Планы тела докембрия
Эта статья включает список общих ссылок , но в ней отсутствуют достаточные соответствующие встроенные цитаты . ( Май 2015 г. ) |
До конца 1950-х годов не считалось, что в докембрии существовали многоклеточные организмы. Однако с помощью методов радиометрического датирования было обнаружено, что окаменелости , первоначально найденные на холмах Эдиакара в Южной Австралии, относятся к позднему докембрию. Эти окаменелости представляют собой отпечатки тел организмов, имеющих форму дисков, листьев , а некоторые имеют ленточные узоры, которые, скорее всего, были щупальцами .
Это самые ранние многоклеточные организмы в истории Земли, несмотря на то, что одноклеточные существовали задолго до этого. Требования многоклеточности были заложены в генах некоторых из этих клеток, особенно хоанофлагеллят . Считается, что они являются предшественниками всех животных. Они тесно связаны с губками (Porifera), простейшими многоклеточными животными.
Чтобы понять переход к многоклеточности в докембрии, важно взглянуть на требования многоклеточности — как биологические, так и экологические.
Докембрий
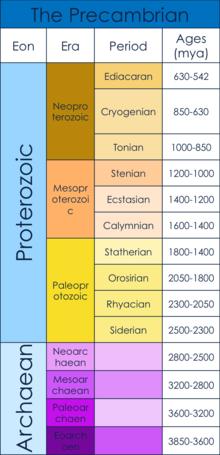
[ редактировать ]Докембрий датируется началом формирования Земли (4,6 миллиарда лет назад) до начала кембрийского периода , 539 миллионов лет назад. [ 1 ] [ 2 ] Докембрий состоит из гадейского , архейского и протерозойского эонов. [ 1 ] В частности, в этой статье рассматривается Эдиакарский период , когда, как полагают, возникли первые многоклеточные тела, а также то, что вызвало возникновение многоклеточности. [ 3 ] Этот временной период возник после Земли-снежка среднего неопротерозоя. «Земля-снежок» была периодом мирового оледенения , которое, как полагают, послужило узким местом популяций для последующей эволюции многоклеточных организмов. [ 4 ]
Докембрийские тела
[ редактировать ]Земля образовалась около 4,6 миллиарда лет назад, а одноклеточная жизнь появилась несколько позже, после прекращения поздней тяжелой бомбардировки, периода интенсивных ударов астероидов, возможно, вызванных миграцией планет-газовых гигантов на их нынешние орбиты, однако многоклеточность и тела являются относительно недавнее событие в истории Земли. [ 5 ] Тела впервые начали появляться ближе к концу докембрии , в эдиакарский период. Окаменелости эдиакарского периода были впервые найдены в Южной Австралии на холмах Эдиакара , отсюда и название. Однако первоначально считалось, что эти окаменелости относятся к кембрийскому периоду , и только в конце 1950-х годов Мартин Глесснер определил, что эти окаменелости на самом деле относятся к докембрийской эпохе. Найденные окаменелости датируются примерно 600 миллионами лет назад и имеют различную морфологию. [ 5 ]
Окаменелости Эдиакарского моря
[ редактировать ]Дополнительную информацию см. в разделе «Биота Эдиакара» .
Обнаруженные окаменелости, относящиеся к докембрию, не имеют четкой структуры, поскольку в этот период не существовало скелетных форм. [ 5 ] Скелеты не возникали до кембрийского периода, когда уровень кислорода увеличился. Это связано с тем, что скелетам необходим коллаген , который использует витамин С в качестве кофактора, которому необходим кислород . [ 6 ] Дополнительную информацию о повышении уровня кислорода см. в разделе, посвященном кислороду. Большинство окаменелостей этой эпохи происходит либо из Мистейкен-Пойнт на восточном побережье Канады, либо из холмов Эдиакара в Южной Австралии . [ 5 ]
Большинство окаменелостей находят как отпечатки мягкотелых организмов в форме дисков, лент или листьев. [ 3 ] [ 5 ] Есть также следы окаменелостей, которые свидетельствуют о том, что некоторые из этих докембрийских организмов, скорее всего, были червеобразными существами, которые были локомотивами. [ 7 ] У большинства этих окаменелостей отсутствуют узнаваемые головы, рты или органы пищеварения, и считается, что они питались посредством абсорбционных механизмов и симбиотических отношений с хемоавтотрофами ( хемотрофами ), фотоавтотрофами ( фототрофами ) или осмоавтотрофами. [ 1 ] Лентообразные окаменелости напоминают организмы с щупальцами и, как полагают, питались, захватывая добычу. Окаменелости фрондозы напоминают морские загоны и других книдарий . Следы окаменелостей позволяют предположить, что существовали существа типа кольчатых червей , а окаменелости дисков напоминают губки. Несмотря на это сходство, большая часть идентификации является предположением, поскольку окаменелости не имеют четко выраженной структуры. Другие окаменелости не похожи ни на одну из известных линий. [ 1 ]
Многие из организмов, такие как Charnia , найденные в Mistaken Point , не были похожи ни на один организм, наблюдаемый сегодня. У них были отдельные тела, однако у них не было головы и пищеварительных отделов. Скорее, их тело было организовано по очень простой, напоминающей фрактальную структуру ветвления. [ 8 ] Каждый элемент тела был тонко разветвлен и рос за счет повторяющихся ветвей. Это позволило организму иметь большую площадь поверхности и максимизировать усвоение питательных веществ без необходимости использования рта и пищеварительной системы . Однако генетическая информация была минимальной, и поэтому не было требований, которые позволили бы им разработать более эффективные методы кормления. Это означает, что они, вероятно, были вытеснены другими организмами и, таким образом, вымерли. [ 8 ]
Организмы, обнаруженные на Эдиакарских холмах в Южной Австралии, демонстрировали либо радиально-симметричное строение тела, либо один организм, Сприггина, продемонстрировал первую двустороннюю симметрию. Считается, что на Эдиакарских холмах когда-то был неглубокий риф , через который больше света могло проникать на дно океана. Это позволило добиться большего разнообразия организмов. Найденные здесь организмы напоминают родственников книдарий , моллюсков или кольчатых червей . [ 8 ]
Чарния
[ редактировать ]
Окаменелости чарнии были первоначально найдены в Чарнвудском лесу в Англии , поэтому и названы Чарния . [ 8 ] Эти окаменелости принадлежат морским организмам, обитавшим на дне океана. Окаменелости имеют фрактальную форму тела и форму листьев, что означает, что они напоминали широколиственные растения, такие как папоротники. Однако они не могли быть растениями, поскольку обитали в темных глубинах океанского дна. В Чарнвудском лесу чарния была обнаружена как изолированный вид, однако на восточном побережье Канады в Мистейкен-Пойнт в Ньюфаундленде было обнаружено гораздо больше окаменелостей. Чарния была прикреплена ко дну океана и находилась под сильным течением. есть дискообразные формы Это заметно потому, что в нижней части окаменелости Чарнии , которые показывают, где была привязана Чарния , и все близлежащие окаменелости обращены в одном направлении. Эти окаменелости в Мистейкен-Пойнт хорошо сохранились под вулканическим пеплом и слоями мягкой грязи. [ 8 ] С помощью радиометрического датирования окаменелостей было установлено, что Чарния , должно быть, жила около 565 миллионов лет назад. [ 4 ] [ 9 ]
Дикинсония
[ редактировать ]
Окаменелости дикинсонии — еще одна примечательная окаменелость эдиакарского периода, найденная в Южной Австралии и России . [ 10 ] Остается неизвестным, каким типом организма была Дикинсония ; однако его считали полихетой , турбеллярием/кольчатым червем, медузой , полипом, протистом, лишайником или грибом . [ 10 ] Они сохранились в кварцевых песчаниках и датируются примерно 550 миллионами лет назад. Дикинсонии были организмами с мягким телом, демонстрирующими некоторые признаки очень медленного движения. [ 4 ] На камне есть слабые круглые отпечатки, которые следуют по тропе, а затем, по тому же пути, появляется более четкий круглый отпечаток того же размера. Это указывает на то, что организм, вероятно, медленно перемещался из одного места кормления в другое и поглощал питательные вещества. Предполагается, что у этого организма, вероятно, были очень маленькие придатки, которые позволяли ему двигаться так же, как это делают современные морские звезды. [ 11 ]
Сприггин
[ редактировать ]
Окаменелости Сприггины представляют собой первые известные организмы с двусторонне-симметричным строением тела. У них были голова, хвост и почти одинаковые половинки. [ 3 ] Вероятно, у них были органы чувств в голове и органы пищеварения в хвосте, что позволяло им более эффективно находить пищу. Они были способны к передвижению, что давало им преимущество перед другими организмами той эпохи, которые либо были привязаны ко дну океана, либо двигались очень медленно. Сприггина имела мягкое тело, которое оставляет на окаменелостях слабые отпечатки. Скорее всего, он связан с кольчатыми червями, однако есть некоторые предположения, что он может быть связан с членистоногими, поскольку он чем-то напоминает окаменелости трилобитов. [ 3 ] [ 5 ]
Следы окаменелостей
[ редактировать ]Эдиакарские . окаменелости Южной Австралии содержат следы окаменелостей, которые указывают на существование подвижных донных организмов Все организмы, производившие следы в отложениях, были червеобразными питателями отложений или детритофагами ( Detritivore ). Есть несколько окаменелостей, напоминающих следы членистоногих . Имеющиеся данные свидетельствуют о том, что организмы, подобные членистоногим, существовали в докембрии . Это свидетельство в виде следов, оставленных после себя; в частности, один экземпляр, на котором видны шесть пар симметрично расположенных отпечатков, напоминающих пешеходные тропы трилобитов. [ 7 ]
Переход от одноклеточности к многоклеточности
[ редактировать ]На протяжении большей части истории Земли жизнь была одноклеточной . Однако в одноклеточных организмах были ингредиенты, необходимые для возникновения многоклеточности. Несмотря на наличие ингредиентов для многоклеточности, существование организмов было ограничено из-за отсутствия благоприятных условий окружающей среды. Появление кислорода ( Великое событие оксигенации ) привело к тому, что организмы смогли разработать более сложные структуры тела. Чтобы возникла многоклеточность, организмы должны были быть способны к клеточной коммуникации , агрегации и специализированным функциям. Переход к многоклеточности, положивший начало эволюции животных от простейших, является одним из наиболее малоизученных событий жизни в истории. Понимание хоанофлагеллят и их связи с губками важно при выдвижении теорий о происхождении многоклеточности. [ 12 ]
Хоанофлагелляты
[ редактировать ]Хоанофлагелляты , также называемые «воротничковыми жгутиками», — это одноклеточные протисты, обитающие как в пресных водах, так и в океанах. [ 13 ] Хоанофлагелляты имеют сферическое (или яйцевидное) тело клетки и жгутик, окруженный воротником, состоящим из актиновых микроворсинок. [ 13 ] [ 14 ] Жгутик используется для облегчения движения и приема пищи. Когда жгутик бьется, он впитывает воду через микроворсинки, прикрепленные к воротнику, что помогает отфильтровывать нежелательные бактерии и другие мельчайшие частицы пищи . [ 13 ] Хоанофлагелляты насчитывают около 150 видов и размножаются простым делением. [ 15 ]
Хоанофлагеллята Salpingoeca Rosetta
[ редактировать ](также известный как хоанофлагеллята протероспонгия)
Хоанофлагеллята Salpingoeca Rosetta — редкий пресноводный эукариот , состоящий из множества клеток, заключенных в желеобразный матрикс. Этот организм демонстрирует очень примитивный уровень дифференциации и специализации клеток . [ 15 ] Это видно по жгутиковым клеткам и их воротниковым структурам, которые перемещают колонию клеток по воде, в то время как амебоидные клетки внутри служат для деления на новые клетки, способствующие росту колонии.
Подобный низкий уровень клеточной дифференциации и спецификации можно наблюдать и у губок. У них также есть воротниковые клетки (также называемые хоаноцитами из-за их сходства с хоанофлагеллятами) и амебоидные клетки, расположенные в студенистом матриксе. В отличие от хоанофлагелляты Salpingoeca Rosetta , у губок есть и другие типы клеток, которые могут выполнять разные функции (см. Губки ). Кроме того, воротниковые клетки губок бьются внутри каналов в теле губки, тогда как воротниковые клетки Salpingoeca Rosetta расположены внутри и не имеют внутренних каналов. Несмотря на эти незначительные различия, есть убедительные доказательства того, что Proterospongia и Metazoa тесно связаны. [ 15 ]
Перплекса Хоанофлагеллята
[ редактировать ]Эти хоанофлагелляты способны прикрепляться друг к другу посредством спаривания микроворсинок воротника. [ 16 ]
Хоанофлагеллята Codosiga Botrytis и Desmerella
[ редактировать ]Эти хоанофлагелляты способны образовывать колонии посредством тонких межклеточных мостиков, которые позволяют отдельным клеткам прикрепляться. Эти мостики напоминают кольцевые каналы, соединяющие развивающиеся сперматогонии или оогонии у животных. [ 16 ]
Губки (Porifera)
[ редактировать ]Губки — одни из старейших и самых распространенных животных на Земле. Появление окаменелостей спикул губки относится к докембрийской эпохе около 580 миллионов лет назад. [ 17 ] Совокупность этих окаменелостей была найдена в формации Доушанто в Южном Китае. Сообщается, что некоторые круглые отпечатки с Эдиакарских холмов в Южной Австралии также представляют собой губки. Это одна из немногих линий многоклеточных животных этой эпохи, которые продолжают выживать и остаются относительно неизменными. [ 17 ] [ 18 ] Губки являются такими успешными организмами благодаря своей простой, но эффективной морфологии . У них нет рта или пищеварительной, нервной или кровеносной систем. Вместо этого они являются фильтраторами , а это означает, что они получают пищу через питательные вещества в воде. [ 19 ] У них есть поры, называемые устьями , через которые вода проходит в камеру, называемую спонгоцель , и выходит через камеру, называемую оскулум . [ 19 ] Через эту систему фильтрации воды они получают питательные вещества, необходимые для их выживания. В частности, они внутриклеточно переваривают бактерии, микроводоросли или коллоиды. [ 20 ]
Скелеты губок состоят либо из спонгина , либо из известково-кремнистых спикул с вкраплениями некоторых молекул коллагена. [ 21 ] Коллаген скрепляет клетки губки. Различные линии губок различают по составу их скелетов. Три основных класса губок — Demospongiae , Hexactinellid и Calcareous .
Демонические губки — самый известный тип губок, поскольку они используются людьми. Они отличаются кремнистым скелетом из двух- и четырехлучевых спикул и содержат белок спонгин.
Гексатинеллиды также называют стеклянными губками и отличаются шестилучевым стеклянным скелетом. Эти губки также способны проводить потенциалы действия.
Известковые губки характеризуются карбонатным кальциевым скелетом и составляют менее 5% губок. [ 21 ]
Клетки
[ редактировать ]Губки имеют около 6 различных типов клеток, которые могут выполнять разные функции. [ 21 ] Губки являются хорошей моделью для изучения происхождения многоклеточности, поскольку клетки способны общаться друг с другом и повторно агрегировать. В эксперименте, проведенном Генри Ван Питерсом Уилсоном в 1910 году, было обнаружено, что клетки диссоциированных губок могут посылать сигналы и распознавать друг друга, чтобы сформировать новую особь. [ 22 ] Это говорит о том, что клетки, из которых состоят губки, способны к независимой жизни, однако, как только стало возможным многоклеточное существование, агрегирование вместе для формирования одного организма стало более эффективным способом жизни.
Наиболее заметными типами клеток губок являются бокаловидные клетки, называемые хоаноцитами , названные так из-за их сходства с хоанофлагеллятами. [ 21 ] Сходство между этими двумя типами клеток заставляет ученых полагать, что хоанофлагелляты являются сестринскими таксонами многоклеточных животных. Жгутики этих клеток обеспечивают движение воды через тело губки. [ 23 ] Клеточное тело хоаноцитов отвечает за поглощение питательных веществ. У некоторых видов эти клетки могут развиваться в гаметы . [ 21 ]
Пинакоциты — это клетки на внешней стороне губки, выстилающие тело клетки. Они плотно прилегают друг к другу и очень тонкие. [ 21 ]
Мезенхима . выстилает область между пинакоцитами и хоаноцитами Они содержат матрикс, состоящий из белков и спикул. [ 21 ]
Археоциты — это особые типы клеток, которые могут трансформироваться во все другие типы клеток. Они будут делать то, что необходимо в теле губки, например, заглатывать и переваривать пищу, транспортировать питательные вещества к другим клеткам тела губки. Эти клетки также способны развиваться в гаметы у некоторых видов губок. [ 21 ]
Склероциты . отвечают за секрецию спикул У видов губок, которые используют спонгин вместо известковых и кремнеземных спикул, склероциты заменены спонгоцитами, которые секретируют скелетные волокна спонгина. [ 21 ]
и . пороциты миоциты За сокращение губки отвечают Эти сокращения аналогичны сокращениям мышц у других организмов, поскольку у губок нет мышц. Они отвечают за регулирование потока воды через губку. [ 21 ]
Формирование многоклеточности
[ редактировать ]Формирование многоклеточности стало поворотным моментом в эволюции жизни на Земле. эры произошло огромное увеличение разнообразия живых организмов Вскоре после возникновения многоклеточности в начале кембрийской , получившее название «Кембрийский взрыв» . Считается, что многоклеточность неоднократно развивалась на Земле, поскольку она была полезной стратегией жизни для организмов. [ 24 ] Для возникновения многоклеточности клетки должны быть способны к самовоспроизведению, межклеточной адгезии и межклеточной коммуникации. В окружающей среде также должны были присутствовать кислород и селективное давление.
Теория клеточного деления: С. Розетта.
[ редактировать ]Работа Фэйрклафа, Дейеля и Кинга предполагает, что S. Rosetta может существовать либо в одноклеточной форме, либо в колониях из 4–50 клеток, которые образуют плотно сплоченные пакеты сфер. [ 16 ] Это было установлено путем проведения эксперимента по введению хищной бактерии вида Algoriphagus в образец одноклеточного организма S. Rosetta и контролю активности в течение 12 часов. Результаты этого исследования показали, что клеточные колонии образовывались за счет клеточного деления исходной одиночной клетки S. Rosetta, а не за счет агрегации клеток. Дальнейшие исследования в поддержку теории клеточной пролиферации были проведены путем введения и удаления препарата афидиколина, который блокирует деление клеток. При введении препарата деление клеток прекращалось и образование колоний происходило за счет межклеточной агрегации. Когда препарат был отменен, деление клеток снова стало преобладать. [ 16 ]
Строительные блоки для клеточной адгезии
[ редактировать ]Изучая геном хоанофлагелляты « Monosiga brevicollis», ученые пришли к выводу , что хоанофлагелляты играют ключевую роль в развитии многоклеточности. [ 13 ] Николь Кинг провела работу по изучению генома Monisiga brevicollis и обнаружила ключевые белковые домены, которые являются общими для многоклеточных животных и хоанофлагеллят. Эти домены играют роль в клеточной передаче сигналов и процессах адгезии у многоклеточных животных. Обнаружение того, что хоанофлагелляты также имеют эти гены, является невероятным открытием, поскольку ранее считалось, что только у многоклеточных животных есть гены, ответственные за межклеточную связь и агрегацию. Это позволяет предположить, что эти домены играют ключевую роль в происхождении многоклеточности, поскольку они связывают одноклеточный организм (хоанофлагелляты) с многоклеточными организмами (многоклеточными организмами). Это показывает, что компоненты, необходимые для многоклеточности, присутствовали у общего предка многоклеточных животных и хоанофлагеллят. [ 13 ]
Сотовая сигнализация и сотовая связь
[ редактировать ]Ни губки, ни плакозойные особи Trichoplax adhaerens, по-видимому, не оснащены нейронными синапсами , однако оба они обладают несколькими факторами, связанными с одной и той же синаптической функцией. [ 25 ] Следовательно, вполне вероятно, что центральные особенности, участвующие в синаптической передаче, возникли на ранних стадиях эволюции многоклеточных животных, скорее всего, примерно в то время, когда большая часть жизни на Земле переходила к многоклеточности. Было обнаружено, что комплекс Munc18 / синтаксин 1 может быть важным компонентом продукции белка SNARE . Считается, что секреция белка SNARE из синаптических везикул имеет решающее значение для нейронной коммуникации . Комплекс Munc18/синтаксин 1, обнаруженный у M. brevicollis, структурно и функционально подобен комплексу многоклеточных животных. Это предполагает, что это представляет собой важный шаг на пути реакции сборки SNARE . Считается, что общий предок хоанофлагеллят и многоклеточных животных использовал этот первичный механизм секреции в качестве предшественника синаптической коммуникации. Этот механизм в конечном итоге будет использоваться для межклеточной коммуникации у животных. [ 25 ]
Причины развития многоклеточности
[ редактировать ]Несмотря на то, что прокариотические клетки содержали строительные блоки, необходимые для возникновения многоклеточности, этот переход не произошел примерно через 1500 миллионов лет после возникновения первой эукариотической клетки. [ 12 ] Ученые предложили две основные теории по причине того, что многоклеточность возникла так поздно после появления жизни на Земле.
Теория хищничества многоклеточности
[ редактировать ]Эта теория постулирует, что многоклеточность возникла как средство спасения добычи от хищников. На более крупную добычу с меньшей вероятностью будут охотиться, а более крупные хищники с большей вероятностью поймают добычу. Поэтому вполне вероятно, что многоклеточность возникла, когда появились первые хищники. Объединившись в более крупный многоклеточный организм, добыча могла избежать посягательств хищника. [ 12 ] Поэтому многоклеточность избирательно отдавалась предпочтение одноклеточности. Это можно увидеть в простом эксперименте, проведенном Бораасом и др. (1998). [ 26 ] Когда хищный протист Ochromonas valencia был введен в популяцию Chlorella vulgaris , являющуюся добычей, было замечено, что менее чем за 100 поколений вида-жертвы многоклеточная форма роста водоросли стала доминирующей. поскольку до появления хищника популяция Chlorella Это интересно , vulgaris сохраняла одноклеточную форму роста на протяжении тысяч поколений. Вероятно, он оставался бы одноклеточным на неопределенный срок, если бы не было введено селективное давление, вызванное хищниками. После нескольких поколений с хищником виды водорослей сохранили форму роста из 8-10 клеток, которые были достаточно большими, чтобы избежать хищника, но достаточно маленькими, чтобы каждая клетка все еще имела доступ к питательным веществам. [ 26 ] Эти отношения хищник-жертва, вероятно, являются причиной того, почему организмам было выгодно быть многоклеточными.
Теория повышения уровня кислорода при многоклеточности
[ редактировать ]Несмотря на то, что организмы могли стать многоклеточными, вполне вероятно, что это было невозможно до позднего неопротерозоя . Это связано с тем, что многоклеточность требует кислорода , а до позднего неопротерозоя была очень ограничена доступность кислорода . [ 12 ] После таяния « Земли-снежка » в середине неопротерозоя питательные вещества, задержанные льдом, затопили океаны. [ 8 ] Выжившие бактерии процветали благодаря повышенному уровню питательных веществ. Среди этих микробов были цианобактерии и другие кислород , производящие бактерии , что привело к массовому повышению уровня кислорода. Повышенная доступность кислорода позволила клеткам использовать его для производства коллагена. Коллаген является ключевым компонентом агрегации клеток. Это веревочная молекула, которая «связывает» клетки вместе. Кислород необходим для коллагена синтеза , поскольку аскорбиновая кислота ( витамин С ) необходима для этого процесса. [ 6 ] Ключевым компонентом молекулы аскорбиновой кислоты является кислород (химическая формула C 6 H 8 O 6 ). [ 27 ] Таким образом, очевидно, что увеличение содержания кислорода является решающим шагом на пути к развитию многоклеточности , поскольку он необходим для синтеза коллагена. [ 8 ]
Строительные блоки найдены как у губок, так и у людей
[ редактировать ]Коллаген
[ редактировать ]Коллаген является наиболее распространенным белком у млекопитающих и играет важную роль в формировании костей, кожи и других соединительных тканей. Различные типы коллагена обнаружены у всех многоклеточных организмов, включая губки.
Было обнаружено, что у губок действительно имеется последовательность генов, кодирующая коллаген типа IV, который является диагностическим признаком базальной пластинки . [ 28 ]
Также было обнаружено, что 29 типов коллагена у человека существует . Эту обширную группу можно далее разделить на несколько семейств в зависимости от их первичной структуры и супрамолекулярной организации. Среди многих типов коллагенов только фибриллярный коллаген и коллаген базальной мембраны (тип IV) были обнаружены у губок и книдарий, которые являются двумя самыми ранними ветвящимися линиями многоклеточных животных. Исследования были сосредоточены на происхождении фибриллярных молекул коллагена. У губок существуют три клады фибриллярных молекул: A, B и C. Предполагается, что только фибриллярные коллагены клады B сохранили свою характерную модульную структуру от губки до человека. [ 29 ]
У млекопитающих фибриллярные коллагены, участвующие в образовании поперечно- исчерченных фибрилл, относятся к типам I–III, V и XI. Коллагены типа II и типа XI составляют фибриллы , присутствующие в хряще . Их можно отличить от коллагенов, расположенных в нехрящевых тканях, к которым относятся коллагены типов I, III и V. [ 29 ]
Белок
[ редактировать ]Дополнительные исследования белков губок показали, что из 42 проанализированных белков губок все они имели гомологичные белки, обнаруженные у человека. Сходству белков губки и человека присвоен показатель идентичности 53% по сравнению с показателем 42% при сравнении той же последовательности с последовательностью C. elegans . [ 30 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д Валентин, JW (1994). «Позднедокембрийские билатерии: степени и клады» . Труды Национальной академии наук . 91 (15): 6751–6757. Бибкод : 1994PNAS...91.6751V . дои : 10.1073/pnas.91.15.6751 . ПМЦ 44279 . ПМИД 8041693 .
- ^ «Стратиграфическая схема 2022» (PDF) . Международная стратиграфическая комиссия. февраль 2022 года . Проверено 25 апреля 2022 г.
- ^ Jump up to: а б с д Эрвин, Дуглас; Валентин, Джеймс; Яблонски, Дэвид (1997). «Происхождение строения тела: недавние находки окаменелостей и новое понимание развития животных открывают новый взгляд на загадку стремительного развития животных в раннем кембрии» . Американский учёный . 85 : 126–137.
- ^ Jump up to: а б с Нарбонн, Гай; Гелинг, Джеймс (2003). «Жизнь после снежного кома: древнейшие сложные эдиакарские окаменелости» (PDF) . Геология . 31 (1): 27–30. Бибкод : 2003Geo....31...27N . doi : 10.1130/0091-7613(2003)031<0027:lastoc>2.0.co;2 . Архивировано из оригинала (PDF) 31 октября 2004 г.
- ^ Jump up to: а б с д и ж Глесснер, Мартин (1959). «Самые старые ископаемые фауны Южной Австралии» (PDF) . Геологическое обозрение . 47 (2): 522–531. Бибкод : 1959ГеоРу..47..522Г . дои : 10.1007/bf01800671 . S2CID 140615593 .
- ^ Jump up to: а б Тау, Кеннет (1970). «Кислородно-коллагеновый приоритет и окаменелости ранних многоклеточных организмов» . Труды Национальной академии наук . 65 (4): 781–788. Бибкод : 1970ПНАС...65..781Т . дои : 10.1073/pnas.65.4.781 . ПМК 282983 . ПМИД 5266150 .
- ^ Jump up to: а б Глесснер, Мартин (1969). «Следы окаменелостей докембрия и базального кембрия» . Летайя . 2 (4): 369–393. дои : 10.1111/j.1502-3931.1969.tb01258.x .
- ^ Jump up to: а б с д и ж г Аттенборо, Дэвид. «Первая жизнь» . Ютуб .
- ^ Антклифф, Джонатан; Бразье, Мартин (2008). «Чарния в 50 лет: модели развития эдиакарских листьев» . Палеонтология . 51 (1): 1475–4983. Бибкод : 2008Palgy..51...11A . дои : 10.1111/j.1475-4983.2007.00738.x . S2CID 83486435 .
- ^ Jump up to: а б Реталлак, Грегори (2007). «Рост, распад и уплотнение захоронений дикинсонии, культового эдиакарского ископаемого» . Алчеринга . 31 (3): 215–240. дои : 10.1080/03115510701484705 . S2CID 17181699 .
- ^ Сперлинг, Эрик; Винтер, Якоб (2010). «Сродство плакозой к дикинсонии и эволюция способов питания многоклеточных животных позднего протерозоя». Эволюция и развитие . 12 (2): 201–209. дои : 10.1111/j.1525-142X.2010.00404.x . ПМИД 20433459 . S2CID 38559058 .
- ^ Jump up to: а б с д Кинг, Николь (сентябрь 2004 г.). «Одноклеточное происхождение развития животных» (PDF) . Развивающая клетка . 7 (3): 313–325. дои : 10.1016/j.devcel.2004.08.010 . ПМИД 15363407 .
- ^ Jump up to: а б с д и Король, Николь; и др. (2008). «Геном хоанофлагелляты Monosiga brevicollis и происхождение многоклеточных животных» . Природа . 451 (7180): 783–788. Бибкод : 2008Natur.451..783K . дои : 10.1038/nature06617 . ПМК 2562698 . ПМИД 18273011 .
- ^ Король, Николь; Кэррол, Шон Б. (10 сентября 2001 г.). «Рецепторная тирозинкиназа из хоанофлагеллят: молекулярное понимание ранней эволюции животных» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (26): 15032–15037. Бибкод : 2001PNAS...9815032K . дои : 10.1073/pnas.261477698 . ПМК 64978 . ПМИД 11752452 .
- ^ Jump up to: а б с Ваггонер, Бен. «Знакомство с хоанофлагеллатами: с чего все началось для животных» . UCMP Беркли . Беркли.
- ^ Jump up to: а б с д Дайел, Марк; Алегадо, Розанна; Фэрклаф, Стивен; Левин, Тера; Николс, Скотт; Макдональд, Кент; Кинг, Николь (сентябрь 2011 г.). «Дифференцировка клеток и морфогенез в колониеобразующей хоанофлагелляте Salpingoeca Rosetta» . Биология развития . 357 (1): 73–82. дои : 10.1016/j.ydbio.2011.06.003 . ПМК 3156392 . ПМИД 21699890 .
- ^ Jump up to: а б Ли, Цзя-Вэй; Чен, Цзюнь-Юань; Хуа, Цзы-Эн (февраль 1998 г.). «Докембрийские губки с ячеистым строением». Наука . 279 (5352): 879–882. Бибкод : 1998Sci...279..879L . дои : 10.1126/science.279.5352.879 . ПМИД 9452391 .
- ^ Гелинг, Джеймс; Ригби, Кейт (1996). «Долгожданные губки из неопротерозойской эдиакарской фауны Южной Австралии». Журнал палеонтологии . 70 (2): 185–195. дои : 10.1017/S0022336000023283 . JSTOR 1306383 . S2CID 130802211 .
- ^ Jump up to: а б Коллинз, Аллен Г.; Ваггонер, Бен. «Porifera: Еще о морфологии» . UCMP Беркли . Беркли.
- ^ Дюпон, Самуэль; Корре, Эрван; Ли, Яньян; Васелет, Жан; Бурге-Кондрацкий, Мари-Лиза (декабрь 2013 г.). «Первые сведения о микробиоме плотоядной губки» . ФЭМС Микробиология Экология . 86 (3): 520–531. дои : 10.1111/1574-6941.12178 . ПМИД 23845054 .
- ^ Jump up to: а б с д и ж г час я дж Коллинз, Аллен Г. «Porifera: Клетки» . UCMP Беркли . Беркли.
- ^ Ларру, Клэр; и др. (2006). «Эволюционная экспрессия генов факторов транскрипции у демогубки: понимание происхождения многоклеточности многоклеточных животных» . Эволюция и развитие . 8 (2): 150–173. дои : 10.1111/j.1525-142x.2006.00086.x . ПМИД 16509894 . S2CID 36836333 .
- ^ Майерс, Фил. «Губки Porifera» . Сеть разнообразия животных .
- ^ Иваса, Джанет; Шостак, Джек. «Хронология эволюции жизни» . Исследование истоков жизни .
- ^ Jump up to: а б Букхардт, Павел; и др. (2 августа 2011 г.). «Первичный нейросекреторный аппарат, выявленный у хоанофлагелляты Monosiga brevicollis» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (37): 15264–15269. Бибкод : 2011PNAS..10815264B . дои : 10.1073/pnas.1106189108 . ПМК 3174607 . ПМИД 21876177 .
- ^ Jump up to: а б Бораас, Мартин; Сил, Дайан; Боксхорн, Джозеф (1998). «Фаготрофия жгутиконосцев выбирает колониальную добычу: возможное происхождение многоклеточности». Эволюционная экология . 12 (2): 153–164. дои : 10.1023/А:1006527528063 . S2CID 12298325 .
- ^ Найду, Ахилиндер (2003). «Витамин С в здоровье и болезнях человека до сих пор остается загадкой? Обзор» . Журнал питания . 2 (7): 7. дои : 10.1186/1475-2891-2-7 . ПМК 201008 . ПМИД 14498993 .
- ^ Тайлер, Сет (2003). «Эпителий — основной строительный блок многоклеточных животных» . Интегративная и сравнительная биология . 43 (1): 55–63. дои : 10.1093/icb/43.1.55 . ПМИД 21680409 .
- ^ Jump up to: а б Экспозито, Жан-Ив; Ларру, Клэр; Клюзель, Кэролайн; Валькур, Ульрих; Летиас, Клэр; Дегнан, Бернар (2008). «Разнообразие фибриллярного коллагена демоспонги и морских анемонов свидетельствует о раннем появлении клад A/C и сохранении модульной структуры коллагенов типа V/XI от губки до человека» . Журнал биологической химии . 283 (42): 28226–28235. дои : 10.1074/jbc.M804573200 . ПМЦ 2661393 . ПМИД 18697744 .
- ^ Гамулин, Вера; Мюллер, Изабель; Мюллер, Вернер (2008). «Белки губок больше похожи на белки Homo sapiens, чем на Caenorhabditis elegans» . Биологический журнал Линнеевского общества . 71 (4): 821–828. дои : 10.1111/j.1095-8312.2000.tb01293.x .