Естественный отбор


| Часть серии о |
| Эволюционная биология |
|---|
 |
Естественный отбор — это дифференциальное выживание и воспроизводство особей вследствие различий в фенотипе . Это ключевой механизм эволюции , изменение наследственных признаков, характерных для популяции на протяжении поколений. Чарльз Дарвин популяризировал термин «естественный отбор», противопоставив его искусственному отбору , который является преднамеренным, а естественный отбор — нет.
Изменчивость признаков, как генотипических , так и фенотипических , существует во всех популяциях организмов . Однако некоторые черты с большей вероятностью способствуют выживанию и репродуктивному успеху . Таким образом, эти черты передаются следующему поколению. Эти черты также могут стать более распространенными в популяции , если среда, благоприятствующая этим чертам, останется неизменной. Если новые черты становятся более предпочтительными из-за изменений в определенной нише , микроэволюция происходит . Если новые черты становятся более предпочтительными из-за изменений в более широкой среде, макроэволюция происходит . Иногда могут возникнуть новые виды , особенно если эти новые черты радикально отличаются от черт, которыми обладали их предшественники.
Вероятность того, что эти черты будут «выбраны» и переданы по наследству, определяется многими факторами. Некоторые из них, вероятно, будут переданы по наследству, поскольку они хорошо адаптируются к окружающей среде. Другие передаются по наследству, потому что партнеры по спариванию активно предпочитают эти черты, что известно как половой отбор . Женские тела также предпочитают черты, которые приносят наименьшие затраты на их репродуктивное здоровье, что известно как отбор плодовитости .
Естественный отбор является краеугольным камнем современной биологии . Концепция, опубликованная Дарвином и Альфредом Расселом Уоллесом в совместной презентации статей в 1858 году , была развита во влиятельной книге Дарвина 1859 года «Происхождение видов посредством естественного отбора, или сохранение избранных рас в борьбе за жизнь» . Он описал естественный отбор как аналог искусственного отбора, процесса, посредством которого животные и растения с признаками, которые селекционеры считают желательными, систематически отдают предпочтение для воспроизводства. Концепция естественного отбора первоначально возникла в отсутствие действующей теории наследственности; на момент написания Дарвина наука еще не разработала современные теории генетики. Объединение традиционной дарвиновской эволюции с последующими открытиями классической генетики сформировало современный синтез середины 20 века. Появление молекулярной генетики привело к появлению эволюционной биологии развития , которая объясняет эволюцию на молекулярном уровне. Пока генотипы могут медленно меняться в результате случайного генетического дрейфа , естественный отбор остается основным объяснением адаптивной эволюции .
Историческое развитие
Додарвиновские теории

Несколько философов классической эпохи , в том числе Эмпедокл [1] и его интеллектуальный преемник, римский поэт Лукреций , [2] высказал идею о том, что природа производит огромное разнообразие существ случайным образом и что выживают только те существа, которым удается обеспечить себя и успешно размножаться. Идея Эмпедокла о том, что организмы возникли исключительно в результате случайного действия таких причин, как тепло и холод, подверглась критике со стороны Аристотеля во второй книге физики . [3] Он поставил на место естественную телеологию и считал, что форма была достигнута с определенной целью, ссылаясь на регулярность наследственности у видов в качестве доказательства. [4] [5] Тем не менее он принял в своей биологии , что новые виды животных, чудовища (τερας), могут встречаться в очень редких случаях ( «Поколение животных» , Книга IV). [6] 1872 года Как цитируется в издании Дарвина «Происхождение видов» , Аристотель рассматривал вопрос о том, могли ли разные формы (например, зубы) появиться случайно, но выжили только полезные формы:
Так что же мешает различным частям [тела] иметь это просто случайное соотношение в природе? так как зубы, например, растут по необходимости, передние острые, приспособленные для деления, а измельчители плоские и пригодные для пережевывания пищи; так как они были сделаны не ради этого, а это было результатом случайности. То же самое касается и других частей, в которых, по-видимому, существует приспособление к цели. Итак, где бы все вещи вместе (т. е. все части одного целого) ни происходили так, как если бы они были созданы ради чего-то, они сохранялись, будучи соответствующим образом составлены внутренней спонтанностью, а все, что вещи не были составлены таким образом, , погиб и до сих пор погибает.
- Аристотель, Физика , Книга II, Глава 8. [7]
Но Аристотель отверг эту возможность в следующем абзаце, уточнив, что он говорит о развитии животных как эмбрионов с фразой «либо неизменно, либо нормально возникает», а не о происхождении видов:
... Однако невозможно, чтобы это было истинное мнение. Ведь зубы и все другие естественные вещи либо неизменно, либо обычно возникают определенным образом; но это не относится ни к одному из результатов случайности или спонтанности. Частоту дождя зимой мы не приписываем случайности или простому совпадению, но мы приписываем частые дожди летом; ни тепла в собачьи дни, но только если оно у нас есть зимой. Итак, если принято, что вещи существуют либо в результате совпадения, либо ради цели, а они не могут быть результатом совпадения или спонтанности, то отсюда следует, что они должны существовать ради цели; и что все эти вещи обусловлены природой, с этим согласятся даже сторонники теории, которая находится перед нами. Следовательно, действие ради цели присутствует в вещах, которые возникают и существуют по своей природе.
- Аристотель, Физика , Книга II, Глава 8. [8]
Борьба за существование была позже описана исламским писателем Аль-Джахизом в 9 веке, особенно в контексте нисходящего регулирования численности населения, но не в отношении индивидуальных вариаций или естественного отбора. [9] [10]
На рубеже 16-го века Леонардо да Винчи собрал множество окаменелостей аммонитов, а также другой биологический материал. В своих трудах он подробно рассуждал, что формы животных не даны раз и навсегда «высшими силами», а порождаются в различных формах естественным путем и затем отбираются для воспроизводства по их совместимости с окружающей средой. [11]
Более поздние классические аргументы были вновь представлены в 18 веке Пьером Луи Мопертюи. [12] и другие, в том числе дедушка Дарвина, Эразм Дарвин .
До начала XIX века преобладала точка зрения в западных обществах , согласно которой различия между особями одного вида были неинтересными отклонениями от их платонических идеалов (или типов ) созданных видов . Однако теория униформизма в геологии продвигала идею о том, что простые, слабые силы могут действовать непрерывно в течение длительных периодов времени, вызывая радикальные изменения в ландшафте Земли. Успех этой теории повысил осознание огромных масштабов геологического времени и сделал правдоподобной идею о том, что крошечные, практически незаметные изменения в последующих поколениях могут привести к последствиям масштаба различий между видами. [13]
Зоолог начала XIX века Жан-Батист Ламарк предположил, что наследование приобретенных характеристик является механизмом эволюционных изменений; адаптивные черты, приобретенные организмом в течение его жизни, могут быть унаследованы потомством этого организма, что в конечном итоге приводит к трансмутации видов . [14] Эта теория, ламаркизм , оказала влияние на советского биолога Трофима Лысенко с господствующей генетической теорией еще в середине 20-го века. злополучный антагонизм [15]
Между 1835 и 1837 годами зоолог Эдвард Блит работал над областью изменчивости, искусственного отбора и над тем, как подобный процесс происходит в природе. Дарвин признал идеи Блита в первой главе о вариациях « Происхождения видов» . [16]
Теория Дарвина
В 1859 году Чарльз Дарвин изложил свою теорию эволюции путем естественного отбора как объяснение адаптации и видообразования. Он определил естественный отбор как «принцип, согласно которому сохраняется каждое незначительное изменение [признака], если оно полезно». [17] Идея была простой, но мощной: особи, лучше всего приспособленные к окружающей среде, с большей вероятностью выживут и размножатся. Пока между ними существуют некоторые различия, и эти различия передаются по наследству , будет неизбежен отбор особей с наиболее выгодными вариациями. Если вариации наследуются, то дифференциальный репродуктивный успех приводит к эволюции определенных популяций вида, и популяции, которые эволюционируют и становятся достаточно разными, в конечном итоге становятся разными видами. [18] [19]

Идеи Дарвина были вдохновлены наблюдениями, которые он сделал во время второго рейса HMS Beagle (1831–1836), а также работой политического экономиста Томаса Роберта Мальтуса , который в «Очерке о принципе народонаселения» (1798) , отметил, что население (если его не контролировать) увеличивается в геометрической прогрессии , тогда как запасы продовольствия растут только арифметически ; таким образом, неизбежные ограничения ресурсов будут иметь демографические последствия, ведущие к «борьбе за существование». [20] уже позволила ему Когда Дарвин прочитал Мальтуса в 1838 году, его работа как натуралиста оценить «борьбу за существование» в природе. Его поразило, что по мере того, как популяция перерастет ресурсы, «благоприятные вариации будут иметь тенденцию сохраняться, а неблагоприятные — уничтожаться. Результатом этого будет образование новых видов». [21] Дарвин писал:
Если в течение длительного периода времени и при различных условиях жизни органические существа вообще различаются в различных частях своей организации, и я думаю, что это не может быть оспорено; если из-за высоких геометрических способностей размножения каждого вида в каком-то возрасте, сезоне или году происходит жестокая борьба за жизнь, и это, конечно, не может быть оспорено; тогда, принимая во внимание бесконечную сложность отношений всех органических существ друг к другу и к условиям их существования, приводящую к тому, что бесконечное разнообразие в строении, конституции и привычках оказывается для них выгодным, я думаю, что это был бы самый необычайный факт если бы никогда не происходило ни одного изменения, полезного для собственного благополучия каждого существа, точно так же, как столько же изменений произошло бы, полезного для человека. Но если действительно произойдут вариации, полезные для какого-либо органического существа, то, несомненно, особи, характеризующиеся таким образом, будут иметь наилучшие шансы сохраниться в борьбе за жизнь; и в силу сильного принципа наследования они будут иметь тенденцию производить потомство с аналогичными характеристиками. Этот принцип сохранения я для краткости назвал естественным отбором.
- Дарвин резюмирует естественный отбор в четвертой главе книги «Происхождение видов». [22]
Получив свою теорию, Дарвин тщательно собирал и уточнял доказательства, прежде чем обнародовать свою идею. Он был в процессе написания своей «большой книги», чтобы представить свои исследования, когда натуралист Альфред Рассел Уоллес независимо придумал этот принцип и описал его в эссе, которое он отправил Дарвину для передачи Чарльзу Лайелю . Лайель и Джозеф Далтон Хукер решили представить свое эссе вместе с неопубликованными работами, которые Дарвин разослал коллегам-натуралистам, и « О склонности видов к образованию разновидностей»; и «Увековечение разновидностей и видов естественными средствами селекции» были зачитаны Лондонскому Линнеевскому обществу, в котором было объявлено о совместном открытии этого принципа в июле 1858 года. [23] Дарвин опубликовал подробный отчет о своих доказательствах и выводах в книге «Происхождение видов» в 1859 году. В третьем издании 1861 года Дарвин признал, что другие, такие как Уильям Чарльз Уэллс в 1813 году и Патрик Мэтью в 1831 году, предлагали аналогичные идеи, но не разработал их и не представил в заметных научных публикациях. [24]

Дарвин думал о естественном отборе по аналогии с тем, как фермеры отбирают сельскохозяйственные культуры или домашний скот для разведения, что он назвал « искусственным отбором »; в своих ранних рукописях он ссылался на «Природу», которая будет осуществлять отбор. В то время другие механизмы эволюции, такие как эволюция путем генетического дрейфа, еще не были четко сформулированы, и Дарвин полагал, что отбор, вероятно, был лишь частью истории: «Я убежден, что естественный отбор был главным, но не исключительным средством модификации. ." [25] В письме Чарльзу Лайелю в сентябре 1860 года Дарвин выразил сожаление по поводу использования термина «естественный отбор», отдав предпочтение термину «естественное сохранение». [26]
Для Дарвина и его современников естественный отбор был, по сути, синонимом эволюции путем естественного отбора. После публикации « Происхождения видов » [27] образованные люди обычно признавали, что эволюция произошла в той или иной форме. Однако естественный отбор как механизм оставался спорным, отчасти потому, что он считался слишком слабым для объяснения диапазона наблюдаемых характеристик живых организмов, а отчасти потому, что даже сторонники эволюции возражали против его «неуправляемой» и непрогрессивной природы . [28] ответ, который был охарактеризован как единственное и наиболее существенное препятствие на пути принятия идеи. [29] Однако некоторые мыслители с энтузиазмом поддержали естественный отбор; Прочитав Дарвина, Герберт Спенсер ввел фразу «выживает сильнейший» , которая стала популярным изложением теории. [30] [31] В пятое издание « О происхождении видов» , опубликованное в 1869 году, была включена фраза Спенсера как альтернатива естественному отбору, с указанием авторства: «Но выражение, часто используемое г-ном Гербертом Спенсером в книге «Выживание наиболее приспособленных», является более точным, а иногда и более точным. одинаково удобно». [32] Хотя эта фраза до сих пор часто используется небиологами, современные биологи избегают ее, поскольку она является тавтологией, если «наиболее приспособленный» читается как «функционально превосходящий» и применяется к отдельным людям, а не рассматривается как усредненная величина по популяциям. [33]
Современный синтез
Естественный отбор в решающей степени опирается на идею наследственности, но он был разработан раньше основных концепций генетики . Хотя моравский монах Грегор Мендель , отец современной генетики, был современником Дарвина, его работы оставались в безвестности и были вновь открыты только в 1900 году. [34] С интеграцией эволюции в начале 20-го века с законами наследственности Менделя , так называемым современным синтезом , ученые в целом пришли к признанию естественного отбора. [35] [36] Синтез вырос из достижений в различных областях. Рональд Фишер разработал необходимый математический язык и написал «Генетическая теория естественного отбора» (1930). [37] Дж. Б. С. Холдейн представил концепцию «цены» естественного отбора. [38] [39] Сьюэлл Райт объяснил природу отбора и адаптации. [40] В своей книге «Генетика и происхождение видов » (1937) Феодосий Добжанский установил идею о том, что мутация, которую когда-то считали соперником отбора, на самом деле поставляет сырье для естественного отбора, создавая генетическое разнообразие. [41] [42]
Второй синтез

Эрнст Майр признал ключевую важность репродуктивной изоляции для видообразования в своей «Систематике и происхождении видов» (1942). [44] У. Д. Гамильтон придумал родственный отбор в 1964 году. [45] [46] Этот синтез закрепил естественный отбор в качестве основы эволюционной теории, где он остается и сегодня. Второй синтез был осуществлен в конце 20-го века благодаря достижениям молекулярной генетики , создавшим область эволюционной биологии развития («эво-дево»), которая пытается объяснить эволюцию формы с точки зрения генетических регуляторных программ , которые контролировать развитие эмбриона на молекулярном уровне. Здесь понимается, что естественный отбор воздействует на эмбриональное развитие, изменяя морфологию взрослого тела. [47] [48] [49] [50]
Терминология
Термин «естественный отбор» чаще всего применяется к наследственным признакам, поскольку они непосредственно участвуют в эволюции. Однако естественный отбор «слеп» в том смысле, что изменения фенотипа могут дать репродуктивное преимущество независимо от того, передается ли этот признак по наследству. Следуя основному использованию Дарвина, этот термин используется для обозначения как эволюционных последствий слепого отбора, так и его механизмов. [27] [37] [51] [52] Иногда полезно четко различать механизмы отбора и его эффекты; когда это различие важно, ученые определяют «(фенотипический) естественный отбор» именно как «те механизмы, которые способствуют отбору воспроизводящихся особей», независимо от того, является ли основа отбора наследственной. [53] [54] [55] Говорят, что признаки, которые обеспечивают больший репродуктивный успех организма, отбираются , а те, которые снижают успех, отбираются против . [56]
Механизм
Наследственная изменчивость, дифференциальное воспроизводство

Естественная изменчивость возникает среди особей любой популяции организмов. Некоторые различия могут повысить шансы человека на выживание и размножение, так что его репродуктивная способность в течение всей жизни увеличивается, а это означает, что он оставляет больше потомства. Если признаки, дающие этим особям репродуктивное преимущество, также наследуются , то есть передаются от родителя к потомству, то будет дифференцированное размножение, то есть несколько более высокая доля быстрых кроликов или эффективных водорослей в следующем поколении. Даже если репродуктивное преимущество очень незначительное, в течение многих поколений любой выгодный наследственный признак становится доминирующим в популяции. Таким образом, естественная среда организма «отбирает» черты, которые дают репродуктивное преимущество, вызывая эволюционные изменения, как описал Дарвин. [57] Это создает видимость цели, но в естественном отборе нет намеренного выбора. [а] Искусственный отбор является целенаправленным , в отличие от естественного отбора, хотя биологи часто используют телеологический язык для его описания. [58]
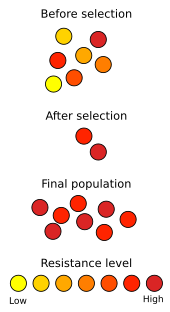
Перцовая моль существует в Великобритании как светлого, так и темного цвета, но во время промышленной революции многие деревья, на которых отдыхала моль, почернели от сажи , что дало темным бабочкам преимущество в укрытии от хищников. Это дало темным бабочкам больше шансов выжить и произвести потомство темного цвета, и всего за пятьдесят лет с момента поимки первой темной бабочки почти все бабочки в промышленном Манчестере были темными. Баланс был нарушен в результате принятия Закона о чистом воздухе 1956 года , и темные бабочки снова стали редкими, демонстрируя влияние естественного отбора на эволюцию перцовой моли . [59] Недавнее исследование с использованием анализа изображений и моделей птичьего зрения показало, что бледные особи более точно соответствуют фону лишайников, чем темные морфы, и впервые количественно оценили маскировку бабочек от риска нападения хищников . [60]
Фитнес
Концепция приспособленности занимает центральное место в естественном отборе. В широком смысле, люди, которые более «приспособлены», имеют лучший потенциал для выживания, как в известной фразе « выживает наиболее приспособленный », но точное значение этого термина гораздо более тонкое. Современная эволюционная теория определяет приспособленность не по тому, как долго живет организм, а по тому, насколько успешно он размножается. Если организм живет вдвое меньше, чем другие представители его вида, но имеет в два раза больше потомков, доживающих до взрослого возраста, его гены становятся более распространенными во взрослой популяции следующего поколения. Хотя естественный отбор действует на особей, влияние случайности означает, что приспособленность действительно может быть определена только «в среднем» для особей в популяции. Пригодность определенного генотипа соответствует среднему эффекту для всех людей с этим генотипом. [61] Необходимо проводить различие между понятиями «выживание наиболее приспособленных» и «улучшение приспособленности». «Выживание наиболее приспособленных» не приводит к «улучшению приспособленности», оно лишь представляет собой удаление из популяции менее приспособленных вариантов. Математический пример «выживания наиболее приспособленных» приводит Холдейн в своей статье «Цена естественного отбора». [62] Холдейн назвал этот процесс «замещением» или, что чаще в биологии, это называется «фиксацией». Это правильно описывается дифференциальной выживаемостью и размножением особей из-за различий в фенотипе. С другой стороны, «улучшение приспособленности» не зависит от дифференциальной выживаемости и воспроизводства особей из-за различий в фенотипе, оно зависит от абсолютной выживаемости конкретного варианта. Вероятность возникновения полезной мутации у какого-либо члена популяции зависит от общего количества репликаций этого варианта. Математика «улучшения физической формы» была описана Кляйнманом. [63] Эмпирический пример «улучшения физической формы» дан экспериментом «Мега-пластина Кишони». [64] В этом эксперименте «улучшение приспособленности» зависит от количества повторений конкретного варианта, чтобы появился новый вариант, способный расти в следующей области с более высокой концентрацией лекарства. Для такого «улучшения пригодности» не требуется фиксации или замены. С другой стороны, «улучшение приспособленности» может произойти в среде, где также действует «выживание наиболее приспособленных». эксперимент Ричарда Ленски Классический E. coli по долгосрочной эволюции является примером адаптации в конкурентной среде («улучшение физической формы» во время «выживания наиболее приспособленных»). [65] Вероятность того, что у какого-либо представителя линии произойдет полезная мутация, улучшающая приспособленность, снижается из-за конкуренции. Вариант, который является кандидатом на полезную мутацию в этой среде с ограниченной несущей способностью, должен сначала превзойти «менее подходящие» варианты, чтобы накопить необходимое количество репликаций, чтобы существовала разумная вероятность возникновения этой полезной мутации. [66]
Соревнование
В биологии конкуренция — это взаимодействие между организмами, при котором приспособленность одного снижается из-за присутствия другого. Это может быть связано с тем, что оба полагаются на ограниченные запасы ресурсов, таких как еда, вода или территория . [67] Конкуренция может быть внутри вида или между видами , а также может быть прямой или косвенной. [68] Виды, менее приспособленные к конкуренции, теоретически должны либо адаптироваться, либо вымереть , поскольку конкуренция играет важную роль в естественном отборе, но согласно теории «пространства для передвижения» она может быть менее важной, чем расширение среди более крупных клад . [68] [69]
Конкуренция моделируется r/K теорией отбора , которая основана на Роберта Макартура и Э.О. Уилсона работах по биогеографии островов . [70] В этой теории давление отбора ведет эволюцию в одном из двух стереотипных направлений: r- или K -отбор. [71] Эти термины r и K можно проиллюстрировать в логистической модели динамики населения : [72]

Классификация

2, стабилизирующий отбор : промежуточные значения предпочтительнее крайних.
3: разрушительный отбор: крайние варианты предпочтительнее промежуточных.
Ось X: фенотипический признак
Ось Y: количество организмов
Группа А: исходное население
Группа B: после отбора
Естественный отбор может воздействовать на любой наследуемый фенотипический признак . [73] и селективное давление может быть произведено любым аспектом окружающей среды, включая половой отбор и конкуренцию с представителями того же или другого вида. [74] [75] Однако это не означает, что естественный отбор всегда направлен и приводит к адаптивной эволюции; естественный отбор часто приводит к сохранению статус-кво за счет исключения менее подходящих вариантов. [57]
Отбор можно классифицировать по-разному, например, по его влиянию на признак, на генетическое разнообразие, по стадии жизненного цикла, на которой он действует, по единице отбора или по ресурсу, за который ведется конкуренция.
По влиянию на признак
Отбор по-разному влияет на признаки. Стабилизирующий отбор удерживает признак на стабильном оптимальном уровне, и в простейшем случае все отклонения от этого оптимума избирательно невыгодны. Направленный отбор благоприятствует крайним значениям признака. Необычный разрушительный отбор также действует во время переходных периодов, когда текущий режим неоптимален, но изменяет признак более чем в одном направлении. В частности, если признак является количественным и одномерным, то предпочтение отдается как более высокому, так и более низкому уровню признака. Разрушительный отбор может быть предшественником видообразования . [57]
По влиянию на генетическое разнообразие
Альтернативно, отбор можно разделить в зависимости от его влияния на генетическое разнообразие . Очищающий или негативный отбор направлен на удаление генетических вариаций из популяции (и этому противостоит de novo мутация , которая вводит новые вариации). [76] [77] Напротив, балансирующий отбор способствует поддержанию генетической изменчивости в популяции даже при отсутствии мутаций de novo посредством отрицательного частотно-зависимого отбора . Одним из механизмов этого является преимущество гетерозигот , когда люди с двумя разными аллелями имеют селективное преимущество перед людьми с одним аллелем. полиморфизм локуса группы крови АВО человека. Таким образом был объяснен [78]
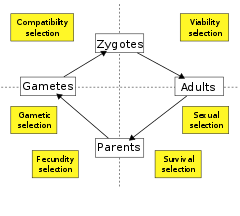
По стадии жизненного цикла
Другой вариант — классифицировать отбор по стадиям жизненного цикла , на которых он действует. Некоторые биологи выделяют всего два типа: отбор на жизнеспособность (или выживание) , который увеличивает вероятность выживания организма, и отбор по плодовитости (или плодовитости, или репродуктивности), который увеличивает скорость воспроизводства при условии выживания. Другие разделяют жизненный цикл на дополнительные компоненты отбора. Таким образом, отбор на жизнеспособность и выживание можно определить отдельно и соответственно как действие, направленное на повышение вероятности выживания до и после достижения репродуктивного возраста, в то время как отбор плодовитости может быть разделен на дополнительные подкомпоненты, включая половой отбор, гаметический отбор, воздействие на гамет выживание , и отбор совместимости, действующий на зигот . образование [79]
По единице выбора
Отбор также можно классифицировать по уровню или единице отбора . Индивидуальный отбор действует на индивидуума в том смысле, что адаптации осуществляются «для» блага индивидуума и являются результатом отбора среди индивидуумов. Генный отбор действует непосредственно на уровне гена. В родственном отборе и внутригеномном конфликте отбор на уровне генов обеспечивает более подходящее объяснение лежащего в основе процесса. Групповой отбор , если он имеет место, действует на группы организмов, исходя из предположения, что группы реплицируются и мутируют аналогично генам и особям. Продолжаются споры о том, в какой степени групповой отбор происходит в природе. [80]
По ресурсу, за который конкурируют

Наконец, отбор можно классифицировать в зависимости от ресурса, за который ведется конкуренция. Половой отбор является результатом конкуренции за партнеров. Половой отбор обычно происходит через отбор по плодовитости, иногда в ущерб жизнеспособности. Экологический отбор — это естественный отбор, осуществляемый любыми способами, кроме полового отбора, такими как родственный отбор, конкуренция и детоубийство . Вслед за Дарвином естественный отбор иногда определяют как экологический отбор, и в этом случае половой отбор считают отдельным механизмом. [83]
Половой отбор, впервые сформулированный Дарвином (на примере павлина ) хвоста [81] относится конкретно к конкуренции за партнеров, [84] который может быть внутрисексуальным , между особями одного пола, то есть соревнованием между мужчинами, или интерсексуальным , когда один пол выбирает партнеров , чаще всего при этом самцы демонстрируют, а самки выбирают. [85] Однако у некоторых видов выбор партнера осуществляется преимущественно самцами, как у некоторых рыб семейства Syngnathidae . [86] [87]
Фенотипические признаки могут проявляться у одного пола и желательны у другого пола, вызывая петлю положительной обратной связи , называемую « побегом Фишера» , например, экстравагантное оперение некоторых самцов птиц, таких как павлин. [82] Альтернативная теория, предложенная тем же Рональдом Фишером в 1930 году, — это гипотеза сексуального сына , согласно которой матери хотят, чтобы беспорядочные сыновья подарили им большое количество внуков, и поэтому выбирают беспорядочных отцов для своих детей. Агрессия между представителями одного пола иногда связана с очень отличительными чертами, такими как рога оленей , которые используются в бою с другими оленями. В более общем смысле, внутриполовой отбор часто связан с половым диморфизмом , включая различия в размерах тела между самцами и самками одного вида. [85]
Гонка вооружений
Естественный отбор проявляется в развитии устойчивости к микроорганизмов антибиотикам . С момента открытия пенициллина в 1928 году антибиотики использовались для борьбы с бактериальными заболеваниями. Широкое злоупотребление антибиотиками привело к развитию устойчивости микробов к антибиотикам при клиническом использовании до такой степени, что устойчивый к метициллину золотистый стафилококк (MRSA) был описан как «супербактерия» из-за угрозы, которую он представляет для здоровья, и его относительной неуязвимости к вирусам. существующие препараты. [88] Стратегии реагирования обычно включают использование других, более сильных антибиотиков; однако недавно появились новые штаммы MRSA, устойчивые даже к этим препаратам. [89] Это эволюционная гонка вооружений , в которой бактерии создают штаммы, менее восприимчивые к антибиотикам, в то время как исследователи-медики пытаются разработать новые антибиотики, способные их убить. Аналогичная ситуация происходит с устойчивостью растений и насекомых к пестицидам. Гонка вооружений не обязательно вызвана человеком; хорошо задокументированный пример включает распространение гена у бабочки Hypolimnas bolina, убивающих самцов, подавляющего активность бактерий-паразитов Wolbachia, на острове Самоа , где, как известно, распространение гена произошло в течение всего пяти лет. [90] [91]
Эволюция посредством естественного отбора
Предпосылкой для того, чтобы естественный отбор привел к адаптивной эволюции, новым признакам и видообразованию, является наличие наследуемых генетических вариаций, которые приводят к различиям в приспособленности. Генетическая изменчивость является результатом мутаций, генетических рекомбинаций и изменений кариотипа ( числа, формы, размера и внутреннего расположения хромосом ) . Любое из этих изменений может иметь как очень выгодный, так и крайне невыгодный эффект, но значительные последствия редки. В прошлом большинство изменений в генетическом материале считались нейтральными или близкими к нейтральным, поскольку они происходили в некодирующей ДНК или приводили к синонимичным заменам . Однако многие мутации в некодирующей ДНК имеют пагубные последствия. [92] [93] Хотя как частота мутаций, так и средний эффект мутаций на приспособленность зависят от организма, большинство мутаций у людей слегка вредны. [94]
Некоторые мутации происходят в «наборе инструментов» или регуляторных генах . Изменения в них часто оказывают большое влияние на фенотип человека, поскольку они регулируют функцию многих других генов. Большинство, но не все, мутации в регуляторных генах приводят к нежизнеспособности эмбрионов. у человека происходят некоторые несмертельные регуляторные мутации В генах HOX , которые могут привести к повреждению шейного ребра. [95] или полидактилия , увеличение количества пальцев рук или ног. [96] Когда такие мутации приводят к более высокой приспособленности, естественный отбор благоприятствует этим фенотипам, и новый признак распространяется в популяции.Установленные черты не являются неизменными; черты, которые имеют высокую приспособленность в одном контексте окружающей среды, могут оказаться гораздо менее подходящими, если условия окружающей среды изменятся. В отсутствие естественного отбора для сохранения такого признака он становится более изменчивым и со временем ухудшается, что, возможно, приводит к рудиментарному проявлению признака, также называемому эволюционным багажом . Во многих обстоятельствах очевидно рудиментарная структура может сохранять ограниченную функциональность или может быть использована для получения других полезных свойств в результате явления, известного как преадаптация . Считается , что известный пример рудиментарной структуры — глаз слепого землекопа — сохраняет функцию восприятия фотопериода . [97]
Видообразование
Видообразование требует определенной степени репродуктивной изоляции , то есть уменьшения потока генов. Однако это присуще концепции вида , против которого отбираются гибриды , что противоречит эволюции репродуктивной изоляции - проблеме, которая была признана Дарвином. Проблема не возникает при аллопатрическом видообразовании с географически разделенными популяциями, которые могут расходиться с разными наборами мутаций. Э.Б. Поултон понял в 1903 году, что репродуктивная изоляция может развиваться посредством дивергенции, если каждая линия приобретет разные, несовместимые аллели одного и того же гена. Отбор против гетерозиготы тогда непосредственно создал бы репродуктивную изоляцию, что привело бы к модели Бейтсона-Добжанского-Мюллера , далее развитой Х. Алленом Орром. [98] and Sergey Gavrilets . [99] Однако при подкреплении естественный отбор может способствовать увеличению презиготической изоляции, напрямую влияя на процесс видообразования. [100]
Генетическая основа
Генотип и фенотип
Естественный отбор действует на фенотип или физические характеристики организма. Фенотип определяется генетической структурой организма (генотипом) и средой, в которой живет организм. Когда разные организмы в популяции обладают разными версиями гена определенного признака, каждая из этих версий известна как аллель . Именно эта генетическая изменчивость лежит в основе различий в фенотипе. Примером являются АВО группы крови антигены у человека, фенотип которых управляют тремя аллелями. [101]
Некоторые признаки управляются только одним геном, но на большинство признаков влияет взаимодействие многих генов. Вариация одного из многих генов, влияющих на признак, может иметь лишь незначительное влияние на фенотип; вместе эти гены могут производить континуум возможных фенотипических значений. [102]
Направленность выбора
Когда какой-то компонент признака передается по наследству, отбор изменяет частоты различных аллелей или вариантов гена, который производит варианты признака. Отбор можно разделить на три класса в зависимости от его влияния на частоты аллелей: направленный , стабилизирующий и разрушительный отбор . [103] Направленный отбор происходит, когда аллель имеет большую приспособленность, чем другие, поэтому ее частота увеличивается, занимая все большую долю в популяции. Этот процесс может продолжаться до тех пор, пока аллель не зафиксируется и вся популяция не будет иметь более подходящий фенотип. [104] Гораздо более распространенным является стабилизирующий отбор, который снижает частоту аллелей, оказывающих вредное воздействие на фенотип, то есть производящих организмы с более низкой приспособленностью. Этот процесс может продолжаться до тех пор, пока аллель не будет удален из популяции. Стабилизирующий отбор сохраняет функциональные генетические особенности, такие как гены, кодирующие белки , или регуляторные последовательности , с течением времени за счет селективного давления против вредных вариантов. [105] Подрывной (или диверсифицирующий) отбор — это отбор, отдающий предпочтение крайним значениям признаков над промежуточными значениями признаков. Разрушительный отбор может вызвать симпатрическое видообразование посредством разделения ниш .
Некоторые формы балансирующего отбора не приводят к фиксации, но поддерживают аллель на промежуточных частотах в популяции. Это может произойти у диплоидных видов (с парами хромосом), когда гетерозиготные особи (всего с одной копией аллеля) имеют более высокую приспособленность, чем гомозиготные особи (с двумя копиями). Это называется преимуществом гетерозигот или сверхдоминированием, наиболее известным примером которого является устойчивость к малярии у людей, гетерозиготных по серповидноклеточной анемии . Поддержание аллельной изменчивости также может происходить посредством разрушительного или диверсифицирующего отбора , который благоприятствует генотипам, отклоняющимся от среднего в любом направлении (то есть в противоположность сверхдоминированию), и может привести к бимодальному распределению значений признаков. Наконец, балансирующий отбор может происходить посредством частотно-зависимого отбора, когда приспособленность одного конкретного фенотипа зависит от распределения других фенотипов в популяции. Принципы теории игр применялись для понимания распределения приспособленности в таких ситуациях, особенно при изучении родственного отбора и эволюции взаимный альтруизм . [106] [107]
Отбор, генетическая изменчивость и дрейф
Часть всех генетических вариаций функционально нейтральна, не оказывает фенотипического эффекта или существенных различий в приспособленности. Мотоо Кимуры предполагает Нейтральная теория молекулярной эволюции за счет генетического дрейфа , что эта вариация составляет значительную часть наблюдаемого генетического разнообразия. [108] Нейтральные события могут радикально сократить генетическую изменчивость за счет узких мест в популяциях . [109] что, среди прочего, может вызвать эффект основателя в изначально небольших новых популяциях. [110] Когда генетическая изменчивость не приводит к различиям в приспособленности, отбор не может напрямую влиять на частоту таких вариаций. В результате генетическая изменчивость на этих участках выше, чем на участках, где вариации действительно влияют на приспособленность. [103] Однако после периода отсутствия новых мутаций генетическая вариация в этих сайтах устраняется из-за генетического дрейфа. Естественный отбор уменьшает генетическую изменчивость, устраняя дезадаптированных особей и, следовательно, мутации, вызвавшие дезадаптацию. В то же время возникают новые мутации, приводящие к балансу мутаций и отбора . Точный результат этих двух процессов зависит как от скорости возникновения новых мутаций, так и от силы естественного отбора, который зависит от того, насколько неблагоприятной окажется мутация. [111]
Генетическая связь возникает, когда локусы двух аллелей расположены близко на хромосоме. Во время образования гамет рекомбинация меняет аллели. Вероятность того, что такая перестановка произойдет между двумя аллелями, обратно пропорциональна расстоянию между ними. Селективные зачистки происходят, когда аллель становится более распространенной в популяции в результате положительного отбора. По мере увеличения распространенности одного аллеля, тесно связанные аллели также могут стать более распространенными в результате « генетического автостопа », независимо от того, являются ли они нейтральными или даже слегка вредными. Сильная селективная проверка приводит к образованию области генома, где положительно выбранный гаплотип (аллель и его соседи), по сути, являются единственными, существующими в популяции. Выборочные зачистки можно обнаружить путем измерения неравновесия по сцеплению или определения того, чрезмерно ли представлен данный гаплотип в популяции. Поскольку селективная проверка также приводит к отбору соседних аллелей, наличие блока с сильным неравновесием по сцеплению может указывать на «недавнюю» селективную проверку вблизи центра блока. [112]
Выбор фона является противоположностью выборочной развертки. Если конкретный сайт подвергается сильному и постоянному очищающему отбору, связанные вариации имеют тенденцию отсеиваться вместе с ним, создавая область генома с низкой общей изменчивостью. Поскольку фоновый отбор является результатом новых вредных мутаций, которые могут произойти случайно в любом гаплотипе, он не приводит к четким блокам неравновесия по сцеплению, хотя при низкой рекомбинации он все же может привести к слегка отрицательному неравновесию по сцеплению в целом. [113]
Влияние
Идеи Дарвина, наряду с идеями Адама Смита и Карла Маркса , оказали глубокое влияние на мысль XIX века, включая его радикальное утверждение о том, что «тщательно сконструированные формы, столь отличающиеся друг от друга и столь сложным образом зависящие друг от друга», эволюционировали. от простейших форм жизни с помощью нескольких простых принципов. [114] Это вдохновило некоторых из самых ярых сторонников Дарвина и вызвало сильнейшую оппозицию. , естественный отбор имел силу По словам Стивена Джея Гулда «свергнуть с престола некоторые из самых глубоких и традиционных утешений западной мысли», таких как вера в то, что люди занимают особое место в мире. [115]
По словам философа Дэниела Деннета , «опасная идея Дарвина» об эволюции посредством естественного отбора — это «универсальная кислота», которую нельзя удержать в каком-либо сосуде или контейнере, поскольку она вскоре вытекает наружу, пробиваясь во все более обширные пространства. окружение. [116] Таким образом, в последние десятилетия концепция естественного отбора распространилась из эволюционной биологии в другие дисциплины, включая эволюционные вычисления , квантовый дарвинизм , эволюционную экономику , эволюционную эпистемологию , эволюционную психологию и космологический естественный отбор . Эта неограниченная применимость была названа универсальным дарвинизмом . [117]
Происхождение жизни
Как зародилась жизнь из неорганической материи, остается нерешенной проблемой биологии. Одна из известных гипотез заключается в том, что жизнь впервые возникла в форме коротких самовоспроизводящихся полимеров РНК. [118] С этой точки зрения жизнь могла возникнуть, когда цепи РНК впервые испытали основные условия, как это было задумано Чарльзом Дарвином, для действия естественного отбора. Этими условиями являются: наследственность, изменчивость типа и конкуренция за ограниченные ресурсы. Приспособленность раннего репликатора РНК , вероятно, была функцией внутренних адаптивных способностей (т. е. определяемых нуклеотидной последовательностью ) и доступности ресурсов. [119] [120] Логически тремя основными адаптивными способностями могли быть: (1) способность воспроизводиться с умеренной точностью (приводящая как к наследственности, так и к изменчивости типа), (2) способность избегать распада и (3) способность приобретать и ресурсы процесса. [119] [120] Эти способности изначально должны были определяться свернутыми конфигурациями (в том числе конфигурациями с рибозимной активностью) репликаторов РНК, которые, в свою очередь, должны были кодироваться в их индивидуальных нуклеотидных последовательностях. [121]
Клеточная и молекулярная биология
В 1881 году эмбриолог Вильгельм Ру опубликовал Der Kampf der Theile im Organismus ( «Борьба частей в организме »), в которой предположил, что развитие организма является результатом дарвиновской конкуренции между частями эмбриона, происходящей на всех уровнях. от молекул к органам. [122] В последние годы современную версию этой теории предложил Жан-Жак Купец . Согласно этому клеточному дарвинизму, случайные вариации на молекулярном уровне порождают разнообразие типов клеток, тогда как клеточные взаимодействия навязывают характерный порядок развивающемуся эмбриону. [123]
Социально-психологическая теория
Социальные последствия теории эволюции путем естественного отбора также стали источником продолжающихся споров. Фридрих Энгельс , немецкий политический философ и один из создателей идеологии коммунизма , писал в 1872 году, что «Дарвин не знал, какую горькую сатиру он написал на человечество, и особенно на своих соотечественников, когда он показал, что свободная конкуренция, борьба ибо существование, которое экономисты прославляют как высшее историческое достижение, есть нормальное состояние животного царства ». [124] Герберт Спенсер и сторонник евгеники, интерпретация Фрэнсиса Гальтона естественного отбора как обязательно прогрессивного, ведущего к предполагаемому прогрессу в интеллекте и цивилизации, стала оправданием колониализма , евгеники и социального дарвинизма . Например, в 1940 году Конрад Лоренц в трудах, от которых впоследствии отрекся, использовал эту теорию как оправдание политики нацистского государства . Он писал: «... отбор на стойкость, героизм и социальную полезность... должен осуществляться каким-то человеческим институтом, если человечество, при отсутствии факторов отбора, не будет разрушено вырождением, вызванным одомашниванием. Расовая идея как основа нашего государства уже многого добилась в этом отношении». [125] Другие разработали идеи о том, что человеческие общества и культура развиваются по механизмам, аналогичным тем, которые применяются к эволюции видов. [126]
Совсем недавно работа антропологов и психологов привела к развитию социобиологии , а затем и эволюционной психологии, области, которая пытается объяснить особенности психологии человека с точки зрения адаптации к окружающей среде предков. Наиболее ярким примером эволюционной психологии, особенно развитым в ранних работах Хомского , а затем Стивена Пинкера , является гипотеза о том, что человеческий мозг адаптировался к усвоению грамматических Ноама правил естественного языка . [127] Было высказано предположение, что другие аспекты человеческого поведения и социальных структур, от конкретных культурных норм, таких как избегание инцеста , до более широких моделей, таких как гендерные роли , имеют схожее происхождение как адаптация к ранней среде, в которой развивались современные люди. По аналогии с действием естественного отбора на гены возникла концепция мемов — «единиц культурной передачи» или культурных эквивалентов генов, подвергающихся отбору и рекомбинации, впервые описанная в этой форме Ричардом Докинзом в 1976 году. [128] и впоследствии расширены такими философами, как Дэниел Деннет, для объяснения сложной культурной деятельности, включая человеческое сознание . [129]
Теория информации и систем
В 1922 году Альфред Дж. Лотка предположил, что естественный отбор можно понимать как физический принцип, который можно описать с точки зрения использования системой энергии. [130] [131] концепция, позже разработанная Говардом Т. Одумом как принцип максимальной мощности в термодинамике , согласно которому эволюционные системы с избирательным преимуществом максимизируют скорость преобразования полезной энергии. [132]
Принципы естественного отбора вдохновили на создание множества вычислительных методов, таких как «мягкая» искусственная жизнь , которая моделирует процессы отбора и может быть высокоэффективной в «адаптации» объектов к среде, определяемой заданной функцией приспособленности . [133] Например, класс эвристических оптимизации алгоритмов , известный как генетические алгоритмы , впервые разработанный Джоном Генри Холландом в 1970-х годах и расширенный Дэвидом Э. Голдбергом , [134] выявлять оптимальные решения путем моделирования воспроизводства и мутации совокупности решений, определенных начальным распределением вероятностей . [135] Такие алгоритмы особенно полезны при применении к задачам, энергетический ландшафт которых очень груб или имеет множество локальных минимумов. [136]
В художественной литературе
Дарвиновская теория эволюции посредством естественного отбора широко распространена в литературе, как с оптимистической точки зрения, с точки зрения того, как человечество может развиваться в направлении совершенства, так и с пессимистической точки зрения, с точки зрения ужасных последствий взаимодействия человеческой природы и борьбы за выживание. Среди основных откликов — Сэмюэля Батлера 1872 года пессимистический «Эревон» («Нигде», написанный в основном задом наперед). В 1893 году Герберт Уэллс представил себе « Человека года на миллион », превращенного в результате естественного отбора в существо с огромной головой, глазами и сморщенным телом. [137]
Примечания
- ^ При половом отборе можно утверждать, что самка животного, выбирающая партнера, намеревается получить лучшего партнера; нет никаких предположений, что она намерена улучшить родословную в манере заводчика животных.
Ссылки
- ^ Эмпедокл 1898 , О природе , Книга II
- ^ Лукреций 1916 , О природе вещей , Книга V
- ^ Аристотель , Физика , Книга II, главы 4 и 8.
- ^ Лир 1988 , с. 38
- ^ Генри, Девин (сентябрь 2006 г.). «Аристотель о механизме наследования» . Журнал истории биологии . 39 (3): 425–455. дои : 10.1007/s10739-005-3058-y . S2CID 85671523 .
- ^ Ариев 2002
- ^ Дарвин 1872 , с. xiii
- ^ Аристотель , Физика , Книга II, Глава 8.
- ^ Зиркль, Конвей (25 апреля 1941 г.). «Естественный отбор до возникновения видов ». Труды Американского философского общества . 84 (1): 71–123. JSTOR 984852 .
- ^ Агаттер и Уитли 2008 , с. 43
- ^ Леонардо, Кодекс C. Институт Франции. Пер. Рихтер. 2016.
- ^ Мопертюи, Пьер Луи (1746). [ «Вывод законов движения и равновесия из метафизического принципа» ]. История Королевской академии наук и изящной словесности (на французском языке). Берлин: 267–294.
- ^ Боулер, Питер Дж. (2003). Эволюция: история идеи (3-е изд.). Беркли, Калифорния: Издательство Калифорнийского университета . стр. 129–134 . ISBN 978-0-520-23693-6 . ОСЛК 43091892 .
- ^ Ламарк 1809 г.
- ^ Джоравски, Дэвид (январь 1959 г.). «Советский марксизм и биология до Лысенко». Журнал истории идей . 20 (1): 85–104. дои : 10.2307/2707968 . JSTOR 2707968 .
- ^ Дарвин 1859 , с. 18
- ^ Дарвин 1859 , с. 61
- ^ Дарвин 1859 , с. 5
- ^ Холл, Брайан К.; Халлгримссон, Бенедикт (2008). Эволюция Стрикбергера (4-е изд.). Джонс и Бартлетт. стр. 4–6. ISBN 978-0-7637-0066-9 . OCLC 796450355 .
- ^ Мальтус 1798 г.
- ^ Дарвин 1958 , с. 120
- ^ Дарвин 1859 , стр. 126–127.
- ^ Уоллес 1871 г.
- ^ Дарвин 1861 , с. xiii
- ^ Дарвин 1859 , с. 6
- ^ Дарвин, Чарльз (28 сентября 1860 г.). «Дарвин, ЧР — Лайеллю, Чарльзу» . Дарвиновский заочный проект . Кембридж, Великобритания: Библиотека Кембриджского университета . Письмо 2931 . Проверено 1 августа 2015 г.
- ↑ Перейти обратно: Перейти обратно: а б Дарвин 1859 г.
- ^ Эйсли 1958
- ^ Кун 1996
- ^ Дарвин, Чарльз (5 июля 1866 г.). «Дарвин, ЧР — Уоллесу, Арканзас» Дарвинский корреспондентский проект . Кембридж, Великобритания: Библиотека Кембриджского университета. Письмо 5145 . Проверено 12 января 2010 г.
- ^ Штуке, Морис Э. (лето 2008 г.). «Лучшая защита конкуренции» . Обзор закона Святого Иоанна . 82 (3). Ямайка, Нью-Йорк: 951–1036.
Это выживание наиболее приспособленных, которое я здесь пытался выразить в механических терминах, есть то, что г-н Дарвин назвал «естественным отбором, или сохранением избранных рас в борьбе за жизнь».
- Герберт Спенсер , Принципы биологии (1864), том. 1, стр. 444–445. - ^ Дарвин 1872 , с. 49 .
- ^ Миллс, Сьюзен К.; Битти, Джон Х. (1979). «Интерпретация фитнеса по склонности» (PDF) . Философия науки . 46 (2): 263–286. CiteSeerX 10.1.1.332.697 . дои : 10.1086/288865 . S2CID 38015862 . Архивировано из оригинала (PDF) 25 декабря 2015 года . Проверено 4 августа 2015 г.
- ^ Эмброуз, Майк. «Горох Менделя» . Норидж, Великобритания: Отдел ресурсов зародышевой плазмы, Центр Джона Иннеса . Архивировано из оригинала 14 июня 2016 года . Проверено 22 мая 2015 г.
- ^ Хаксли, Джулиан (1929–1930). «Азбука генетики». Наука жизни . Том. 2. Лондон: Объединенная пресса . ОСЛК 3171056 .
- ^ Национальная академия наук (1999). Наука и креационизм: взгляд Национальной академии наук (2-е изд.). Вашингтон, округ Колумбия: Издательство Национальной академии. ISBN 978-0-309-06406-4 . OCLC 43803228 .
- ↑ Перейти обратно: Перейти обратно: а б Фишер 1930 г.
- ^ Холдейн 1932 г.
- ^ Холдейн, JBS (декабрь 1957 г.). «Цена естественного отбора» (PDF) . Журнал генетики . 55 (3): 511–524. дои : 10.1007/BF02984069 . S2CID 32233460 .
- ^ Райт, Сьюэлл (1932). «Роль мутации, инбридинга, скрещивания и отбора в эволюции» . Материалы VI Международного генетического конгресса . 1 : 356–366.
- ^ Добжанский 1937 г.
- ^ Добжанский 1951 г.
- ^ Кэрролл, Шон Б.; Гренье, Дженнифер К.; Уэзерби, Скотт Д. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных – второе издание . Издательство Блэквелл. стр. 66–67. ISBN 978-1-4051-1950-4 .
- ^ Майр 1942 г.
- ^ Гамильтон, В. (1964). «Генетическая эволюция социального поведения. Я». Журнал теоретической биологии . 7 (1): 1–16. Бибкод : 1964JThBi...7....1H . дои : 10.1016/0022-5193(64)90038-4 . ПМИД 5875341 . S2CID 5310280 .
- ^ Гамильтон, В. (1964). «Генетическая эволюция социального поведения. II». Журнал теоретической биологии . 7 (1): 17–52. Бибкод : 1964JThBi...7...17H . дои : 10.1016/0022-5193(64)90039-6 . ПМИД 5875340 .
- ^ Гилберт, Скотт Ф. (2003). «Морфогенез эволюционной биологии развития» (PDF) . Международный журнал биологии развития . 47 (7–8): 467–477. ПМИД 14756322 .
- ^ Гилберт, Сан-Франциско; Опиц, Дж. М.; Рафф, РА (1996). «Ресинтез эволюционной биологии и биологии развития» . Биология развития . 173 (2): 357–372. дои : 10.1006/dbio.1996.0032 . ПМИД 8605997 .
- ^ Мюллер, Великобритания (2007). «Эво-дево: расширение эволюционного синтеза». Обзоры природы Генетика . 8 (12): 943–949. дои : 10.1038/nrg2219 . ПМИД 17984972 . S2CID 19264907 .
- ^ Кэрролл, Шон Б.; Гренье, Дженнифер К.; Уэзерби, Скотт Д. (2005). От ДНК к разнообразию: молекулярная генетика и эволюция дизайна животных – второе издание . Издательство Блэквелл. п. 13. ISBN 978-1-4051-1950-4 .
- ^ Уильямс 1966
- ^ Эндлер 1986
- ^ Холдейн 1954 г.
- ^ Ланде, Рассел ; Арнольд, Стеван Дж. (ноябрь 1983 г.). «Измерение отбора коррелирующих символов». Эволюция . 37 (6): 1210–1226. дои : 10.2307/2408842 . JSTOR 2408842 . ПМИД 28556011 .
- ^ Футуйма 2005 г.
- ^ Трезвый 1993 г.
- ↑ Перейти обратно: Перейти обратно: а б с «Эволюция и естественный отбор» . Мичиганский университет. 10 октября 2010 г. Проверено 9 ноября 2016 г.
- ^ «Телеологические понятия в биологии» . Стэнфордская энциклопедия философии . 18 мая 2003 года . Проверено 28 июля 2016 г.
- ^ Вант Хоф, Арьен Э.; Кампань, Паскаль; Ригден, Дэниел Дж; и др. (июнь 2016 г.). «Мутация промышленного меланизма у британской перцовой моли является мобильным элементом». Природа . 534 (7605): 102–105. Бибкод : 2016Natur.534..102H . дои : 10.1038/nature17951 . ПМИД 27251284 . S2CID 3989607 .
- ^ Уолтон, Оливия; Стивенс, Мартин (2018). «Модели птичьего зрения и полевые эксперименты определяют выживаемость камуфляжа перцовой моли» . Коммуникационная биология . 1 : 118. дои : 10.1038/s42003-018-0126-3 . ПМК 6123793 . ПМИД 30271998 .
- ^ Орр, Х. Аллен (август 2009 г.). «Приспособленность и ее роль в эволюционной генетике» . Нат преподобный Жене . 10 (8): 531–539. дои : 10.1038/nrg2603 . ПМЦ 2753274 . ПМИД 19546856 .
- ^ Холдейн, JBS (ноябрь 1992 г.). «Цена естественного отбора». Современная наука . 63 (9/10): 612–625.
- ^ Кляйнман, А. (2014). «Фундаментальная наука и математика случайных мутаций и естественного отбора» . Статистика в медицине . 33 (29): 5074–5080. дои : 10.1002/sim.6307 . ПМИД 25244620 .
- ^ Байм, М.; Либерман, Т.Д.; Келсик, Эд; Хаит, Р.; Гросс, Р.; Елин И.; Кишони, Р. (2016). «Пространственно-временная эволюция микробов на ландшафтах антибиотиков» . Наука . 353 (6304): 1147–51. Бибкод : 2016Sci...353.1147B . дои : 10.1126/science.aag0822 . ПМЦ 5534434 . ПМИД 27609891 .
- ^ Блаунт, Закари Д.; Борланд, Кристина З.; Ленски, Ричард Э. (2008). «Историческая случайность и эволюция ключевой инновации в экспериментальной популяции Escherichia coli » . Труды Национальной академии наук . 105 (23): 7899–906. Бибкод : 2008PNAS..105.7899B . дои : 10.1073/pnas.0803151105 . JSTOR 25462703 . ПМЦ 2430337 . ПМИД 18524956 .
- ^ Хорошо, Б.Х.; Рузин, И.М.; Балик, диджей; Халлачек, О.; Десаи, ММ (27 февраля 2012 г.). «Распределение фиксированных полезных мутаций и скорость адаптации в бесполых популяциях» . Труды Национальной академии наук . 109 (13): 4950–4955. дои : 10.1073/pnas.1119910109 . ПМЦ 3323973 . ПМИД 22371564 .
- ^ Бегон, Таунсенд и Харпер, 1996 г.
- ↑ Перейти обратно: Перейти обратно: а б Сахни, Сарда; Бентон, Майкл Дж .; Ферри, Пол А. (23 августа 2010 г.). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Джардин, Филипп Э.; Дженис, Кристин М.; Сахни, Сарда; Бентон, Майкл Дж. (1 декабря 2012 г.). «Песчинка, а не трава: согласованные модели раннего происхождения гипсодонтии у копытных Великих равнин и глайров». Палеогеография, Палеоклиматология, Палеоэкология . 365–366: 1–10. Бибкод : 2012PPP...365....1J . дои : 10.1016/j.palaeo.2012.09.001 .
- ^ Макартур и Уилсон, 2001 г.
- ^ Пианка, Эрик Р. (ноябрь – декабрь 1970 г.). «О р- и К -отборе». Американский натуралист . 104 (940): 592–597. дои : 10.1086/282697 . JSTOR 2459020 . S2CID 83933177 .
- ↑ Перейти обратно: Перейти обратно: а б Ферхюльст, Пьер Франсуа (1838). « Заметьте по закону, что население следует в своем росте » . Математическая и физическая переписка (на французском языке). 10 . Брюссель, Бельгия: 113–121. OCLC 490225808 .
- ^ Циммер и Эмлен, 2013 г.
- ^ Миллер 2000 , с. 8
- ^ Арнквист, Йоран; Роу, Локк (2005). Сексуальный конфликт . Издательство Принстонского университета. стр. 14–43. ISBN 978-0-691-12218-2 . OCLC 937342534 .
- ^ Леми, Салеми и Вандамм, 2009 г.
- ^ Лоу, Лоуренс (2008). «Негативный отбор» . Природное образование . Кембридж, Массачусетс: Издательская группа Nature . OCLC 310450541 .
- ^ Вилланеа, Фернандо А.; Сафи, Кристин Н.; Буш, Иеремия В. (май 2015 г.). «Общая модель отрицательного частотно-зависимого отбора объясняет глобальные закономерности полиморфизма АВО человека» . ПЛОС ОДИН . 10 (5): e0125003. Бибкод : 2015PLoSO..1025003V . дои : 10.1371/journal.pone.0125003 . ПМЦ 4422588 . ПМИД 25946124 .
- ↑ Перейти обратно: Перейти обратно: а б Кристиансен 1984 , стр. 65–79.
- ^ Уэйд, Майкл Дж.; и др. (2010). «Многоуровневость и родственный отбор в связанном мире» . Природа . 463 (7283): Е8–Е9. Бибкод : 2010Natur.463....8W . дои : 10.1038/nature08809 . ПМК 3151728 . ПМИД 20164866 .
- ↑ Перейти обратно: Перейти обратно: а б Дарвин, Чарльз (1859). О происхождении видов (1-е издание). Глава 4, стр. 88. «И это заставляет меня сказать несколько слов о том, что я называю половым отбором. Это зависит…» http://darwin-online.org.uk/content/frameset?viewtype=side&itemID=F373&pageseq =12
- ↑ Перейти обратно: Перейти обратно: а б Гринфилд, доктор медицины; Алем, С.; Лимузен, Д.; Бейли, Северо-Запад (2014). «Дилемма фишеровского полового отбора: выбор партнера ради косвенной выгоды, несмотря на редкость и общую слабость генетической корреляции между чертами и предпочтениями» . Эволюция . 68 (12): 3524–3536. дои : 10.1111/evo.12542 . ПМИД 25308282 . S2CID 2619084 .
- ^ Майр 2006 г.
- ^ Андерсон 1994
- ↑ Перейти обратно: Перейти обратно: а б Хоскен, Дэвид Дж.; Хаус, Кларисса М. (январь 2011 г.). «Половой отбор» . Современная биология . 21 (2): Р62–Р65. Бибкод : 2011CBio...21..R62H . дои : 10.1016/j.cub.2010.11.053 . ПМИД 21256434 . S2CID 18470445 .
- ^ Энс, Марсель; Пинкстен, Рианна (5 октября 2000 г.). «Смена половых ролей у позвоночных: поведенческие и эндокринологические аспекты». Поведенческие процессы . 51 (1–3): 135–147. дои : 10.1016/S0376-6357(00)00124-8 . ПМИД 11074317 . S2CID 20732874 .
- ^ Барлоу, Джордж У. (март 2005 г.). «Как мы решаем, что у вида поменяна половая роль?». Ежеквартальный обзор биологии . 80 (1): 28–35. дои : 10.1086/431022 . ПМИД 15884733 . S2CID 44774132 .
- ^ Харви, Фиона; Карсон, Мэри; О'Кейн, Мэгги; Уэсли, Эндрю (18 июня 2015 г.). «Супербактерия MRSA, обнаруженная в свинине в супермаркетах, вызывает тревогу по поводу рисков, связанных с сельским хозяйством» . Хранитель .
- ^ Шито, Джан К. (март 2006 г.). «Важность развития устойчивости золотистого стафилококка к антибиотикам » . Клиническая микробиология и инфекции . 12 (Приложение 1): 3–8. дои : 10.1111/j.1469-0691.2006.01343.x . ПМИД 16445718 .
- ^ Шарла, Сильвен; Хорнетт, Эмили А.; Фуллард, Джеймс Х.; и др. (13 июля 2007 г.). «Необычайное изменение соотношения полов». Наука . 317 (5835): 214. Бибкод : 2007Sci...317..214C . дои : 10.1126/science.1143369 . ПМИД 17626876 . S2CID 45723069 .
- ^ Моран, Грегори Дж.; и др. (2006). «Метициллин-резистентные инфекции, вызванные S. Aureus, у пациентов отделения неотложной помощи» . Медицинский журнал Новой Англии . 355 (7): 666–674. doi : 10.1056/NEJMoa055356 . ПМИД 16914702 .
- ^ Крюков Григорий Васильевич; Шмидт, Штеффен; Сюняев, Шамиль (1 августа 2005 г.). «Небольшой эффект приспособленности мутаций в высококонсервативных некодирующих регионах» . Молекулярная генетика человека . 14 (15): 2221–2229. дои : 10.1093/hmg/ddi226 . ПМИД 15994173 .
- ^ Беджерано, Джилл; Фазан, Майкл; Макунин Игорь; и др. (28 мая 2004 г.). «Ультраконсервативные элементы в геноме человека» (PDF) . Наука . 304 (5675): 1321–1325. Бибкод : 2004Sci...304.1321B . CiteSeerX 10.1.1.380.9305 . дои : 10.1126/science.1098119 . ПМИД 15131266 . S2CID 2790337 .
- ^ Эйр-Уокер, Адам; Вулфит, Меган; Фелпс, Тед (июнь 2006 г.). «Распределение фитнес-эффектов новых вредных мутаций аминокислот у людей» . Генетика . 173 (2): 891–900. doi : 10.1534/genetics.106.057570 . ПМК 1526495 . ПМИД 16547091 .
- ^ Галис, Фритсон (апрель 1999 г.). «Почему почти у всех млекопитающих семь шейных позвонков? Ограничения развития, Hox -гены и рак». Журнал экспериментальной зоологии . 285 (1): 19–26. Бибкод : 1999JEZ...285...19G . doi : 10.1002/(SICI)1097-010X(19990415)285:1<19::AID-JEZ3>3.0.CO;2-Z . ПМИД 10327647 .
- ^ Закань, Йожеф; Фроманталь-Рамен, Катрин; Варот, Ксавье; Дюбул, Дени (9 декабря 1997 г.). «Регуляция количества и размера пальцев задними Hox- генами: дозозависимый механизм с потенциальными эволюционными последствиями» . Труды Национальной академии наук Соединенных Штатов Америки . 94 (25): 13695–13700. Бибкод : 1997PNAS...9413695Z . дои : 10.1073/pnas.94.25.13695 . ПМК 28368 . ПМИД 9391088 .
- ^ Саньял, Сомес; Янсен, Гарри Г.; де Грип, Виллем Дж.; Нево, Эвиатар ; и др. (июль 1990 г.). «Глаз слепого землекопа, Spalax ehrenbergi . Рудимент со скрытой функцией?» . Исследовательская офтальмология и визуальные науки . 31 (7): 1398–1404. ПМИД 2142147 .
- ^ Орр, ХА (1996). «Добжанский, Бейтсон и генетика видообразования» . Генетика . 144 (4): 1331–1335. дои : 10.1093/генетика/144.4.1331 . ПМК 1207686 . ПМИД 8978022 .
- ^ Гаврилец, С. (2004), Фитнес-ландшафты и происхождение видов , Princeton University Press, ISBN 978-0-691-11983-0
- ^ Шулер, Ханнес; Худ, Глен Р.; Иган, Скотт П.; Федер, Джеффри Л. (2016). «Способы и механизмы видообразования». Обзоры по клеточной биологии и молекулярной медицине . 2 (3): 60–93.
- ^ МакКьюсик, Виктор А.; Гросс, Мэтью Б. (18 ноября 2014 г.). «АВО-гликозилтрансфераза; АВО» . Интернет-менделевское наследование у человека . Национальная медицинская библиотека . Проверено 7 ноября 2016 г.
- ^ Фальконер и Маккей, 1996 г.
- ↑ Перейти обратно: Перейти обратно: а б Rice 2004. Количественный анализ см., в частности, в главах 5 и 6.
- ^ Ризеберг, Л.Х.; Видмер, А.; Арнц, AM; Берк, Дж. М. (2002). «Направленный отбор является основной причиной фенотипической диверсификации» . ПНАС . 99 (19): 12242–12245. Бибкод : 2002PNAS...9912242R . дои : 10.1073/pnas.192360899 . ПМК 129429 . ПМИД 12221290 .
- ^ Чарльзуорт Б., Ланде Р., Слаткин М. (1982). «Неодарвинистский комментарий к макроэволюции» . Эволюция . 36 (3): 474–498. дои : 10.1111/j.1558-5646.1982.tb05068.x . JSTOR 2408095 . ПМИД 28568049 . S2CID 27361293 .
- ^ Гамильтон, Уильям Д. (июль 1964 г.). «Генетическая эволюция социального поведения. II». Журнал теоретической биологии . 7 (1): 17–52. Бибкод : 1964JThBi...7...17H . дои : 10.1016/0022-5193(64)90039-6 . ПМИД 5875340 .
- ^ Триверс, Роберт Л. (март 1971 г.). «Эволюция взаимного альтруизма». Ежеквартальный обзор биологии . 46 (1): 35–57. дои : 10.1086/406755 . JSTOR 2822435 . S2CID 19027999 .
- ^ Кимура, Мотоо (1983). Нейтральная теория молекулярной эволюции . Издательство Кембриджского университета. ISBN 978-0-521-23109-1 . OCLC 8776549 .
- ^ Робинсон, Ричард, изд. (2003). «Узкое место народонаселения» . Генетика . Том. 3. Справочник Macmillan, США. ISBN 978-0-02-865609-0 . OCLC 3373856121 .
- ^ Кэмпбелл, Нил А. (1996). Биология (4-е изд.). Бенджамин Каммингс . п. 423 . ISBN 978-0-8053-1940-8 . OCLC 3138680061 .
- ^ Линч, Майкл (август 2010 г.). «Эволюция скорости мутаций» . Тенденции в генетике . 26 (8): 345–352. дои : 10.1016/j.tig.2010.05.003 . ПМЦ 2910838 . ПМИД 20594608 .
- ^ Смит, Джон Мейнард ; Хей, Джон (1974). «Автостопный эффект благоприятного гена» . Генетические исследования . 23 (1): 23–35. дои : 10.1017/S0016672300014634 . ПМИД 4407212 .
- ^ Кейтли, Питер Д .; Отто, Сара П. (7 сентября 2006 г.). «Вмешательство в вредные мутации благоприятствует сексу и рекомбинации в ограниченных популяциях». Природа . 443 (7107): 89–92. Бибкод : 2006Natur.443...89K . дои : 10.1038/nature05049 . ПМИД 16957730 . S2CID 4422532 .
- ^ Дарвин 1859 , с. 489
- ^ Гулд, Стивен Джей (12 июня 1997 г.). «Дарвиновский фундаментализм» . Нью-Йоркское обозрение книг . 44 (10).
- ^ Деннетт 1995
- ^ фон Сюдов, М. (2012). От дарвиновской метафизики к пониманию эволюции эволюционных механизмов. Историко-философский анализ генного дарвинизма и универсального дарвинизма . Университет Геттингена. ISBN 978-3-86395-006-4 . ОСЛК 1088022023 .
- ^ Эйген, Манфред ; Гардинер, Уильям; Шустер, Питер ; и др. (апрель 1981 г.). «Происхождение генетической информации». Научный американец . 244 (4): 88–92, 96 и др . Бибкод : 1981SciAm.244d..88E . doi : 10.1038/scientificamerican0481-88 . ПМИД 6164094 .
- ↑ Перейти обратно: Перейти обратно: а б Бернштейн, Харрис; Байерли, Генри К.; Хопф, Фредерик А.; и др. (июнь 1983 г.). «Дарвиновская динамика». Ежеквартальный обзор биологии . 58 (2): 185–207. дои : 10.1086/413216 . JSTOR 2828805 . S2CID 83956410 .
- ↑ Перейти обратно: Перейти обратно: а б Мишо 1999 г.
- ^ Оргел, Лесли Э. (1987). «Эволюция генетического аппарата: обзор». Симпозиумы Колд-Спринг-Харбор по количественной биологии . 52 : 9–16. дои : 10.1101/sqb.1987.052.01.004 . ПМИД 2456886 .
- ^ Ру 1881 г.
- ^ Купец, Жан-Жак [на французском языке] (3 мая 2010 г.). «Клеточный дарвинизм (стохастическая экспрессия генов при дифференцировке клеток и развитии эмбрионов)» . Научные темы . Архивировано из оригинала 4 августа 2010 года . Проверено 11 августа 2015 г.
- ^ Энгельс 1964 г.
- ^ Айзенберг, Леон (сентябрь 2005 г.). «Какой образ для Лоренца?». Американский журнал психиатрии (письмо в редакцию). 162 (9): 1760. doi : 10.1176/appi.ajp.162.9.1760 . ПМИД 16135651 . Айзенберг цитирует перевод книги «Нарушения видоспецифического поведения, вызванные одомашниванием» (1940, стр. 2) Конрада Лоренца .
- ^ Уилсон 2002
- ^ Пинкер 1995
- ^ Докинз 1976 , с. 192
- ^ Деннетт 1991
- ^ Лотка, Альфред Дж. (июнь 1922 г.). «Вклад в энергетику эволюции» . ПНАС . 8 (6): 147–151. Бибкод : 1922PNAS....8..147L . дои : 10.1073/pnas.8.6.147 . ПМЦ 1085052 . ПМИД 16576642 .
- ^ Лотка, Альфред Дж. (июнь 1922 г.). «Естественный отбор как физический принцип» . ПНАС . 8 (6): 151–154. Бибкод : 1922PNAS....8..151L . дои : 10.1073/pnas.8.6.151 . ПМК 1085053 . ПМИД 16576643 .
- ^ Одум, ХТ (1995). Холл, CAS (ред.). Самоорганизация и максимальная сила . Издательство Университета Колорадо.
- ^ Кауфман 1993
- ^ Гольдберг 1989
- ^ Митчелл 1996
- ^ «Генетические алгоритмы» . Фармакологические науки . 7 ноября 2016 г. Проверено 7 ноября 2016 г.
- ^ Стейблфорд, Брайан М.; Лэнгфорд, Дэвид Р. (5 июля 2018 г.). «Эволюция» . Энциклопедия научной фантастики . Голланц . Проверено 24 июля 2018 г.
Источники
- Агаттер, Пол С.; Уитли, Денис Н. (2008). Размышление о жизни: история и философия биологии и других наук . Дордрехт, Нидерланды; Лондон: Springer Science+Business Media . ISBN 978-1-4020-8865-0 . LCCN 2008933269 . OCLC 304561132 .
- Андерссон, Мальте (1994). Половой отбор . Монографии по поведению и экологии. Принстон, Нью-Джерси: Издательство Принстонского университета . ISBN 978-0-691-00057-2 . LCCN 93033276 . OCLC 28891551 .
- Ариев, Андре (2002). «Платонические и аристотелевские корни телеологических аргументов» (PDF) . В Арье, Андре; Камминс, Роберт; Перлман, Марк (ред.). Функции: Новые очерки философии психологии и биологии . Оксфорд; Нью-Йорк: Издательство Оксфордского университета . ISBN 978-0-19-824103-4 . LCCN 2002020184 . OCLC 48965141 . Архивировано из оригинала (PDF) 19 февраля 2009 года.
- Аристотель . Физика . Перевод Р. П. Харди и Р. К. Гэй. Архив интернет-классики. OCLC 54350394 .
- Бегон, Майкл; Таунсенд, Колин Р.; Харпер, Джон Л. (1996). Экология: Индивиды, популяции и сообщества (3-е изд.). Оксфорд; Кембридж, Массачусетс: Blackwell Science . ISBN 978-0-632-03801-5 . LCCN 95024627 . OCLC 32893848 .
- Кристиансен, Фредди Б. (1984). «Определение и измерение фитнеса» . В Шорроксе, Брайан (ред.). Эволюционная экология: 23-й симпозиум Британского экологического общества, Лидс, 1982 . Симпозиум Британского экологического общества . Том. 23. Оксфорд; Бостон: Научные публикации Блэквелла . ISBN 978-0-632-01189-6 . LCCN 85106855 . OCLC 12586581 . Модифицирован Кристиансена путем добавления отбора на выживание на репродуктивной фазе.
- Дарвин, Чарльз (1859). О происхождении видов путем естественного отбора, или о сохранении избранных рас в борьбе за жизнь (1-е изд.). Лондон: Джон Мюррей . LCCN 06017473 . OCLC 741260650 . Книга доступна в Полном собрании сочинений Чарльза Дарвина в Интернете . Проверено 23 июля 2015 г.
- Дарвин, Чарльз (1861). О происхождении видов путем естественного отбора, или сохранение избранных рас в борьбе за жизнь (3-е изд.). Лондон: Джон Мюррей. LCCN 04001284 . OCLC 550913 .
- Дарвин, Чарльз (1872). Происхождение видов путем естественного отбора, или сохранение избранных рас в борьбе за жизнь (6-е изд.). Лондон: Джон Мюррей. OCLC 1185571 .
- Дарвин, Чарльз (1958). Барлоу, Нора (ред.). Автобиография Чарльза Дарвина, 1809–1882: с восстановленными первоначальными упущениями; Отредактировано, с приложениями и примечаниями его внучки Норы Барлоу . Лондон: Коллинз . LCCN 93017940 . OCLC 869541868 .
- Докинз, Ричард (1976). Эгоистичный ген . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-857519-1 . LCCN 76029168 . ОСЛК 2681149 .
- Деннетт, Дэниел К. (1991). Объяснение сознания (1-е изд.). Бостон, Массачусетс: Литтл, Браун и компания . ISBN 978-0-316-18065-8 . LCCN 91015614 . OCLC 23648691 .
- Деннетт, Дэниел К. (1995). Опасная идея Дарвина: эволюция и смысл жизни . Нью-Йорк: Саймон и Шустер . ISBN 978-0-684-80290-9 . LCCN 94049158 . OCLC 31867409 .
- Добжанский, Феодосий (1937). Генетика и происхождение видов . Биологическая серия Колумбийского университета . Нью-Йорк: Издательство Колумбийского университета . LCCN 37033383 . OCLC 766405 .
- —— (1951). Генетика и происхождение видов . Биологическая серия Колумбийского университета (3-е исправленное издание). Нью-Йорк: Издательство Колумбийского университета. LCCN 51014816 . OCLC 295774 .
- Эйсли, Лорен (1958). Век Дарвина: эволюция и люди, которые ее открыли (1-е изд.). Гарден-Сити, Нью-Йорк: Даблдей . LCCN 58006638 . OCLC 168989 .
- Эмпедокл (1898 г.). «Эмпедокл» . В Фэрбенксе, Артур (ред.). Первые философы Греции . Перевод Артура Фэрбенкса. Лондон: Кеган Пол, Trench, Trübner & Co. Ltd. LCCN 03031810 . ОСЛК 1376248 . Первые философы Греции в Интернет-архиве .
- Эндлер, Джон А. (1986). Естественный отбор в дикой природе . Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-08386-5 . LCCN 85042683 . OCLC 12262762 .
- Энгельс, Фридрих (1964) [1883]. Диалектика природы . Предисловие Дж.Б.С. Холдейна , 1939 г. (3-е изд.). Москва, СССР: Издательство «Прогресс» . LCCN 66044448 . OCLC 807047245 . Книга доступна в Марксистском Интернет-архиве .
- Фальконер, Дуглас С .; Маккей, Труди (1996). Введение в количественную генетику (4-е изд.). Харлоу, Англия: Лонгман . ISBN 978-0-582-24302-6 . OCLC 824656731 .
- Фишер, Рональд Эйлмер (1930). Генетическая теория естественного отбора . Оксфорд: Кларендон Пресс . LCCN 30029177 . OCLC 493745635 .
- Футуйма, Дуглас Дж. (2005). Эволюция . Сандерленд, Массачусетс: Sinauer Associates . ISBN 978-0-87893-187-3 . LCCN 2004029808 . OCLC 57311264 .
- Голдберг, Дэвид Э. (1989). Генетические алгоритмы в поиске, оптимизации и машинном обучении . Ридинг, Массачусетс: Издательская компания Addison-Wesley . ISBN 978-0-201-15767-3 . LCCN 88006276 . OCLC 17674450 .
- Холдейн, Дж. Б.С. (1932). Причины эволюции . Лондон; Нью-Йорк: Longmans, Green & Co. LCCN 32033284 . OCLC 5006266 . «Эта книга основана на серии лекций, прочитанных в январе 1931 года в Прифисгол Симру, Аберистуит, и озаглавлена «Пересмотр дарвинизма».
- Холдейн, JBS (1954). «Измерение естественного отбора» . В Монталенти, Джузеппе; Кьяруги, А. (ред.). Материалы 9-го Международного конгресса генетиков, Белладжио (Комо), 24–31 августа 1953 г. [ Труды 9-го Международного конгресса генетиков ]. Кариология. Том 6 (1953/54) Доп. Флоренция, Италия: Флорентийский университет . стр. 480–487. ОСЛК 9069245 .
- Кауфман, Стюарт (1993). Истоки порядка: самоорганизация и отбор в эволюции . Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-507951-7 . LCCN 91011148 . ОСЛК 23253930 .
- Ламарк, Жан-Батист (1809). Зоологическая философия . Париж: Денту и автор. ОСЛК 2210044 . Зоологическая философия (1809 г.) в Интернет-архиве .
- Лир, Джонатан (1988). Аристотель: Желание понять . Кембридж, Великобритания; Нью-Йорк: Издательство Кембриджского университета . ISBN 978-0-521-34762-4 . LCCN 87020284 . OCLC 16352317 .
- Кун, Томас С. (1996). Структура научных революций (3-е изд.). Чикаго, Иллинойс: Издательство Чикагского университета. ISBN 978-0-226-45808-3 . LCCN 96013195 . OCLC 34548541 .
- Леми, Филипп; Салеми, Марко; Вандам, Анн-Мик, ред. (2009). Филогенетический справочник: практический подход к филогенетическому анализу и проверке гипотез (2-е изд.). Кембридж, Великобритания; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-73071-6 . LCCN 2009464132 . ОСЛК 295002266 .
- Лукреций (1916). «Книга V». В Леонарде, Уильям Эллери (ред.). De rerum natura . Перевод Уильяма Эллери Леонарда. Медфорд/Сомервилл, Массачусетс: Университет Тафтса . OCLC 33233743 .
- Макартур, Роберт Х .; Уилсон, Эдвард О. (2001) [первоначально опубликовано в 1967 году]. Теория островной биогеографии . Принстонские достопримечательности в области биологии. Новое предисловие Эдварда О. Уилсона. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-08836-5 . LCCN 00051495 . ОСЛК 45202069 .
- Мальтус, Томас Роберт (1798). Очерк о принципе народонаселения, как он влияет на будущее улучшение общества: с замечаниями по поводу размышлений г-на Годвина, М. Кондорсе и других писателей (1-е изд.). Лондон: Дж. Джонсон. LCCN 46038215 . OCLC 65344349 . Книгу можно приобрести здесь у Фрэнка Элвелла, Государственный университет Роджерса .
- Майр, Эрнст (1942). Систематика и происхождение видов с точки зрения зоолога . Колумбийская биологическая серия. Том. 13. Нью-Йорк: Издательство Колумбийского университета. LCCN 43001098 . OCLC 766053 .
- Майр, Эрнст (2006) [Первоначально опубликовано в 1972 году; Чикаго, Иллинойс: Aldine Publishing Co.]. «Половой отбор и естественный отбор». В Кэмпбелле, Бернард Г. (ред.). Половой отбор и происхождение человека: дарвиновский поворот . Нью-Брансуик, Нью-Джерси: AldineTransaction . ISBN 978-0-202-30845-6 . LCCN 2005046652 . OCLC 62857839 .
- Мишод, Ричард А. (1999). Дарвиновская динамика: эволюционные переходы в приспособленности и индивидуальности . Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-02699-2 . LCCN 98004166 . OCLC 38948118 .
- Миллер, Джеффри (2000). Брачный разум: как сексуальный выбор повлиял на эволюцию человеческой природы (1-е изд.). Нью-Йорк: Даблдей . ISBN 978-0-385-49516-5 . LCCN 00022673 . OCLC 43648482 .
- Митчелл, Мелани (1996). Введение в генетические алгоритмы . Сложные адаптивные системы. Кембридж, Массачусетс: MIT Press . ISBN 978-0-262-13316-6 . LCCN 95024489 . OCLC 42854439 .
- Пинкер, Стивен (1995) [Первоначально опубликовано в 1994 году; Нью-Йорк: Уильям Морроу и компания ]. Языковой инстинкт: как разум создает язык (1-е издание Harper Perennial ). Нью-Йорк: Харпер Многолетник. ISBN 978-0-06-097651-4 . LCCN 94039138 . OCLC 670524593 .
- Райс, Шон Х. (2004). Эволюционная теория: математические и концептуальные основы . Сандерленд, Массачусетс: Sinauer Associates. ISBN 978-0-87893-702-8 . LCCN 2004008054 . OCLC 54988554 .
- Ру, Вильгельм (1881). Борьба частей в организме . Лейпциг: Вильгельм Энгельманн . OCLC 8200805 . Борьба частей организма в Интернет-архиве . Проверено 11 августа 2015 г.
- Трезвый, Эллиотт (1993) [Первоначально опубликовано в 1984 году; Кембридж, Массачусетс: MIT Press]. Природа отбора: эволюционная теория в философском фокусе . Чикаго, Иллинойс: Издательство Чикагского университета . ISBN 978-0-226-76748-2 . LCCN 93010367 . OCLC 896826726 .
- Уоллес, Альфред Рассел (1871 г.) [первоначально опубликовано в 1870 г.]. Вклад в теорию естественного отбора. Серия очерков (2-е, с исправлениями и дополнениями под ред.). Нью-Йорк: Macmillan & Co. LCCN agr04000394 . OCLC 809350209 .
- Уильямс, Джордж К. (1966). Адаптация и естественный отбор: критика некоторых современных эволюционных идей . Принстонская научная библиотека. Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0-691-02615-2 . LCCN 65017164 . ОСЛК 35230452 .
- Уилсон, Дэвид Слоан (2002). Дарвиновский собор: эволюция, религия и природа общества . Чикаго, Иллинойс: Издательство Чикагского университета. ISBN 978-0-691-02615-2 . LCCN 2002017375 . OCLC 48777441 .
- Циммер, Карл ; Эмлен, Дуглас Дж. (2013). Эволюция: обретение смысла жизни (1-е изд.). Гринвуд-Виллидж, Колорадо: Издатели Roberts and Company. ISBN 978-1-936221-17-2 . LCCN 2012025118 . OCLC 767565909 .
Дальнейшее чтение
- Для технической аудитории
- Белл, Грэм (2008). Выбор: Механизм эволюции (2-е изд.). Оксфорд; Нью-Йорк: Издательство Оксфордского университета . ISBN 978-0-19-856972-5 . LCCN 2007039692 . OCLC 170034792 .
- Джонсон, Клиффорд (1976). Введение в естественный отбор . Балтимор, Мэриленд: University Park Press. ISBN 978-0-8391-0936-5 . LCCN 76008175 . ОСЛК 2091640 .
- Гулд, Стивен Джей (2002). Структура эволюционной теории . Кембридж, Массачусетс: Belknap Press издательства Гарвардского университета . ISBN 978-0-674-00613-3 . LCCN 2001043556 . OCLC 47869352 .
- Мейнард Смит, Джон (1993) [Первоначально опубликовано в 1958 году; Хармондсворт, Англия: Penguin Books ]. Теория эволюции (изд. Canto). Кембридж, Нью-Йорк: Издательство Кембриджского университета . ISBN 978-0-521-45128-4 . LCCN 93020358 . OCLC 27676642 .
- Поппер, Карл (декабрь 1978 г.). «Естественный отбор и возникновение разума» . Диалектика . 32 (3–4): 339–355. дои : 10.1111/j.1746-8361.1978.tb01321.x . ISSN 0012-2017 .
- Саммут-Бонничи, Таня; Уэнсли, Робин (сентябрь 2002 г.). «Дарвинизм, вероятность и сложность: рыночные организационные трансформации и изменения, объясненные теориями эволюции» (PDF) . Международный журнал обзоров менеджмента . 4 (3): 291–315. дои : 10.1111/1468-2370.00088 . ISSN 1460-8545 .
- Трезвый, Эллиотт , изд. (1994). Концептуальные проблемы эволюционной биологии (2-е изд.). Кембридж, Массачусетс: MIT Press . ISBN 978-0-262-69162-8 . LCCN 93008199 . ОСЛК 28150417 .
- Уильямс, Джордж К. (1992). Естественный отбор: области, уровни и проблемы . Оксфордская серия по экологии и эволюции. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-506933-4 . LCCN 91038938 . OCLC 228136567 .
- Для широкой аудитории
- Докинз, Ричард (1996). Восхождение на гору Невероятное (1-е американское изд.). Нью-Йорк: WW Norton & Company. ISBN 978-0-393-03930-6 . LCCN 34633422 . OCLC 34633422 .
- Гулд, Стивен Джей (1977). Со времен Дарвина: размышления о естествознании (1-е изд.). Нью-Йорк: WW Norton & Company. ISBN 978-0-393-06425-4 . LCCN 77022504 . ОСЛК 3090189 .
- Джонс, Стив (2000). Призрак Дарвина: Обновленное происхождение видов (1-е изд.). Нью-Йорк: Рэндом Хаус . ISBN 978-0-375-50103-6 . LCCN 99053246 . OCLC 42690131 .
- Левонтин, Ричард К. (сентябрь 1978 г.). "Приспособление". Научный американец . 239 (3): 212–230. Бибкод : 1978SciAm.239c.212L . doi : 10.1038/scientificamerican0978-212 . ISSN 0036-8733 . ПМИД 705323 .
- Майр, Эрнст (2002) [Первоначально опубликовано в 2001 году; Нью-Йорк: Основные книги ]. Что такое эволюция . Магистр наук. Лондон: Вайденфельд и Николсон . ISBN 978-0-297-60741-0 . LCCN 2001036562 . ОСЛК 248107061 .
- Вайнер, Джонатан (1994). Клюв зяблика: история эволюции в наше время (1-е изд.). Нью-Йорк: Кнопф . ISBN 978-0-679-40003-5 . LCCN 93036755 . ОСЛК 29029572 .
- Исторический
- Кон, Марек (2004). Причина всего: естественный отбор и английское воображение . Лондон: Фабер и Фабер . ISBN 978-0-571-22392-3 . LCCN 2005360890 . OCLC 57200626 .
- Зиркль, Конвей (25 апреля 1941 г.). «Естественный отбор до возникновения видов ». Труды Американского философского общества . 84 (1): 71–123. ISSN 0003-049X . JSTOR 984852 .
Внешние ссылки
- Дарвин, Чарльз . «О происхождении видов» . Архивировано из оригинала 25 февраля 2001 года. - Глава 4, Естественный отбор.