]
Функции
[ редактировать ]
Обонятельные рецепторы на антеннах связываются с свободными молекулами, такими как водяной пара , и запахи, включая феромоны . Нейроны , которые обладают этими рецепторами , сигнализируют об этом связывании, отправляя потенциалы действия вниз по своим аксонам в антеннальную долю в мозге . Оттуда нейроны в антеннальных долях соединяются с грибными телами , которые идентифицируют запах. Сумма электрических потенциалов антенн до данного запаха может быть измерена с использованием электроантрограммы . [ 14 ]
В бабочке монарха во времени антенны необходимы для правильной ориентации на солнечный компас во время миграции. Угодные часы существуют в монархах, и они, вероятно, обеспечат основной механизм времени для ориентации на солнцезащитный компас. [ 15 ]
У африканского хлопкового листового червя антенны имеют важную функцию в сигнализации ухаживания. В частности, антенны необходимы для мужчин ответить на звонок для женского спаривания. Хотя женщины не требуют антенн для спаривания, спаривание, которое возникло от самок без антенн, было ненормальным. [ 16 ]
В Diamondback Moth антенны служат для сбора информации о вкусе и запахе растения. После того, как желаемый вкус и запах были идентифицированы, самка мотылька будет положить свои яйца на растение. [ 17 ] Гигантские бабочки с ласточками также полагаются на чувствительность антенны к летучим соединениям для идентификации растений -хозяев. Было обнаружено, что женщины на самом деле более отзывчивы с их зондированием антенны, скорее всего, потому что они ответственны за яйцу на правильном растении. [ 18 ]
В мотыльках с сумасшедшим ястребом ( Manduca Sexta ) антенны помощи при стабилизации полета. Подобно недостаткам у насекомых диптеранов, антенны передают силы кориолиса через орган Джонстона, которые затем можно использовать для корректирующего поведения. Серия исследований стабильности с низким освещением, устойчивости полета, в которых мотыльки с жгутиками ампутировались вблизи цветной части, показали значительно снижение стабильности полета по сравнению с интактными антеннами. [ 19 ] Чтобы определить, могут ли быть другие чувственные входы антенна, вторая группа мотыльков была ампутирована их антенны, а затем повторно прикреплена, прежде чем проверить в том же исследовании стабильности. Эти мотыльки показали слегка сниженную производительность от неповрежденных моли, что указывает на то, что есть, возможно, другие сенсорные входы, используемые при стабилизации полета. Повторное усиление антенн привело к резкому снижению стабильности полета, чтобы соответствовать уровню первой ампутированной группы. [ Цитация необходима ]
Ссылки
[ редактировать ]- ^ Чепмен, RF (1998). Насекомые: структура и функция (4 -е изд.). Издательство Кембриджского университета. С. 8–11 . ISBN 978-0521570480 .
- ^ Jump up to: а беременный в дюймовый Boxshall, Джефф; Jaume, D. (2013). Функциональная морфология и разнообразие: антенны и антенны в ракообразных . Издательство Оксфордского университета. С. 199–236. doi : 10.1093/acprof: osobl/9780195398038.003.0007 .
- ^ Fortey, Richard A.; Томас, Ричард Х. (1998). Отношения членистоногих: филогенетический анализ (1 -е изд.). Систематическая ассоциация. п. 117. ISBN 978-94-011-4904-4 .
- ^ Коттон, Тревор Дж.; Брэдди, Саймон Дж. (2004). «Филогения арахноморфных членистоногих и происхождение челицераты». Земля и экологическая наука Сделок Королевского общества Эдинбурга . 94 (3): 169–193. doi : 10.1017/s0263593300000596 . S2CID 54841296 .
- ^ Boxshall, GA (2004). «Эволюция конечностей членистоногих». Биологические обзоры Кембриджского философского общества . 79 (2): 253–300. doi : 10.1017/s1464793103006274 . PMID 15191225 . S2CID 25339728 .
- ^ Luque, J.; Фельдманн, RM; Вернигора, О.; Schweitzer, CE; Кэмерон, CB; Керр, Ка; Вега, FJ; Duque, A.; Странно, м.; Палмер, Ар; Jaramillo, C. (2019). «Исключительное сохранение средне-мозговых членистоногих членистоногих и эволюция новых форм через гетерохронию» . Наука достижения . 5 (4): EAAV3875. Bibcode : 2019scia .... 5.3875l . doi : 10.1126/sciadv.aav3875 . PMC 6482010 . PMID 31032408 .
- ^ Дарби, Джин (1958). Что такое бабочка? Полем Чикаго: бенефициальная пресса . п. 8. OCLC 1391997 .
- ^ Гуллан, Пенни Дж.; Cranston, Peter S. (2005). Насекомые: схема энтомологии (3 -е изд.). Оксфорд, Великобритания: Blackwell Publishing . п. 38 ISBN 978-1-4051-1113-3 .
- ^ Чепмен, Реджинальд Фредерик (1998). Насекомые: структура и функция (4 -е изд.). Нью -Йорк: издательство Кембриджского университета . п. 8 ISBN 978-0-521-57890-5 .
- ^ Jump up to: а беременный в Томас А. Кейл (1999). «Морфология и развитие периферических обонятельных органов». В Ханссон, Билл С. (ред.). Организация насекомых (1 -е изд.). Спрингер . С. 5–48. ISBN 978-3-540-65034-8 .
- ^ Насекомые: схема энтомологии, четвертое издание
- ^ Пасс, Гюнтер. Анатомия и ультраструктура антеннальных циркуляторных органов в кокчаферском жуке Melolontha Melolontha L. (Coleoptera, Scarabaeidae) Zoomorphology 01/1980; 96 (1): 77-89. Два : 10.1007/bf00310078
- ^ Лоуренс, Элеонора, изд. (2005). Словарь Хендерсона биологических терминов (13 -е изд.). Пирсон Образование . п. 51 ISBN 978-0-13-127384-9 .
- ^ «Электроантрография (EAG)» . GEORG-AUGUST-UNIVERITät Göttingen . Получено 27 марта 2010 года .
- ^ Мерлин, Кристина; Гегир, Роберт Дж.; Репперт, Стивен М. (сентябрь 2009 г.). «Антеннальные циркадные часы координируют ориентацию солнечного компаса у бабочек миграционного монарха» . Наука . 325 (5948): 1700–1704. Bibcode : 2009Sci ... 325.1700M . doi : 10.1126/science.1176221 . PMC 2754321 . PMID 19779201 .
- ^ Эллис, Пегги; Бримакомб, Линда (1980). «Сопрягающее поведение египетской хлопковой мотыльки, Spodoptera littoralis (Boisd.)». Поведение животных . 28 (4): 1239–1248. doi : 10.1016/s0003-3472 (80) 80112-6 . S2CID 53188823 .
- ^ Юстус, Ка; Митчелл, BK (ноябрь 1996 г.). «Выбор сайта яйцекладки с помощью Diamondback Moth, Plutella Xylostella (L.) (Lepidoptera: Plutellidae)». Журнал поведения насекомых . 9 (6): 887–898. Bibcode : 1996jibeh ... 9..887j . doi : 10.1007/bf02208976 . S2CID 28455636 .
- ^ Мичиган, Восточный Лансинг; RV, Dowell (1991) MI) , Scriber, JM (Университет штата . .. ISSN 0090-0222 .
{{cite journal}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ SANE, Sanjay P.; Dieudonné, Александр; Уиллис, Марк А.; Даниэль, Томас Л. (2007-02-09). «Антеннальные механососенсоры опосредуют управление полетом в мотынах». Наука . 315 (5813): 863–866. Bibcode : 2007sci ... 315..863s . Citeseerx 10.1.1.205.7318 . doi : 10.1126/science.1133598 . ISSN 0036-8075 . PMID 17290001 . S2CID 2429129 .
| Базы данных управления авторитетом : национальный |
|---|