Дигенея
| Дигенея | |
|---|---|
| |
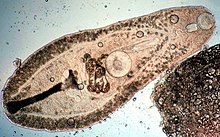
| Хеликометра сп. ( Plagiorchiida : Opecoelidae ) из кишечника Flame Cardinal рыбы . | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Платихельминты |
| Подтип: | Рабдитофора |
| Суперкласс: | неодерматы |
| Сорт: | Трематоды |
| Подкласс: | Дигенея Дорогая , 1863 г. |
| Семьи | |
Дигенея (греч. Dis — двойной, Genos — раса) — класс трематод , одной брюшной из Platyhelminthes типа , состоящий из паразитических плоских червей (известных как сосальщики ) с синцитиальным тегументом и, обычно, двумя присосками и одной ротовой. Взрослые особи обычно живут в пищеварительном тракте, но встречаются и во всех системах органов всех классов позвоночных . Когда-то считалось, что они связаны с Monogenea , теперь признано, что они наиболее близки к Aspidogastrea и что Monogenea более тесно связаны с Cestoda . На сегодняшний день описано около 6000 видов.
Морфология
[ редактировать ]Ключевые особенности
[ редактировать ]Характерные особенности Digenea включают синцитиальный тегумент; то есть тегумент, в котором соединения между клетками разрушены, и все животное окружает единая сплошная цитоплазма. Подобный тегумент встречается и у других представителей Neodermata ; группа платихельминтов, включающая Digenea, Aspidogastrea , Monogenea и Cestoda . Дигенеи обладают червеобразным , несегментированным строением тела и твердой паренхимой без полости тела ( целома ), как у всех платихельминтов.

Обычно имеется две присоски : передняя ротовая присоска, окружающая рот , и брюшная присоска, иногда называемая вертлужной впадиной , на вентральной поверхности. Ротовая присоска окружает рот, а брюшная присоска представляет собой слепой мышечный орган, не связанный с какой-либо внутренней структурой.
Моностом – червь с одной присоской (ротовой). Сосальщики с ротовой присоской и вертлужной впадиной на заднем конце тела называются амфистомами . Дистомы — это сосальщики с ротовой и брюшной присосками, но брюшная присоска находится не сзади. Эти термины часто встречаются в более старой литературе, когда считалось, что они отражают систематические отношения внутри групп. Они вышли из употребления в современной дигенейской систематике.
Репродуктивная система
[ редактировать ]Подавляющее большинство дигенеев — гермафродиты . Вероятно, это адаптация к низкой численности хозяев, позволяющая продолжить жизненный цикл, когда только одна особь успешно заражает окончательного хозяина. Оплодотворение внутреннее, при этом сперма переносится через циррус в канал Лаурера или половое отверстие . Ключевой группой раздельнополых двудомных являются шистосомы . Бесполое размножение на первой личиночной стадии встречается повсеместно.
Хотя о половом формировании дигенеевых яиц и бесполом размножении на первой личиночной стадии (мирацидии) широко сообщается, биология развития бесполых стадий остается проблемой. Электронно-микроскопические исследования показали, что светлые, видимые под микроскопом зародышевые шарики состоят из митотически делящихся клеток , которые дают начало эмбрионам и линии новых зародышевых клеток , которые включаются в эти эмбриональные стадии. Поскольку отсутствие мейотических процессов не доказано, точное определение остается сомнительным.
Мужские органы
[ редактировать ]Протандрия — общее правило среди дигенеев. Обычно два семенника присутствуют , но у некоторых сосальщиков их может быть более 100. Также присутствуют выносящие протоки , семявыносящие протоки , семенной пузырек , семявыбрасывающий проток и циррус (аналог полового члена), обычно (но не всегда) заключенные в половую бурсу. . Циррус может быть покрыт или не покрыт белковыми шипами. Точная конформация этих органов в терминальных гениталиях самцов таксономически важна на семейном и родовом уровнях.
Женские органы
[ редактировать ]Обычно имеется один яичник с яйцеводом , семяприемник , пара желточных желез (участвующих в выработке желтка и яичной скорлупы) с протоками, оотип (камера, в которой образуются яйца), сложная совокупность железистых клеток, называемая Железа Мелиса , которая, как полагают, смазывает матку для прохождения яйцеклетки.
Кроме того, у некоторых дигенеев имеется канал, называемый каналом Лаурера , который ведет от яйцевода к дорсальной поверхности тела. Функция этого канала дискуссионна, но его можно использовать для осеменения у некоторых видов или для удаления отходов размножения у других видов. У большинства трематод есть яйцевод — увеличенная часть яйцевода, где он соединяется с яичником. Вероятно, он контролирует выброс яйцеклеток и распределяет их спуск по матке.
Матка обычно открывается в общее половое предсердие, в которое также входит дистальный мужской копулятивный орган (циррус), а затем сразу же открывается на внешнюю поверхность червя. Дистальная часть матки может быть расширена в метратерм, отделенный от проксимальной части матки мышечным сфинктером, либо выстлана шипами, как у Monorchiidae и некоторых других семейств.
Пищеварительная система
[ редактировать ]Во взрослом возрасте большинство дигенеев обладают терминальным или субтерминальным ртом, мускулистой глоткой, обеспечивающей силу для заглатывания пищи, и раздвоенной слепой пищеварительной системой, состоящей из двух трубчатых мешочков, называемых слепыми отростками (sing. caecum ). У некоторых видов две слепые кишки кишки соединяются сзади, образуя кольцеобразную кишку или циклоцель . У других слепая кишка может сливаться сзади со стенкой тела, образуя один или несколько анусов , или с экскреторным пузырьком, образуя уропрокт . способны напрямую поглощать питательные вещества через тегумент путем пиноцитоза и фагоцитоза синцития Digeneans также . позвоночных Большинство взрослых дигенеев встречаются в пищеварительном канале или связанных с ним органах , где они чаще всего питаются содержимым просвета (например, пищей, проглоченной хозяином, желчью, слизью), но они также могут питаться через стенку слизистой оболочки (например, подслизистая оболочка , кровь хозяина). Кровеносные сосальщики, такие как шистосомы, спиорхииды и сангвиниколиды, питаются исключительно кровью. Бесполые стадии у моллюсков- промежуточных хозяев питаются в основном за счет прямого всасывания, хотя стадия redia , обнаруженная в некоторых группах, действительно имеет рот, глотку и простую кишку и может активно поедать ткани хозяина или даже других паразитов. Инцистированные стадии метацеркария и свободноживущие стадии церкария не питаются.
Нервная система
[ редактировать ]Парные ганглии на переднем конце тела служат мозгом . От него нервы отходят вперед и назад. Сенсорные рецепторы у взрослых по большей части отсутствуют, хотя тангорецепторные клетки у них есть. Личиночные стадии имеют множество видов сенсорных рецепторов, включая световые рецепторы и хеморецепторы. Хеморецепция играет важную роль в распознавании и определении местонахождения свободноживущей мирацидной личинки своего хозяина.
Жизненные циклы
[ редактировать ]Существует ошеломляющее разнообразие вариаций сложного жизненного цикла дигенеев, и пластичность этой черты, вероятно, является ключом к успеху группы. Как правило, в жизненном цикле могут быть два, три или четыре облигатных (необходимых) хозяина, иногда с транспортными или паратеническими хозяевами между ними. Жизненный цикл с тремя хостами, вероятно, является наиболее распространенным. Почти у всех видов первым хозяином в жизненном цикле является моллюск . [ 2 ] Это привело к выводу, что предки дигеней были моллюсками-паразитами и что впоследствии к ним были добавлены позвоночные-хозяева.
Чередование полового и бесполого поколений — важная особенность дигенеев. Это явление предполагает наличие нескольких дискретных поколений в одном жизненном цикле.
Типичный жизненный цикл дигенейских трематод выглядит следующим образом. Яйца покидают позвоночного хозяина с фекалиями и используют различные стратегии для заражения первого промежуточного хозяина , у которого половое размножение не происходит. Дигенеи могут заразить первого промежуточного хозяина (обычно улитку ) пассивным или активным путем. Яйца некоторых дигенеев, например, (пассивно) поедаются улитками (или, реже, кольчатыми червями), [ 2 ] в котором они продолжают вылупляться. Альтернативно, яйца могут вылупляться в воде, чтобы выпустить активно плавающую реснитчатую личинку, мирацидий , которая должна обнаружить и проникнуть в стенку тела улитки-хозяина.
После вылупления после проглатывания или проникновения в улитку мирацидий превращается в простую мешковидную материнскую спороцисту . Материнская спороциста подвергается циклу внутреннего бесполого размножения , давая начало либо редиям (sing. redia), либо дочерним спороцистам . Таким образом, второе поколение представляет собой дочернюю последовательность парфениты. Они, в свою очередь, подвергаются дальнейшему бесполому размножению, в конечном итоге давая большое количество второй свободноживущей стадии, церкарий (мн. cercariae).
Свободно плавающие церкарии покидают хозяина-улитку и перемещаются по водной или морской среде, часто используя хлыстообразный хвост, хотя наблюдается огромное разнообразие морфологии хвоста. Церкарии заразны для второго хозяина в жизненном цикле, и заражение может происходить пассивно (например, рыба потребляет церкарии) или активно (церкарии проникают в рыбу).
Жизненный цикл некоторых дигенеев включает только двух хозяев, второй из которых является позвоночным. В этих группах половая зрелость наступает после проникновения церкарий во второго хозяина, который в данном случае является и окончательным хозяином . Жизненные циклы с двумя хозяевами могут быть первичными (третьего хозяина никогда не было), как у Bivesiculidae , или вторичными (когда-то в эволюционной истории существовал третий хозяин, но он был утерян).
В жизненных циклах с тремя хозяевами церкарии развиваются во втором промежуточном хозяине в стадию покоя, метацеркарию , которая обычно инцистируется в кисте хозяина и паразитического происхождения или инкапсулируется в слой ткани, полученной только от хозяина. Эта стадия заразна для окончательного хозяина . Передача происходит, когда окончательный хозяин охотится на зараженного второго промежуточного хозяина. Метацеркарии эксцистируются в кишечнике окончательного хозяина в ответ на различные физические и химические сигналы, такие как уровень pH кишечника , пищеварительные ферменты , температура и т. д. После эксцистирования взрослые дигенеи мигрируют в более или менее специфические участки окончательного хозяина и живого организма. цикл повторяется.
Эволюция
[ редактировать ]Эволюционное происхождение дигенеи обсуждалось в течение некоторого времени, но, по-видимому, существует общее мнение, что протодигенея была паразитом моллюска, возможно, обитавшего в мантийной полости. Доказательством этого является повсеместное распространение моллюсков в качестве первых промежуточных хозяев для дигенеев, а также тот факт, что большинство аспидогастреев (сестринская группа дигенеи) также имеют ассоциации с моллюсками. Считается, что ранние трематоды (собирательное название дигенеев и аспидогастрий), вероятно, произошли от рабдоцелевых турбеллярий, колонизировавших открытую мантийную полость ранних моллюсков.
Вполне вероятно, что более сложные жизненные циклы развились в процессе терминального добавления, в результате чего дигенеи пережили хищничество своего моллюска-хозяина, вероятно, рыбы. Другие хосты добавлялись тем же процессом, пока не развилось современное ошеломляющее разнообразие моделей жизненного цикла.
Важные семьи
[ редактировать ]Дигенея включает около 80 семейств. [ 3 ] Они перечислены ниже, упорядочены по порядку.
Дигенея
- Диплостомида
- Подотряд Diplostomata
- Надсемейство Brachylaimoidea Joyeux & Foley, 1930 г.
- Brachylaimidae Joyeux & Foley, 1930 г.
- Leucoхлоридiidae Poche, 1907 г.
- Надсемейство Diplostomoidea Poirier, 1886 г.
- Брауниниды Волк, 1903 г.
- Cyathocotylidae Mühling, 1898 г.
- Диплостомиды Пуарье, 1886 г.
- Proterodiplostomidae Dubois, 1936.
- Strigeidae Railliet, 1919 г.
- Надсемейство Schistosomatoidea Stiles & Hassall, 1898 г.
- Апорокотилиды Однер, 1912 г.
- Schistosomatidae Стайлз и Хассал, 1898 г.
- Spirorchiidae Stunkard, 1921 г.
- Надсемейство Brachylaimoidea Joyeux & Foley, 1930 г.
- Подотряд Diplostomata
- Плагиат
- Апокредиата
- Апокредиоидея Скрябин, 1942 г.
- Апокреадииды Скрябин, 1942
- Апокредиоидея Скрябин, 1942 г.
- Бивезикулата
- Бивезикулоидея
- Бивесикулиды Ямагути, 1934 г.
- Бивезикулоидея
- Буцефалата
- Буцефалоидея Поче, 1907 г.
- Буцефалиды Поче, 1907 г.
- Nuitrematidae Курочкин, 1975 г.
- Гимнофаллоидея Однера, 1905 г.
- Botulisaccidae Ямагути, 1971 г.
- Fellodistomidae Николл, 1909 г.
- Гимнофаллиды Однер, 1905 г.
- Танданиколиды Джонстон, 1927 г.
- Буцефалоидея Поче, 1907 г.
- Ихиностомы
- Эхиностомоидеа Лусс, 1902 г.
- Каликодиды Дольфус , 1929 г.
- Cyclocoelidae Stossich, 1902 г.
- Echinochasmidae Odhner, 1910 г.
- Эхиностоматиды Лусс, 1899 г.
- Эвкотилиды Кона, 1904 г.
- Fasciolidae Railliet, 1895 г.
- Химастлиды Однер, 1910 г.
- Филофтальмиды Лусс, 1899 г.
- Псилостомиды Лусс, 1900 г.
- Ритидодиды Однер, 1926 г.
- Typhlocoelidae Harrah, 1922 г.
- Эхиностомоидеа Лусс, 1902 г.
- Хаплопланхната
- Haploplanchnoidea Poche, 1925 г.
- Haploplanchnidae Poche, 1926 г.
- Haploplanchnoidea Poche, 1925 г.
- Гемиурат
- Азигиоидея Люэ, 1909 г.
- Азигииды Люэ, 1909 г.
- Гемиуродея Лусс, 1899 г.
- Accacoeliidae Odhner, 1911 г.
- Bathycotylidae Dollfus , 1932 г.
- Дерогениды Николл, 1910 г.
- Dictysarcidae Скрябин и Гущанская, 1955
- Didymozoidae Monticelli, 1888 г.
- Гоноцерциды Скрябина и Гущанская, 1955.
- Hemiuridae Looss, 1899 г.
- Hirudinellidae Dollfus , 1932 г.
- Isoparorchiidae Travassos, 1922 г.
- Лецитастериды Однер, 1905 г.
- Ptychogonimidae Dollfus , 1937 г.
- Sclerodistomidae Однер, 1927 г.
- Sclerodistomoididae Gibson & Bray, 1979.
- Syncoeliidae Looss, 1899 г.
- Азигиоидея Люэ, 1909 г.
- Герониматы
- Херонимоидея Уорд, 1918 год.
- Геронимиды Уорд, 1918 год.
- Херонимоидея Уорд, 1918 год.
- Красиво создано
- Лепокредиоидеа Однер, 1905 г.
- Aephnidiogenidae Ямагути, 1934 г.
- Деропристиды Кейбл и Хуннинен, 1942 г.
- Энэнтериды Ямагути, 1958 г.
- Горгоцефалиды Мантер, 1966 г.
- Gyliauchenidae Фукуи, 1929 г.
- Лепидапедиды Ямагути, 1958 г.
- Лепокреадииды Однер, 1905 г.
- Liliatrematidae Губанов, 1953
- Лепокредиоидеа Однер, 1905 г.
- Целеустремленный
- Монорхиоидеа Однер, 1911 г.
- Лиссорхиды Магат, 1917 г.
- Монорхиииды Однер, 1911 г.
- Монорхиоидеа Однер, 1911 г.
- Описторхиата
- Описторхиоидеа Браун, 1901 г.
- Криптогонимиды Уорд, 1917 г.
- Heterophyidae Лейпер, 1909 г.
- Opisthorchiidae Looss, 1899 г.
- Описторхиоидеа Браун, 1901 г.
- Проноцефалата
- Парамфистомоидея Фишодер, 1901 г.
- Cladorchiidae Fischoeder, 1901 г.
- Мезометриды Poche, 1926 г.
- Microscaphidiidae Лусс, 1900 г.
- Paramphistomidae Fischoeder, 1901 г.
- Проноцефалоидея Лусс, 1899 г.
- Лабиколиды Блэр, 1979 г.
- Notocotylidae Lühe, 1909 г.
- Нудакотилиды Баркер, 1916 г.
- Opisthotrematidae Poche, 1926 г.
- Проноцефалиды Лусс, 1899 г.
- Рабдиопеиды Поче, 1926 г.
- Парамфистомоидея Фишодер, 1901 г.
- Трансверсотрематы
- Transversotrematoidea Витенберг, 1944 г.
- Transversotrematidae Витенберг, 1944 г.
- Transversotrematoidea Витенберг, 1944 г.
- Ксифидиата
- Аллокредиоидеа Лусс, 1902 г.
- Acanthocolpidae Люэ, 1906 г.
- Аллокреадииды Лусс, 1902 г.
- Батрахотрематиды Дольфус и Уильямс, 1966 г.
- Брахикладииды Однер, 1905 г.
- Опекоэлиды Озаки, 1925 г.
- Горгодероидеа Лусс, 1901 г.
- Callodistomidae Однер, 1910 г.
- Dicrocoeliidae Looss, 1899 г.
- Горгодериды Лусс, 1899 г.
- Гаплопороидеа Николл, 1914 г.
- Atractotrematidae Ямагути, 1939 г.
- Гаплопориды Николл, 1914 г.
- Микрофаллоидея Уорд, 1901 год.
- Диплангиды Ямагути, 1971 г.
- Exotidendriidae Mehra, 1935 г.
- Фаустулиды Поче, 1926 г.
- Микрофаллиды Уорд, 1901 г.
- Pachypsolidae Ямагути, 1958 г.
- Фанеропсолиды Мехра, 1935 г.
- Pleurogenidae Looss, 1899 г.
- Простогонимиды Люэ, 1909 г.
- Рениколиды Дольфус , 1939 г.
- Зоогониды Однер, 1902 г.
- Плагиорхиоидеа Люэ, 1901 г.
- Ауридистомиды Люэ, 1901 г.
- Брахикоелииды Лусс, 1899 г.
- Цефалогонимиды Лусс, 1899 г.
- Choanocotylidae Джу Сью и Платт, 1998 г.
- Echinoporidae Краснолобова и Тимофеева, 1965.
- Энциклометриды Мехра, 1931 г.
- Лептофаллиды Даял, 1938 г.
- Macroderoididae МакМаллен, 1937 г.
- Meristocotylidae Fischthal & Kuntz, 1981.
- Ocadiatrematidae Fischthal & Kuntz, 1981.
- Orientocreadiidae Ямагути, 1958 г.
- Plagiorchiidae Люэ, 1901 г.
- Стифлотрематиды Баер, 1924 г.
- Telorchiidae Looss, 1899 г.
- Thrinascotrematidae Джу Сью и Платт, 1999 г.
- Urotrematidae Poche, 1926 г.
- Аллокредиоидеа Лусс, 1902 г.
- Апокредиата
Дигенеевые инфекции человека
[ редактировать ]Только около 12 из 6000 известных видов заразны для человека, но некоторые из этих видов представляют собой серьезные заболевания, от которых страдают более 200 миллионов человек. Виды, поражающие человека, можно разделить на группы: шистосомы и нешистосомы.
Шистосомы
[ редактировать ]Шистосомы встречаются в кровеносной системе окончательного хозяина. Человек заражается после проникновения в кожу свободно плавающих церкарий, выделенных из зараженных улиток. Эти раздельнополые черви длинные и тонкие, размером от 10–30 мм в длину до 0,2–1,0 мм в диаметре. Взрослые самцы короче и толще самок и имеют длинную бороздку вдоль одной стороны тела, в которой зажата самка. Самки достигают половой зрелости после соединения с самцом. После спаривания они остаются запертыми вместе до конца своей жизни. Они могут жить несколько лет и производить многие тысячи яиц.
Четыре вида шистосом, поражающих человека, относятся к роду Schistosoma .
| Научное название | Первый промежуточный хост | Эндемичная зона |
|---|---|---|
| Шистосома Мансони | Биомфалярии виды. | Африка , Южная Америка , Карибский бассейн , Средний Восток |
| Шистосома гематобиум | Булинус виды. | Африка , Ближний Восток |
| Шистосома японская | Онкомелания виды. | Китай , Восточная Азия , Филиппины |
| Вставочная шистосома | Булинус виды. | Африка |
Нешистосомы
[ редактировать ]Ниже перечислены семь основных видов нешистосом, поражающих человека. Люди заражаются после проглатывания метацеркариальных цист на растениях или недоваренного мяса животных. Большинство видов обитают в желудочно-кишечном тракте человека, где откладывают яйца вместе с фекалиями хозяина. Paragonimus westermani , колонизирующий легкие, также может выделять яйца через слюну . Эти трематоды обычно вызывают у людей легкую патологию, но могут возникнуть и более серьезные последствия.
| Научное название | Первый промежуточный хост | Способ заражения человека | Эндемичная зона |
|---|---|---|---|
| Фасциолопсис буски | Сегментина сп. | Растения | Азия , Индия |
| Гетерофии | Пиринелла [ нужна ссылка ] | Кефаль , Тилапия | Азия , Восточная Европа , Египет , Ближний Восток |
| Метагонимус йокогавайи | Полусулькоспира сп. [ нужна ссылка ] | Карп , Форель | Сибирь |
| Гастродискоиды человека | Хеликорбис сп. [ нужна ссылка ] | Растения | Индия , Вьетнам , Филиппины |
| Клонорхис китайский | Булинус сп. | Рыба | Восточная Азия , Северная Америка |
| Фасциола печеночная | Гальба усеченная | Растения | По всему миру |
| Парагонимус вестермани | Онкомелания сп. | Крабы , раки | Азия |
Ссылки
[ редактировать ]- ^ Брей, РА; Жюстин, Дж.Л. (2014). «Обзор Zoogonidae (Digenea: Microphalloidea) из рыб вод Новой Каледонии с описанием Overstreetia cribbi n. sp» . ПерДж . 2 : е292. дои : 10.7717/peerj.292 . ПМЦ 3961169 . ПМИД 24688868 .
- ^ Jump up to: а б «Принципы паразитизма: Дигенея» . www.biology.ualberta.ca . Проверено 9 января 2020 г.
- ^ Олсон П.Д., Крибб Т.Х., Ткач В.В., Брей Р.А. и Литтлвуд DTJ (2003). «Филогения и классификация Digenea (Platyhelminthes: Trematoda)». Международный журнал паразитологии 33 (7): 733–755. два : 10.1016/S0020-7519(03)00049-3
Примечания
[ редактировать ]- Гибсон Д.И., Джонс А. и Брей Р.А. (2002). Ключ к Трематоде, том 1 ISBN 0-85199-547-0
- Литтлвуд DTJ и Брей Р.А. (2001) Взаимоотношения Platyhelminthes. ISBN 0-7484-0903-3
- Ямагути, С. (1971). Конспект дигенетических трематод позвоночных. Издательство Кэйгаку, Токио.
Внешние ссылки
[ редактировать ]- Группа исследования шистосом Кембриджского университета
- Паразитические черви в Музее естественной истории в Лондоне
- Fishdisease.net
| Базы данных органов управления : Национальные |
|---|