Человеческие рудименты

В контексте эволюции человеческой рудиментарность человека включает в себя те черты, которые наблюдаются у людей , которые утратили все или большую часть своих первоначальных функций в ходе эволюции . Хотя структуры, называемые рудиментарными , часто кажутся нефункциональными, рудиментарная структура может сохранять меньшие функции или развивать новые, незначительные. В некоторых случаях структуры, которые когда-то считались рудиментарными, просто выполняли нераспознанную функцию. Рудиментарные органы иногда называют рудиментарными органами . [1] Многие человеческие характеристики также рудиментарны у других приматов и родственных им животных.
История [ править ]
Чарльз Дарвин перечислил ряд предполагаемых рудиментарных особенностей человека, которые он назвал рудиментарными, в книге «Происхождение человека» (1871). К ним относятся мышцы уха ; зубы мудрости ; приложение ; копчик ; волосы на теле ; и полулунная складка в уголке глаза . Дарвин также отметил спорадический характер многих рудиментарных особенностей, особенно мускулатуры. Ссылаясь на работы анатома Уильяма Тернера , Дарвин выделил ряд спорадических мышц, которые он идентифицировал как рудиментарные остатки panniculus carnosus , особенно грудинную мышцу . [2] [3]
В 1893 году Роберт Видерсхайм опубликовал «Структуру человека» — книгу об анатомии человека и ее значении для истории эволюции человека. В этой книге содержался список из 86 человеческих органов, которые он считал рудиментарными, или, как объяснял сам Видерсхайм: «Органы стали полностью или частично нефункциональными, некоторые появляются только в эмбрионе, другие присутствуют в течение жизни постоянно или непостоянно. По большей части органы который можно по праву назвать Рудиментарным». [4] Его список предположительно рудиментарных органов включал многие примеры на этой странице, а также другие, которые тогда ошибочно считались чисто рудиментарными, такие как шишковидная железа , вилочковая железа и гипофиз . Некоторые из этих органов, утратившие свои очевидные, первоначальные функции, впоследствии, как выяснилось, сохранили функции, которые оставались непризнанными до открытия гормонов или многих функций и тканей иммунной системы. [5] [6] Примеры включали:
- роль шишковидной железы в регуляции циркадного ритма (ни функция, ни даже существование мелатонина еще не были известны);
- открытие роли тимуса в иммунной системе отложено на многие десятилетия вперед; он оставался загадочным органом до середины 20 века;
- гипофиз с их многочисленными и разнообразными гормонами были далеки от понимания, не говоря уже о и гипоталамус сложности их взаимоотношений.
Исторически существовала тенденция не только считать аппендикс бесполезным рудиментарным явлением, но и считать его анатомической опасностью, предрасположенностью к опасному воспалению . Еще в середине 20-го века многие авторитетные авторитеты не признавали за ним никакой полезной функции. [7] Эту точку зрения поддержал или, возможно, вдохновил сам Дарвин в издании 1874 года его книги « Происхождение человека и отбор в отношении пола» . Заявленная склонность органа к аппендициту и его плохо изученная роль привели к тому, что аппендикс стал виноват в ряде, возможно, не связанных между собой состояний. Например, в 1916 году хирург заявил, что удаление аппендикса излечило несколько случаев невралгии тройничного нерва и других болей в области нервов головы и лица, хотя он заявил, что доказательства аппендицита у этих пациентов были неубедительными. [8] Открытие гормонов и гормональных принципов, особенно Бэйлисом и Старлингом , выступило против этих взглядов, но в начале двадцатого века предстояло провести множество фундаментальных исследований функций крупных частей пищеварительного тракта. В 1916 году автор счел необходимым выступить против идеи о том, что толстая кишка не выполняет важной функции и что «окончательное исчезновение аппендикса является скоординированным действием и не обязательно связано с такими частыми воспалениями, которые мы наблюдаем у человека». [9]
Подобные пренебрежительные взгляды уже давно вызывают сомнения. Примерно в 1920 году выдающийся хирург Кенелм Хатчинсон Дигби задокументировал предыдущие наблюдения, продолжавшиеся более тридцати лет, которые предполагали, что лимфатические ткани, такие как миндалины и аппендикс, могут выполнять существенные иммунологические функции.
Анатомический [ править ]
Приложение [ править ]

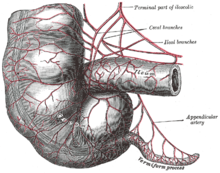
Когда-то считалось, что у современных людей аппендикс является пережитком избыточного органа, который у предковых видов выполнял пищеварительные функции, так же, как и до сих пор у современных видов, у которых кишечная флора гидролизует целлюлозу и подобные неперевариваемые растительные материалы. [10] Эта точка зрения изменилась за последние десятилетия, [11] исследования показывают, что аппендикс может служить важной цели. В частности, он может служить резервуаром для полезных кишечных бактерий . Одна из предлагаемых функций этого метода — позволить кишечным бактериям восстановиться в толстой кишке во время выздоровления от диареи или других заболеваний. [12]
У некоторых травоядных животных, таких как кролики, есть терминальный червеобразный отросток и слепая кишка , которые, по-видимому, несут участки ткани с иммунными функциями и могут также играть важную роль в поддержании состава кишечной флоры . Однако, по-видимому, он не выполняет особой пищеварительной функции и присутствует не у всех травоядных, даже у тех, у кого большие слепые отростки. [13] Однако, как показано на прилагаемых изображениях, человеческий аппендикс обычно примерно сравним по размеру с аппендиксом кролика, хотя слепая кишка уменьшена до единственного выступа там, где подвздошная кишка впадает в толстую кишку. [7] У некоторых плотоядных животных также могут быть придатки, но они редко выходят за рамки рудиментарных придатков. [14] В соответствии с возможностью рудиментарных органов развивать новые функции, некоторые исследования показывают, что аппендикс может защищать от потери симбиотических бактерий, которые помогают в пищеварении, хотя это вряд ли будет новой функцией, учитывая наличие червеобразных отростков у многих травоядных животных. . [15] [16] Популяции кишечных бактерий, укоренившиеся в аппендиксе, могут способствовать быстрому восстановлению флоры толстой кишки после болезни, отравления или после того, как лечение антибиотиками истощает или иным образом вызывает вредные изменения в бактериальной популяции толстой кишки. [17]
Однако исследование 2013 года опровергает идею об обратной зависимости между размером слепой кишки, размером и наличием аппендикса. Он широко присутствует у euarchontoglires (надотряд млекопитающих, включающий грызунов, зайцеобразных и приматов), а также независимо эволюционировал у дипротодонтных сумчатых , однопроходных и весьма разнообразен по размеру и форме, что позволяет предположить, что он не является рудиментарным. Исследователи пришли к выводу, что аппендикс способен защищать полезные бактерии в кишечнике. Таким образом, когда на кишечник влияет приступ диареи или другое заболевание, вызывающее очистку кишечника, полезные бактерии в аппендиксе могут заново заселить пищеварительную систему и сохранить здоровье человека. [18]
Копчик [ править ]
Копчик , или копчик , является остатком утраченного хвоста . [19] У всех млекопитающих на определенном этапе развития есть хвост; у людей он присутствует в течение 4 недель на стадиях с 14 по 22 эмбриогенеза человека . [20] Этот хвост наиболее заметен у эмбрионов человека в возрасте 31–35 дней. [21] Копчик, расположенный в конце позвоночника, утратил свою первоначальную функцию обеспечения равновесия и подвижности, хотя он по-прежнему выполняет некоторые второстепенные функции, например, является точкой крепления мышц, что объясняет, почему он не деградировал дальше.
В редких случаях врожденный дефект приводит к тому, что короткая хвостообразная структура при рождении присутствует . С 1884 года в медицинской литературе сообщалось о 23 случаях рождения человеческих младенцев с такой структурой. [22] [23] В таких редких случаях, как этот, позвоночник и череп оказывались совершенно нормальными. Единственной аномалией был хвост длиной около двенадцати сантиметров. Эти хвосты, хотя и не имели никакого вредного воздействия, почти всегда удалялись хирургическим путем. [24]
Зубы мудрости [ править ]
Зубы мудрости — это рудиментарные третьи моляры, которые предки человека использовали для измельчения растительных тканей. Распространено мнение, что черепа предков человека имели более крупные челюсти с большим количеством зубов, которые, возможно, использовались для пережевывания листвы, чтобы компенсировать отсутствие способности эффективно переваривать целлюлозу, составляющую стенку растительной клетки. Поскольку рацион человека изменился, естественным образом были выбраны меньшие челюсти , однако третьи моляры, или «зубы мудрости», по-прежнему обычно развиваются во рту человека. [25]
Агенезия (неразвитие) зубов мудрости в человеческих популяциях колеблется от нуля у аборигенов Тасмании до почти 100% у коренных мексиканцев . [26] Разница связана с геном PAX9 (и, возможно, с другими генами). [27]
Вомероназальный орган [ править ]
У некоторых животных вомероназальный орган (ВНО) является частью второго, совершенно отдельного обоняния, известного как добавочная обонятельная система . Было проведено множество исследований, чтобы выяснить, действительно ли существует ВНО у взрослых людей. Тротье и др. [28] подсчитали, что около 92% их пациентов, не перенесших операцию на перегородке, имели по крайней мере один неповрежденный ВНО. Кьяер и Фишер Хансен, с другой стороны, [29] заявили, что структура VNO исчезла во время внутриутробного развития, как это происходит у некоторых приматов. [30] Однако Смит и Бхатнагар (2000) [31] утверждали, что Кьяер и Фишер Хансен просто упустили структуру у более старых плодов. Вон (2000) обнаружил признаки ВНО у 13 из 22 трупов (59,1%) и у 22 из 78 живых пациентов (28,2%). [32] Учитывая эти результаты, некоторые ученые утверждают, что у взрослых людей существует ВНО. [33] [34] Однако большинство исследователей стремились идентифицировать отверстие вомероназального органа у человека, а не идентифицировать саму трубчатую эпителиальную структуру. [35] Таким образом, утверждалось, что такие исследования с использованием методов макроскопического наблюдения иногда пропускали или даже неправильно идентифицировали сошниково-носовой орган. [36]
Среди исследований, в которых используются микроанатомические методы, нет сообщений о том, что у людей есть активные сенсорные нейроны, подобные тем, которые находятся в рабочих вомероназальных системах других животных. [36] [37] Более того, на сегодняшний день нет доказательств того, что между любыми существующими сенсорными рецепторными клетками, которые могут находиться в ВНО взрослого человека и мозге, существуют нервные и аксонные связи. [38] Аналогичным образом, нет никаких доказательств наличия каких-либо дополнительных обонятельных луковиц у взрослых людей. [36] а ключевые гены, участвующие в функции VNO у других млекопитающих, стали псевдогенами у человека. Таким образом, хотя наличие структуры у взрослых людей обсуждается, обзор научной литературы, проведенный Тристрамом Вяттом, пришел к выводу, что «большинство специалистов в этой области... скептически относятся к вероятности функционального ВНО у взрослых людей на основании имеющихся данных». ." [39]
Ухо [ править ]


Уши . макаки и большинства других обезьян имеют гораздо более развитые мышцы , чем у людей, и поэтому обладают способностью двигать ушами, чтобы лучше слышать потенциальные угрозы [40] людей и других приматов, таких как орангутан и шимпанзе, Однако у ушные мышцы минимально развиты и нефункциональны, но все же достаточно велики, чтобы их можно было идентифицировать. [10] О мышце, прикрепленной к уху, которая по какой-либо причине не может двигать ухом, больше нельзя сказать, что она выполняет какую-либо биологическую функцию. У людей эти мышцы вариабельны, так что некоторые люди могут двигать ушами в разных направлениях, а другие могут добиться такого движения путем повторных попыток. [10] [41] У таких приматов неспособность двигать ухом компенсируется главным образом способностью поворачивать голову в горизонтальной плоскости, способностью, не свойственной большинству обезьян — функция, обеспечиваемая одной структурой, теперь заменяется другой. [42]
Внешняя структура уха также имеет некоторые рудиментарные особенности, такие как узел или точка на завитке уха, известная как бугорок Дарвина , который встречается примерно у 10% населения.
Глаз [ править ]
Полулунная складка — небольшая складка ткани во внутреннем углу глаза. Это рудиментарный остаток мигательной перепонки , то есть третьего века, органа, полностью функционального у некоторых других видов млекопитающих. [43] Связанные с ним мышцы также рудиментарны. [10] Известно, что только один вид приматов , Calabar angwantibo , имеет функционирующую мигательную перепонку. [44]
Орбитальная мышца — рудиментарная или рудиментарная бесполосистая мышца (гладкая мышца) глаза, пересекающая подглазничную борозду и клиновидно-челюстную щель и интимно слитная с надкостницей глазницы. Он был описан Йоханнесом Петером Мюллером и часто называется мышцей Мюллера. Мышца образует важную часть латеральной стенки глазницы у некоторых животных, но у человека неизвестно, выполняет ли она какую-либо значительную функцию. [45] [46]
Репродуктивная система [ править ]
Гениталии [ править ]
Во внутренних половых органах каждого пола человека в период эмбрионального развития имеются остаточные органы мезонефрических и парамезонефральных протоков:
Рудиментарные структуры человека также включают в себя остатки эмбрионального развития, которые когда-то выполняли определенную функцию во время развития, такие как пупок, и аналогичные структуры между биологическими полами. Например, мужчины также рождаются с двумя сосками, которые, как известно, не выполняют никакой функции по сравнению с женщинами. [47] Что касается развития мочеполовой системы, то как внутренние, так и наружные половые органы плодов мужского и женского пола обладают способностью полностью или частично формировать аналогичный фенотип противоположного биологического пола при воздействии недостатка/избытка андрогенов или гена SRY во время внутриутробного развития. [48] [49] Примеры рудиментарных остатков развития мочеполовой системы включают девственную плеву , которая представляет собой мембрану , которая окружает или частично покрывает наружное отверстие влагалища , происходящее из бугорка пазухи во время развития плода и гомологичное мужским семенным бугоркам . [50] Некоторые исследователи [ ВОЗ? ] выдвинули гипотезу, что сохранение девственной плевы может обеспечивать временную защиту от инфекции , поскольку она отделяет просвет влагалища от полости мочеполового синуса во время развития. [51] Другие примеры включают головку полового члена и клитор , малые половые губы и вентральную часть полового члена, а также фолликулы яичников и семенные канальцы. [50]
В наше время ведутся споры относительно того, является ли крайняя плоть жизненно важной или рудиментарной структурой. [52] В 1949 году британский врач Дуглас Гейрднер отметил, что крайняя плоть играет важную защитную роль у новорожденных. Он писал: «Часто утверждают, что крайняя плоть представляет собой рудиментарную структуру, лишенную функции... Однако, по-видимому, не случайно, что в годы недержания у ребенка головка полового члена полностью покрыта крайней плотью, ибо, лишенная Благодаря этой защите головка становится уязвимой к травмам от контакта с мокрой одеждой или салфеткой». [52] Во время физического полового акта крайняя плоть уменьшает трение, что может снизить потребность в дополнительных источниках смазки. [52] «Однако некоторые медицинские исследователи утверждают, что обрезанные мужчины прекрасно получают удовольствие от секса, и что, учитывая недавние исследования по передаче ВИЧ, крайняя плоть причиняет больше проблем, чем пользы». [52] Площадь наружной крайней плоти составляет от 7 до 100 см. 2 , [53] а внутренняя крайняя плоть имеет размеры от 18 до 68 см. 2 , [54] что представляет собой широкий диапазон. Что касается рудиментарных структур, Чарльз Дарвин писал: «Орган, ставший бесполезным, вполне может быть изменчивым, поскольку его вариации не могут быть проверены естественным отбором». [55]
Мускулатура [ править ]
Считается, что ряд мышц в человеческом теле являются рудиментарными либо из-за того, что они значительно уменьшены в размерах по сравнению с гомологичными мышцами у других видов, либо из-за того, что они стали преимущественно сухожильными, либо из-за того, что их частота сильно варьируется внутри или между популяциями.
Глава [ править ]
Малая затылочная мышца — это мышца в задней части головы, которая обычно присоединяется к ушным мышцам уха. Частота этой мышцы очень спорадична: она всегда присутствует у малайцев, присутствует у 56% африканцев, 50% японцев и 36% европейцев и отсутствует у народа койхой на юго-западе Африки и у меланезийцев . [56] Частота других мелких мышц головы, связанных с затылочной областью и заушным мышечным комплексом, часто варьирует. [57]
Платизма , четырехугольная ( четырехсторонняя ) мышца листовидной конфигурации, является рудиментарным остатком панникулы карноза животных. У лошадей именно мышцы позволяют им сбрасывать муху со спины. [ нужна ссылка ]
Лицо [ править ]
У многих низших животных верхняя губа и область пазух связаны с усами или вибриссами, которые выполняют сенсорную функцию. У людей этих усов не существует, но все еще есть спорадические случаи, когда можно обнаружить элементы связанных с ними вибриссальных капсульных мышц или волосяных мышц пазух. На основании гистологических исследований верхних губ 20 трупов Tamatsu et al. обнаружили, что структуры, напоминающие такие мышцы, присутствовали у 35% (7/20) их образцов. [58]
Рука [ править ]
Длинная ладонная мышца рассматривается как небольшое сухожилие между лучевым сгибателем запястья и локтевым сгибателем запястья , хотя оно присутствует не всегда. Мышцы отсутствуют примерно у 14% населения, однако это сильно зависит от этнической принадлежности. Считается, что эта мышца активно участвовала в древесном передвижении приматов, но в настоящее время не выполняет никакой функции, поскольку не обеспечивает большей силы хвата. [59] Одно исследование показало, что распространенность агенезии длинной ладонной мышцы у 500 индийских пациентов составляет 17,2% (8% двусторонняя и 9,2% односторонняя). [60] Ладонная мышца является популярным источником сухожильного материала для трансплантатов, и это побудило исследования, которые показали, что отсутствие ладонной мышцы не оказывает заметного влияния на силу захвата. [61]
Мышца , поднимающая ключицу, в заднем треугольнике шеи — избыточная мышца, присутствующая только у 2–3% всех людей. [62] но почти всегда присутствует у большинства видов млекопитающих, включая гиббонов и орангутанов . [63]
Торс [ править ]
Пирамидная мышца живота представляет собой небольшую треугольную мышцу, расположенную впереди прямой мышцы живота и содержащуюся в влагалище прямой мышцы живота . Он отсутствует у 20% людей, а при его отсутствии нижний конец прямой мышцы пропорционально увеличивается в размерах. Анатомические исследования показывают, что силы, создаваемые пирамидальными мышцами, относительно невелики. [64]
Широчайшая мышца спины имеет несколько спорадических разновидностей . Одним из конкретных вариантов является наличие дорсоэпитрохлеарной или широчайше-мыщелковой мышцы, которая представляет собой мышцу, идущую от сухожилия широчайшей мышцы спины к длинной головке трехглавой мышцы плеча . Он примечателен своим хорошо развитым характером у других человекообразных обезьян и мартышек, где он является важной мышцей для лазания, а именно dorsoepitrochlearis brachii. [65] [66] Эта мышца имеется у ≈5% людей. [67]
Нога [ править ]
состоит Подошвенная мышца из тонкого мышечного брюшка и длинного тонкого сухожилия. Мышечное брюшко имеет длину примерно 5–10 сантиметров (2–4 дюйма) и отсутствует у 7–10% населения. Он имеет некоторые слабые функции при перемещении колена и лодыжки, но обычно считается избыточным и часто используется в качестве источника сухожилия для трансплантатов. Длинное тонкое сухожилие подошвенной мышцы в шутку называют «нервом первокурсника», поскольку студенты-медики часто принимают его за нерв.
Язык [ править ]
Еще одним примером рудиментарности человека является язык, в частности хондроязычная мышца . При морфологическом исследовании 100 японских трупов было обнаружено, что 86% идентифицированных волокон были твердыми и связаны соответствующим образом, чтобы облегчить речь и жевание. Остальные 14% волокон были короткими, тонкими и редкими – почти бесполезными, поэтому был сделан вывод о том, что они имеют рудиментарное происхождение. [68]
Грудь [ править ]
Дополнительные соски или груди иногда появляются по линии молочных желез у людей, являясь остатками предков млекопитающих, у которых было более двух сосков или грудей. [69] [70] Один отчет 2021 года продемонстрировал, что у всех здоровых молодых мужчин и женщин, участвовавших в анатомическом исследовании передней поверхности тела, было обнаружено 8 пар очаговых жировых холмиков, идущих вдоль эмбриологических гребней молочной железы от подмышек до верхней внутренней поверхности бедер. Они всегда располагались в одних и тех же относительных анатомических местах – аналогично участкам груди у других плацентарных млекопитающих – и часто имели соскообразные родинки или дополнительные волосы, расположенные на вершинах холмиков. Следовательно, очаговые жировые выступы на передней части туловища человека, вероятно, представляют собой цепочки рудиментарных грудей, состоящих из первичного грудного жира. [71]
Поведенческий [ править ]
Люди также имеют некоторые рудиментарные модели поведения и рефлексы. [72]
Мурашки по коже [ править ]

Образование мурашек у человека в условиях стресса является рудиментарным рефлексом ; Возможная функция у далеких эволюционных предков человечества заключалась в том, чтобы поднимать волосы на теле, заставляя предка казаться крупнее и отпугивая хищников. [73] [72] Поднятие шерсти также используется для улавливания дополнительного слоя воздуха, сохраняя животное в тепле. [72] Из-за уменьшения количества волос у людей рефлекторное образование мурашек на холоде также является рудиментарным. [72]
Ладонный хватательный рефлекс [ править ]
Ладонный хватательный рефлекс считается рудиментарным поведением у младенцев. Приложив палец или предмет к ладони младенца, он надежно схватит его. Эта хватка оказывается довольно сильной. [74] Некоторые младенцы — 37% по данным исследования 1932 года — способны поддерживать свой вес с помощью стержня. [75] хотя они никак не могут цепляться за свою мать. Хватка также заметна в ногах. Когда ребенок садится, его цепкие ступни принимают свернувшуюся позу, подобную той, которая наблюдается у взрослого шимпанзе. [76] [77] У предков приматов было достаточно волос на теле, за которые младенец мог цепляться, в отличие от современных людей, что позволяло их матери избежать опасности, например, залезть на дерево в присутствии хищника, не занимая свои руки, держащие ребенка. .
Иккинг [ править ]
Было высказано предположение, что икота является эволюционным остатком более раннего земноводных дыхания . [78] Земноводные, такие как головастики, глотают воздух и воду через жабры с помощью довольно простого двигательного рефлекса, похожего на икание млекопитающих. Двигательные пути, обеспечивающие икоту, формируются на ранних стадиях развития плода, до того, как формируются двигательные пути, обеспечивающие нормальную вентиляцию легких. Кроме того, икота и глотание амфибий подавляются повышенным уровнем CO 2 и могут быть остановлены ГАМК В агонистами рецептора , что иллюстрирует возможную общую физиологию и эволюционное наследие. Эти предположения могут объяснить, почему недоношенные дети проводят 2,5% своего времени, икая, возможно, сглатывая, как земноводные, поскольку их легкие еще не полностью сформированы. Внутриутробная икота плода бывает двух видов. Физиологический тип возникает до 28 недель после зачатия и обычно длится от пяти до десяти минут. Эта икота является частью развития плода и связана с миелинизацией диафрагмального нерва , который в первую очередь контролирует грудную диафрагму. Гипотеза филогении объясняет, как мог развиться рефлекс икоты, а если объяснения нет, то она может объяснить икоту как эволюционный пережиток, перешедший от наших предков-амфибий.Эта гипотеза была поставлена под сомнение из-за существования афферентной петли рефлекса, того факта, что она не объясняет причину закрытия голосовой щели, а также из-за того, что очень короткое сокращение икоты вряд ли окажет существенное усиливающее действие на медленную икоту. - подергивания дыхательных мышц. [ нужна ссылка ]
Псевдогены [ править ]
присутствует множество псевдогенов В геноме человека . Одним из примеров этого является L-гулонолактоноксидаза , ген, который функционален у большинства других млекопитающих и продуцирует фермент , синтезирующий витамин С. [79] У людей и других представителей подотряда Haplorrini мутация отключила этот ген и лишила его возможности производить фермент. Однако остатки гена все еще присутствуют в геноме человека. [80]
См. также [ править ]
Ссылки [ править ]
- ^ «Разница между рудиментарным и рудиментарным органом - Биология - Эволюция - 11741123 | Meritnation.com» . www.meritnation.com . Проверено 16 февраля 2021 г.
- ^ Дарвин С., Происхождение человека и отбор по признаку пола , Лондон: Джон Мюррей, 1890, стр.13. [1]
- ^ Тернер В., О грудинной мышце , Proc. Роял Соц. Эдинбургская сессия 1866–1867, стр.65. [2]
- ^ Видерсхайм, Р. (1893) Структура человека: указатель его прошлой истории . Второе издание. Перевод Х. и М. Бернаров. Лондон: Макмиллан и компания, 1895 г. [3]
- ^ Мюллер, Великобритания (2002). «Рудиментарные органы и структуры». В Пейгеле, Марк (ред.). Энциклопедия эволюции . Нью-Йорк: Издательство Оксфордского университета. стр. 1131–1133.
- ^ Керт-Бейкер, Мэгги (30 июля 2009 г.). «Рудиментарные органы не так уж и бесполезны» . Нэшнл Географик . Архивировано из оригинала 4 августа 2009 года . Проверено 27 июля 2013 г.
- ^ Jump up to: Перейти обратно: а б Уэллс, Х.Г. ; Хаксли, Дж .; Уэллс, врач общей практики (1929). Наука жизни . Касселс.
- ^ Розенталь, Мичиган: Журнал Американской медицинской ассоциации , том 67, выпуски 15–26, 1916. с. 1326
- ^ В. Колин Маккензи. «Вклад в биологию червеобразного аппендикса». Медицинская карта , том 89, с. 342, 1916 г.
- ^ Jump up to: Перейти обратно: а б с д Дарвин, Чарльз (1871). Происхождение человека и отбор по признаку пола . Джон Мюррей: Лондон.
- ^ Коой И.А., Сахами С., Мейер С.Л., Бускенс К.Дж., Те Вельде А.А. (октябрь 2016 г.). «Иммунология червеобразного отростка: обзор литературы» . Клиническая и экспериментальная иммунология . 186 (1): 1–9. дои : 10.1111/cei.12821 . ПМК 5011360 . ПМИД 27271818 .
- ^ Да, Дэниел Рен Йи; Луи, Рашид Н; Самол, Йенс; Нгеоу, Джоанн; Сун, Джозеф JY; Вонг, Санни Х. (февраль 2024 г.). «За пределами рудиментарного органа: влияние аппендикса на микробиом кишечника и колоректальный рак» . Журнал гастроэнтерологии и гепатологии . дои : 10.1111/jgh.16497 .
- ^ Стивенс, К. Эдвард; Хьюм, Ян (2004). Сравнительная физиология пищеварительной системы позвоночных . Кембридж: Издательство Кембриджского университета. ISBN 978-0-521-61714-7 .
- ^ Питер Роберт Чик, Эллен С. Диренфельд, Сравнительное питание и обмен веществ животных. Издательство: CABI; 2010 год ISBN 978-1-84593-631-0
- ^ «В конце концов, приложение может оказаться полезным – Здоровье – Здравоохранение – Больше новостей о здоровье – Новости NBC» . Новости Эн-Би-Си.
- ^ Рэндал Боллинджер, Р.; Барбас, Эндрю С.; Буш, Эррол Л.; Лин, Шу С.; Паркер, Уильям (2007). «Биопленки в толстой кишке предполагают очевидную функцию червеобразного отростка человека». Журнал теоретической биологии . 249 (4): 826–31. Бибкод : 2007JThBi.249..826R . дои : 10.1016/j.jtbi.2007.08.032 . ПМИД 17936308 .
- ^ Чарльз К. Чой, «Приложение: полезное и многообещающее», Live Science , 2009, Приложение имеет полезную функцию.
- ^ Смит, Х.Ф.; Фишер, Р.Э.; Эверетт, ML; Томас, AD; Рэндал Боллинджер, Р.; Паркер, В. (2009). «Сравнительная анатомия и филогенетическое распространение слепокишечного отростка млекопитающих» . Журнал эволюционной биологии . 22 (10): 1984–99. дои : 10.1111/j.1420-9101.2009.01809.x . ПМИД 19678866 .
- ^ Бисванат Мукхопадхьяй, Рам М. Шукла, Мадхумита Мукхопадьяй, Картик К. Мандал, Панкадж Халдар и Абхиджит Бенаре (2012), «Спектр человеческих хвостов: отчет о шести случаях», Журнал Индийской ассоциации детских хирургов , 17 (1 ): 23–25, doi : 10.4103/0971-9261.91082 , PMC 3263034 , PMID 22279360
{{citation}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сарага-Бабич, Мирна; Лехтонен, Ээро; Швайгер, Антон; Вартиоваара, Йорма (1994). «Морфологическая и иммуногистохимическая характеристика осевых структур переходного хвоста человека». Анналы анатомии - Анатомическая архитектура 176 (3): 277–86. дои : 10.1016/S0940-9602(11)80496-6 . ПМИД 8059973 .
- ^ Фэллон, Джон Ф.; Симандл, Б. Кей (1978). «Доказательства роли гибели клеток в исчезновении эмбрионального человеческого хвоста». Американский журнал анатомии . 152 (1): 111–29. дои : 10.1002/aja.1001520108 . ПМИД 677043 .
- ^ Дао, Ань Х.; Нетски, Мартин Г. (1984). «Человеческие хвосты и псевдохвосты». Патология человека . 15 (5): 449–53. дои : 10.1016/S0046-8177(84)80079-9 . ПМИД 6373560 .
- ^ Дуброу, Терри Дж.; Ваким, Филип Эшли; Лесавой, Малкольм А. (1988). «Детализация человеческого хвоста». Анналы пластической хирургии . 20 (4): 340–4. дои : 10.1097/00000637-198804000-00009 . ПМИД 3284435 .
- ^ Шпигельманн, Роберто; Шиндер, Эдгардо; Минц, Мордехай; Блакштейн, Александр (1985). «Человеческий хвост: доброкачественное клеймо». Журнал нейрохирургии . 63 (3): 461–2. дои : 10.3171/jns.1985.63.3.0461 . ПМИД 3894599 .
- ↑ Джонсон, доктор Джордж Б. «Доказательства эволюции». Архивировано 10 марта 2008 г. в Wayback Machine . (Страница 12) Txtwriter Inc., 8 июня 2006 г.
- ^ Розковцова, Э; Маркова, М; Долейси, Дж (1999). «Исследования агенезии третьих моляров среди популяций различного происхождения». Сборник Лекарского . 100 (2): 71–84. ПМИД 11220165 .
- ^ Перейра, ТВ; Сальзано, FM; Мостовская, А.; Тшечак, WH; Руис-Линарес, А.; Чиес, JAB; Сааведра, К.; Нагамачи, К.; Уртадо, AM; Хилл, К.; Кастро-Де-Гуэрра, Д.; Сильва-Джуниор, Вашингтон; Бортолини, М.-К. (2006). «Естественный отбор и молекулярная эволюция гена PAX9 приматов, основного детерминанта развития зубов» . Труды Национальной академии наук . 103 (15): 5676–81. Бибкод : 2006PNAS..103.5676P . дои : 10.1073/pnas.0509562103 . ПМЦ 1458632 . ПМИД 16585527 .
- ^ Тротье, Д.; Элойт, К; Вассеф, М; Талмейн, Дж; Бенсимон, Дж.Л.; Дёвинг, КБ; Ферран, Дж (2000). «Вомероназальная полость у взрослых людей» . Химические чувства . 25 (4): 369–80. дои : 10.1093/chemse/25.4.369 . ПМИД 10944499 .
- ^ Кьер, Ингер; Хансен, Биргит Фишер (1996). «Вомероназальный орган человека: пренатальные стадии развития и распределение рилизинг-гормона лютеинизирующего гормона». Европейский журнал устных наук . 104 (1): 34–40. дои : 10.1111/j.1600-0722.1996.tb00043.x . ПМИД 8653495 .
- ^ Смит, Тимоти Д.; Сигел, Майкл И.; Бхатнагар, Кунвар П. (2001). «Переоценка вомероназальной системы катарных приматов: онтогенез, морфология, функциональность и сохраняющиеся вопросы» . Анатомическая запись . 265 (4): 176–192. дои : 10.1002/ar.1152 . ПМИД 11519019 .
- ^ Смит, Тимоти Д.; Бхатнагар, Кунвар П. (2000). «Вомероназальный орган человека. Часть II: Пренатальное развитие» . Журнал анатомии . 197 (3): 421–436. дои : 10.1046/j.1469-7580.2000.19730421.x . ПМЦ 1468143 . ПМИД 11117628 .
- ^ Вон, Дж; Майр, Э.А.; Болджер, МЫ; Конран, RM (2000). «Вомероназальный орган: объективный анатомический анализ его распространенности» . Журнал «Ухо, нос и горло» . 79 (8): 600–605. дои : 10.1177/014556130007900814 . ПМИД 10969469 .
- ^ Джонсон, А.; Джозефсон, Р.; Хоук, М. (1985). «Клинические и гистологические доказательства наличия вомероназального (якобсоновского) органа у взрослых людей». Журнал отоларингологии . 14 (2): 71–79. ПМИД 4068105 .
- ^ Фолтан, Рене; Шеды, Иржи (2009). «Поведенческие изменения больных после ортогнатической операции развиваются на основе утраты сошниково-носового органа: гипотеза» . Медицина головы и лица . 5 :5. дои : 10.1186/1746-160X-5-5 . ПМЦ 2653472 . ПМИД 19161592 .
- ^ Бхатнагар, Кунвар П.; Смит, Тимоти Д. (2001). «Вомероназальный орган человека. III. Постнатальное развитие от младенчества до девятого десятилетия» . Журнал анатомии . 199 (Часть 3): 289–302. дои : 10.1046/j.1469-7580.2001.19930289.x . ПМЦ 1468331 . ПМИД 11554506 .
- ^ Jump up to: Перейти обратно: а б с Бхатнагар, Кунвар П.; Кеннеди, Рэй С.; Барон, Георг; Гринберг, Ричард А. (1987). «Количество митральных клеток и объем луковицы в стареющей обонятельной луковице человека: количественное морфологическое исследование». Анатомическая запись . 218 (1): 73–87. дои : 10.1002/ar.1092180112 . ПМИД 3605663 . S2CID 25630359 .
- ^ Витт, М.; Хаммел, Т. (2006). Вомероназальный и обонятельный эпителий: существует ли клеточная основа для вомероназального восприятия человека? . Международный обзор цитологии. Том. 248. С. 209–259. дои : 10.1016/S0074-7696(06)48004-9 . ISBN 978-0-12-364652-1 . ПМИД 16487792 .
- ^ Высоцкий CJ, Прети Дж (ноябрь 2004 г.). «Факты, заблуждения, страхи и разочарования по поводу человеческих феромонов» . Анатомические записи, часть A: открытия в молекулярной, клеточной и эволюционной биологии . 281 (1): 1201–1211. дои : 10.1002/ar.a.20125 . ПМИД 15470677 .
- ^ Вятт, Тристрам Д. (2003). Феромоны и поведение животных: общение посредством запаха и вкуса . Кембридж: Издательство Кембриджского университета. п. 295 . ISBN 978-0-521-48526-5 .
- ^ Проф. А. Макалистер, Анналы и журнал естественной истории, том. VII., 1871, с. 342.
- ^ Баир, Дж. Х. (1901). «Развитие произвольного контроля» . Психологический обзор . 8 (5): 474–510. дои : 10.1037/h0074157 . hdl : 2027/mdp.39015070189314 .
- ^ Г-н Сент-Джордж Миварт, Элементарная анатомия, 1873, с. 396.
- ^ Оуэн, Р. 1866–1868. Сравнительная анатомия и физиология позвоночных . Лондон. [ нужна страница ]
- ^ Монтанья, В.; Мачида, Х.; Перкинс, Э.М. (1966). «Кожа приматов. XXXIII. Кожа ангвантибо (Arctocebus Calabarensis)». Американский журнал физической антропологии . 25 (3): 277–90. дои : 10.1002/ajpa.1330250307 . ПМИД 5971502 .
- ^ Ториен, MJ; Гоус, А.Е. (1978). «Орбитальная мышца Мюллера». Южноафриканский медицинский журнал . 53 (4): 139–41. ПМИД 653491 .
- ^ Даттон, Дж. Дж., Атлас клинической и хирургической анатомии орбиты, 2-е издание , Elsevier, 2011. стр. 116-117.
- ^ «Анатомия и эмбриология груди». Основы пластической хирургии (2015): 355–361.
- ^ Хаджиатанасиу, CG; Браунер, Р.; Лортат-Джейкоб, С.; Нивот, С.; Жобер, Ф.; Феллоус, М.; Ниуль-Фекете, К.; Раппапорт, Р. (1994). «Истинный гермафродитизм: генетические варианты и клиническое лечение». Журнал педиатрии . 125 (5): 738–744. дои : 10.1016/S0022-3476(06)80172-1 . ПМИД 7965425 .
- ^ Эрен, Эрдал; Эдгюнлю, Туба; Асут, Эмре; Каракаш Челик, Севим (2016). «Гомозиготная мутация Ala65Pro с полиморфизмом V89L при дефиците SRD5A2» . Журнал клинических исследований в детской эндокринологии . 8 (2): 218–223. дои : 10.4274/jcrpe.2495 . ПМК 5096479 . ПМИД 26761946 .
- ^ Jump up to: Перейти обратно: а б Хили, Эндрю (2010). «Эмбриология женской репродуктивной системы». Визуализация гинекологических заболеваний у младенцев и детей . Медицинская радиология. стр. 21–30. дои : 10.1007/174_2010_128 . ISBN 978-3-540-85601-6 .
- ^ Басаран, Мустафа; Усал, Дениз; Айдемир, Джумхур (2009). «Хирургия по сохранению девственной плевы при неперфорированной девственной плеве: отчеты о случаях заболевания и обзор литературы». Журнал детской и подростковой гинекологии . 22 (4): с61–64. дои : 10.1016/j.jpag.2008.03.009 . ПМИД 19646660 .
- ^ Jump up to: Перейти обратно: а б с д Кольер, Роджер (22 ноября 2011 г.). «Жизненная или рудиментарная? У крайней плоти есть свои поклонники и враги» . CMAJ . 183 (17): 1963–1964. дои : 10.1503/cmaj.109-4014 . ISSN 0820-3946 . ПМК 3225416 . ПМИД 22025652 .
- ^ Кигози Г., Вавер М., Ссеттуба А. и др. «Площадь крайней плоти и заражение ВИЧ в Ракаи, Уганда (размер имеет значение)» СПИД 2009 г.; 23 (16): 2209–2213. http://dx.doi.org/10.1097/QAD.0b013e328330ed8.
- ^ Веркер PMN, Тернг ASC, Кон М. «Свободный лоскут крайней плоти: технико-экономическое обоснование рассечения и клиническое применение сверхтонкого нового лоскута». Пластическая реконструктивная хирургия . 1998 год; 102 (4): 1075–1082. 10.1097/00006534-199809020-00024.
- ^ Дарвин К. Происхождение видов посредством естественного отбора . Лондон, Великобритания: Джон Мюррей; 1859.
- ^ Макалистер, Александр (1875). «Дополнительные наблюдения за мышечными аномалиями в анатомии человека. (Третья серия) с каталогом основных мышечных вариаций, опубликованных до сих пор». Труды Королевской ирландской академии . 25 : 1–134. JSTOR 30079154 .
- ^ Герра, Альдо Бенджамин; Метцингер, Стивен Эрик; Метцингер, Ребекка Кроуфорд; Се, Чен; Се, Юэ; Ригби, Питер Листер; Наугл, Томас (2004). «Вариабельность постаурикулярного мышечного комплекса» . Архив пластической хирургии лица . 6 (5): 342–7. дои : 10.1001/archfaci.6.5.342 . ПМИД 15381582 .
- ^ Тамацу, Юичи; Цукахара, Казуэ; Хотта, Мицуюки; Симада, Казуюки (2007). «В верхней губе человека существуют остатки вибриссальных капсульных мышц». Клиническая анатомия . 20 (6): 628–31. дои : 10.1002/ca.20497 . ПМИД 17458869 . S2CID 21055062 .
- ^ Аверси-Феррейра, Роклин AGMF; Бретас, Рафаэль Виейра; Майор, Рафаэль Соуто; Даваасурен, Мункхзул; Парагуассу-Чавес, Карлос Альберто; Нисидзё, Хисао; Аверси-Феррейра, Сказки Александра (2014). «Морфометрический и статистический анализ длинной ладонной мышцы человека и нечеловекообразных приматов» . БиоМед Исследования Интернэшнл . 2014 : 1–6. дои : 10.1155/2014/178906 . ПМК 4016873 . ПМИД 24860810 .
- ^ Капур, Судхир К.; Тивари, Акшай; Кумар, Абхишек; Бхатия, Раджеш; Тантувей, Винай; Капур, Саураб (2008). «Клиническая значимость агенезии длинной ладонной мышцы: распространенная анатомическая аберрация». Международная анатомическая наука . 83 (1): 45–8. дои : 10.1111/j.1447-073X.2007.00199.x . ПМИД 18402087 . S2CID 25324691 .
- ^ Себастин, С; Лим, А; Би, Вт; Вонг, Т; Метил, Б. (2005). «Влияет ли отсутствие длинной ладонной мышцы на силу хвата и сжатия?». Журнал хирургии руки: Журнал Британского общества хирургии руки . 30 (4): 406–8. дои : 10.1016/j.jhsb.2005.03.011 . ПМИД 15935531 . S2CID 35394120 .
- ^ Рубинштейн, Дэвид; Эскотт, Эдвард Дж.; Хендрик, Лаура Л. (апрель 1999 г.). «Распространенность и КТ-изображение мышцы, поднимающей ключицу: нормальный вариант, который не следует путать с аномалией» (PDF) . AJNR Am J Нейрорадиол . 20 (4): 583–6. ПМК 7056035 . ПМИД 10319965 .
- ^ Лукас, М.; Салливан, А.; Таббс, RS; Шоджа, ММ (2008). «Levator claviculae: описание случая и обзор литературы» . Фолия Морфол . 67 (4): 307–310. ПМИД 19085875 .
- ^ Лавинг, Ричард М.; Андерсон, Ларри Д. (2008). «Архитектура и тип волокон пирамидной мышцы» . Международная анатомическая наука . 83 (4): 294–7. дои : 10.1111/j.1447-073X.2007.00226.x . ПМЦ 3531545 . ПМИД 19159363 .
- ^ П., Ханинец; Р., Томаш; Р., Кайзер; Р., Чихак (2009). «Развитие и клиническое значение спинно-эпитрохлеарной мышцы у мужчин». Клиническая анатомия . 22 (4): 481–8. дои : 10.1002/ca.20799 . ПМИД 19373904 . S2CID 221547558 .
- ^ Эдвардс, Уильям Э., Скелетно-мышечная анатомия грудной клетки и плеча взрослой самки шимпанзе , 6571-я лаборатория аэромедицинских исследований, Нью-Мексико, 1965. http://www.dtic.mil/dtic/tr/fulltext/u2/462433 .pdf Архивировано 11 февраля 2017 г. в Wayback Machine.
- ^ «Анатомические атласы: Иллюстрированная энциклопедия анатомических изменений человека: Опус I: Мышечная система: Алфавитный список мышц: L: Широчайшая мышца спины» .
- ^ Огата, Сигэмицу; Мой, Казухару; Тамацу, Юичи; Симада, Казуюки (2002). «Морфологическое исследование хондроглоссальной мышцы человека на японском языке». Анналы анатомии — Anatomischer Anzeiger . 184 (5): 493–9. дои : 10.1016/S0940-9602(02)80087-5 . ПМИД 12392330 .
- ^ Каява, Ю (1915). «Пропорции лишних сосков среди населения Финляндии». Дуодецим . 1 : 143–70.
- ^ Гоял, Таранг; Бакши, СК; Варшней, Анупам (2012). «Семь сосков у мужчины: второй случай в мире» . Индийский журнал генетики человека . 18 (3): 373–5. doi : 10.4103/0971-6866.108051 (неактивен 9 марта 2024 г.). ПМЦ 3656534 . ПМИД 23716953 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на март 2024 г. ( ссылка ) - ^ Теплица, Дэвид; Кович, Грант; Срок, Джейми; Уитакер, Роберт; Джефферс, Эйлин; Вагстафф, Дэвид А. (14 октября 2021 г.). «Недавно выявленная общая анатомия человека: восемь парных рудиментарных грудных бугорков проходят вдоль эмбриологических гребней молочной железы у худых взрослых» . Пластическая и реконструктивная хирургия – Global Open . 9 (10): е3863. дои : 10.1097/GOX.0000000000003863 . ISSN 2169-7574 . ПМЦ 8517303 . ПМИД 34667697 .
- ^ Jump up to: Перейти обратно: а б с д Лаура Спинни (2008), «Остатки эволюции», New Scientist , 198 (2656): 42–45, doi : 10.1016/S0262-4079(08)61231-2
- ^ Дарвин, Чарльз. (1872) Выражение эмоций у человека и животных Джон Мюррей, Лондон. [ нужна страница ]
- ^ Питер Грей (2007). Психология (пятое изд.). Стоит издательства. п. 66 . ISBN 978-0-7167-0617-5 .
- ^ «Развитие поведения у младенцев » (через Google Книги), автор Эвелин Дьюи, со ссылкой на исследование «Рефлексы и другая двигательная активность у новорожденных: отчет о 125 случаях как предварительное исследование поведения младенцев», опубликованное в Bull. Нейрол. Инст. Нью-Йорк , 1932, Том. 2, стр. 1–56.
- ^ Джерри Койн (2009). Почему эволюция верна . Группа Пингвин. стр. 85–86 . ISBN 978-0-670-02053-9 .
- ^ Энтони Стивенс (1982). Архетип: естественная история личности . Рутледж и Кеган Пол. п. 87. ИСБН 978-0-7100-0980-7 .
- ^ Штраус, К.; Василакос, К.; Уилсон, RJA; Осима, Т.; Цельтер, М.; Деренн, доктор философии; Симиловский, Т.; Уайтлоу, Вашингтон (2003). «Филогенетическая гипотеза происхождения икоты». Биоэссе . 25 (2): 182–8. дои : 10.1002/bies.10224 . ПМИД 12539245 .
- ^ Охта, Юрико; Нисикими, Моримицу (1999). «Случайные нуклеотидные замены в нефункциональном гене приматов l-гулоно-γ-лактоноксидазы, недостающего фермента биосинтеза l-аскорбиновой кислоты». Biochimica et Biophysical Acta (BBA) – Общие предметы . 1472 (1–2): 408–11. дои : 10.1016/S0304-4165(99)00123-3 . ПМИД 10572964 .
- ^ Нисикими М., Фукуяма Р., Миношима С., Симидзу Н., Яги К. (6 мая 1994 г.). «Клонирование и хромосомное картирование человеческого нефункционального гена L-гулоно-гамма-лактоноксидазы, фермента биосинтеза L-аскорбиновой кислоты, отсутствующего у человека» . Ж. Биол. Хим . 269 (18): 13685–8. дои : 10.1016/S0021-9258(17)36884-9 . ПМИД 8175804 .
Дальнейшее чтение [ править ]
- Шубин, Нил (2009). Ваша внутренняя рыба: путешествие в 3,5-миллиардную историю человеческого тела . Нью-Йорк: Винтажные книги. ISBN 978-0-307-27745-9 .