E. coli Эксперимент по долгосрочной эволюции

E. coli Эксперимент по долгосрочной эволюции ( LTEE ) — это продолжающееся исследование экспериментальной эволюции, начатое Ричардом Ленски в Калифорнийском университете в Ирвайне и продолженное Ленски и его коллегами из Университета штата Мичиган . [2] и в настоящее время курируется Джеффри Э. Барриком в Техасском университете в Остине . [3] С 24 февраля 1988 года он отслеживает генетические изменения в 12 первоначально идентичных популяциях бесполых бактерий Escherichia coli . [4] Ленски выполнил 10-тысячный перенос эксперимента 13 марта 2017 года. [5] В начале 2020 года популяция достигла более 73 000 поколений, незадолго до того, как была заморожена из-за пандемии COVID-19. [6] В сентябре 2020 года эксперимент LTEE был возобновлен с использованием замороженных запасов. [7]
В ходе эксперимента Ленски и его коллеги сообщили о широком спектре фенотипических и генотипических изменений в развивающихся популяциях. К ним относятся изменения, произошедшие во всех 12 популяциях, а также изменения, произошедшие только в одной или нескольких популяциях. Например, все 12 популяций продемонстрировали аналогичную картину быстрого улучшения физической формы, которое со временем замедлялось, более высоких темпов роста и увеличения размера клеток. У половины популяций развились дефекты репарации ДНК, которые вызвали мутаторные фенотипы, отмеченные повышенной частотой мутаций. Самая поразительная адаптация, о которой сообщалось до сих пор, — это эволюция аэробного роста на цитрате , что необычно для E. coli , в одной популяции в какой-то момент между 31 000 и 31 500 поколениями. [8] [9]
4 мая 2020 года Ленски объявил о продлении на 5 лет гранта в рамках Национального научного фонда , которая поддерживает LTEE. программы долгосрочных исследований в области биологии окружающей среды (LTREB) [10] Он также объявил, что доктор Джеффри Э. Баррик, доцент кафедры молекулярных биологических наук Техасского университета в Остине, возьмет на себя руководство экспериментом в течение 5-летнего периода финансирования. [11] Время эксперимента в Университете штата Мичиган закончилось в мае 2022 года, когда популяция достигла 75 000 поколений, и эксперимент был перенесен в лабораторию Баррика. [12] [ не удалось пройти проверку ] Эксперимент был возобновлен и возобновлен в лаборатории Баррика 21 июня 2022 года. [3]
Экспериментальный подход [ править ]
Эксперимент по долгосрочной эволюции был задуман как неограниченное средство эмпирического изучения центральных особенностей эволюции . Эксперимент был начат с тремя основными целями:
- Изучить динамику эволюции, включая скорость эволюционных изменений.
- Исследовать повторяемость эволюции.
- Чтобы лучше понять взаимосвязь между изменениями на фенотипическом и генотипическом уровнях. [13]
По мере продолжения эксперимента его масштабы расширялись по мере возникновения новых вопросов в эволюционной биологии, для решения которых его можно использовать, поскольку эволюция популяций представила новые явления для изучения, а также по мере развития технологий и методологических методов. [14]
Использование E. coli в качестве экспериментального организма позволило изучить многие поколения и большие популяции за относительно короткий период времени. Более того, благодаря длительному использованию E. coli в качестве основного модельного организма в молекулярной биологии , стал доступен широкий спектр инструментов, протоколов и процедур для изучения изменений на генетическом, фенотипическом и физиологическом уровнях. [15] Бактерии также можно заморозить и сохранить, оставаясь жизнеспособными. Это позволило создать то, что Ленски называет «замороженной летописью окаменелостей» образцов развивающихся популяций, которые можно возродить в любое время. Эта замороженная летопись окаменелостей позволяет возобновить популяцию в случае заражения или другого сбоя в эксперименте, а также позволяет изолировать и сравнивать живые экземпляры предковых и эволюционировавших клонов. Ленски выбрал штамм E. coli , который размножается только бесполым путем , лишен каких-либо плазмид , которые могли бы способствовать бактериальной конъюгации , и не имеет жизнеспособного профага . Как следствие, эволюция в эксперименте происходит только за счет основных эволюционных процессов мутации , генетического дрейфа и естественного отбора . Эта строгая асексуальность также означает, что генетические маркеры сохраняются в линиях и кладах общего происхождения , но не могут иным образом распространяться в популяциях. [13]
Ленски решил провести эксперимент с бактериями, выращенными в среде с ограниченным содержанием глюкозы под названием DM25. [16] который основан на минимальной среде, разработанной Бернардом Дэвисом для использования при выделении ауксотрофных мутантов E. coli с использованием пенициллина в качестве селективного агента. [17] [18] Для создания DM25 в минимальную среду добавляют глюкозу в низкой концентрации (25 мг/л). [16] Ленски выбрал эту концентрацию, чтобы упростить анализ эволюции популяций за счет уменьшения клональной интерференции , при которой в развивающейся популяции конкурируют несколько версий аллелей , а также уменьшения возможности эволюции экологических взаимодействий. [13] Эта концентрация используемой глюкозы поддерживает максимальную популяцию предков в 500 миллионов клеток в культуре объемом 10 мл, хотя теперь максимум варьируется среди развившихся популяций. [17] DM25 также содержит большое количество цитрата (примерно в 11 раз больше концентрации глюкозы), который первоначально был включен Дэвисом, поскольку он улучшал эффективность уничтожения пенициллина во время его экспериментов, хотя теперь известно, что он помогает в кишечной палочки приобретении . железа из среды. [17] [19]
Методы [ править ]
Этот раздел необходимо обновить . ( июнь 2023 г. ) |
12 популяций содержатся в инкубаторе с температурой 37 °C (99 °F) в лаборатории Ленски в Университете штата Мичиган . Каждый день 1% каждой популяции переносят в колбу со свежей питательной средой DM25. Разбавление означает, что в каждой популяции ежедневно появляется 6,64 поколения или удвоение численности. Большие репрезентативные образцы каждой популяции замораживают с использованием глицерина в качестве криопротектора с интервалом в 500 поколений (75 дней). Бактерии в этих образцах остаются жизнеспособными и могут быть возрождены в любое время. Эта коллекция образцов называется «замороженной летописью окаменелостей» и представляет собой историю эволюции каждой популяции на протяжении всего эксперимента. Популяции также регулярно проверяются на предмет изменений средней приспособленности , и регулярно проводятся дополнительные эксперименты для изучения интересных изменений в популяциях. [20] По состоянию на апрель 2016 г. [update]Популяции E. coli изучались на протяжении более 64 500 поколений, и считается, что они претерпели достаточно спонтанных мутаций , так что каждая возможная единичная точечная мутация в геноме E. coli произошла несколько раз. [8]
Штамм-основатель [ править ]
Штамм E. coli, который Ленски решил использовать в эксперименте по долгосрочной эволюции, был получен из «штамма B», как описано в статье Сеймура Ледерберга 1966 года (в которой этот штамм неправильно идентифицирован как «Bc251», хотя более поздний генетический анализ обнаружил вместо этого будет «B») через Брюса Левина, который использовал его в эксперименте по бактериальной экологии в 1972 году. Определяющими генетическими признаками этого штамма были: T6. р , ул. р , р − м − , Мы покупаем − (неспособен расти на арабинозе ). [4] Ленски обозначил исходный штамм-основатель как REL606. Перед началом эксперимента Ленский выделил Ара. + вариант штамма, в котором мутация оперона ara точечная восстановила рост арабинозы, который он обозначил как штамм REL607. Начиная долгосрочный эволюционный эксперимент, Ленски основал шесть популяций с шестью отдельными Ара. − колонии REL606. Эти популяции называются от Ara-1 до Ara-6. Ленский также основал еще шесть популяций из шести отдельных Ара. + колонии REL607. Их называют популяциями от Ara+1 до Ara+6. Маркерные различия позволяют дифференцировать штаммы на чашках с тетразолием арабинозой, на которых Ara − колонии кажутся красными, а Ара + колонии выглядят от белого до розового цвета. В ходе эксперимента каждая популяция накопила большое количество различных мутаций, что позволяет в дальнейшем идентифицировать штаммы по их популяции происхождения. [ нужна ссылка ]
Результаты [ править ]
Изменения в фитнесе [ править ]
Значительная часть анализа эксперимента была посвящена тому, как изменилась приспособленность популяций к их предковой линии. Во всех популяциях наблюдалась тенденция быстрого увеличения относительной приспособленности в течение первых поколений, причем этот рост со временем замедлялся. К 20 000 поколениям популяция росла примерно на 70% быстрее, чем предковая линия. [13] Этот рост и замедление роста продолжались и в последующих поколениях. Исследование Wiser et al., проведенное в 2013 году. сообщили о продолжающемся улучшении на протяжении 50 000 поколений по сравнению с образцами, изолированными на протяжении 40 000 поколений. Они обнаружили, что увеличение приспособленности гораздо лучше соответствует степенной модели, чем гиперболическим моделям, которые использовались ранее. Поскольку модель степенного закона описывает постоянно замедляющийся рост, не имеющий верхнего предела, в то время как гиперболическая модель предполагает жесткий предел, в работе предполагается, что рост будет продолжаться без ограничений, поскольку в популяциях фиксируются мутации с постепенно уменьшающейся пользой. [22] В дальнейшей работе, опубликованной в 2015 году, были представлены результаты более 1100 новых тестов физической формы, в которых изучались изменения физической формы на протяжении 60 000 поколений. Данные снова соответствуют предложенной степенной модели и, действительно, соответствуют предсказаниям модели на основе более ранних данных. Эти результаты показывают, что, вопреки предыдущему мнению, адаптация и адаптивная дивергенция потенциально могут увеличиваться бесконечно, даже в постоянной среде. [23] [24] [25]
Эволюция генома [ править ]
Сообщается, что из 12 популяций у шести развились дефекты в способности восстанавливать ДНК , что значительно увеличивает скорость мутаций в этих штаммах. [9] [26] [27] Хотя считается, что бактерии в каждой популяции произвели сотни миллионов мутаций в течение первых 20 000 поколений, Ленски подсчитал, что за этот период времени только от 10 до 20 полезных мутаций достигли фиксации в каждой популяции, при этом общее количество точковых мутаций составляет менее 100. (включая нейтральные мутации ), достигающие фиксации в каждой популяции. [13] В 2009 году Баррик и др. сообщили о результатах изучения последовательностей генома из нескольких моментов времени в популяции Ara-1. Они обнаружили, что, в отличие от снижения темпов улучшения физической формы, накопление мутаций было линейным и часовым, хотя некоторые данные свидетельствуют о том, что большая часть накопления была полезной, а не нейтральной. [28]
Эволюция увеличения размера клеток во двенадцати всех популяциях
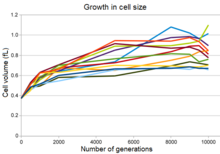
Все двенадцать экспериментальных популяций демонстрируют увеличение размера клеток одновременно со снижением максимальной плотности популяции, а во многих популяциях - более округлую форму клеток. [29] Частично это изменение было результатом мутации, которая изменила экспрессию гена пенициллин -связывающего белка , что позволило мутантным бактериям вытеснить предковые бактерии в условиях долгосрочного эволюционного эксперимента. Однако, хотя эта мутация повысила приспособленность бактерий в этих условиях, она также увеличила чувствительность бактерий к осмотическому стрессу и снизила их способность выживать в течение длительного времени в культурах стационарной фазы. [29]
Экологическая специализация [ править ]
В ходе эксперимента популяции эволюционировали и стали специализироваться на ресурсе глюкозы, на котором они растут. Впервые это было описано в 2000 году, когда Купер и Ленски продемонстрировали, что все популяции испытали распад неиспользованных метаболических функций после 20 000 поколений, что ограничивает диапазон веществ, на которых могут расти бактерии. Их анализ показал, что этот распад произошел из-за антагонистической плейотропии , при которой мутации, улучшающие способность к росту на глюкозе, снижали или устраняли способность к росту на других веществах. [30] Более позднее исследование Лейби и Маркса, в котором использовались более продвинутые методы, показало, что большая часть распадов, выявленных Купером и Ленски, была экспериментальными артефактами, что потеря неиспользуемых функций была не такой обширной, как предполагалось на первый взгляд, и что некоторые неиспользуемые функции улучшились. Более того, они пришли к выводу, что метаболические потери были вызваны не антагонистической плейотропией, а нейтральным накоплением мутаций в неиспользуемых частях генома, предполагая, что адаптация к простой среде не обязательно может привести к специализации. [31]
сбалансированного полиморфизма и Эволюция экосистем простых
Два различных варианта, S и L, были идентифицированы в популяции, обозначенной как Ara-2, в 18 000 поколениях на основании формирования ими малых и больших колоний соответственно. [32] Клоны типов S и L могли стабильно сосуществовать в совместной культуре друг с другом, что указывает на то, что они занимали отдельные ниши в популяции. Это было подтверждено обнаружением того, что тип L имел преимущество во время роста на глюкозе, но что тип S имел преимущество во время стационарной фазы, после того как глюкоза закончилась. Было обнаружено, что эти два типа первоначально развились до 6000 поколений, а затем сосуществовали после этого. [32] Филогенетический анализ клонов двух типов, выделенных из разных поколений, показал, что типы S и L принадлежали к разным, сосуществующим линиям в популяции и, возможно, подвергались зачаточному видообразованию. [33]
Эволюция использования аэробного цитрата популяции одной в
Предыстория [ править ]

E. coli расти обычно не способна аэробно на цитрате из-за неспособности экспрессировать переносчик цитрата в присутствии кислорода. [35] Однако E. coli имеет полный цикл лимонной кислоты и поэтому метаболизирует цитрат в качестве промежуточного продукта во время аэробного роста на другие вещества, включая глюкозу. Большинство E. coli могут расти анаэробно на цитрате посредством ферментации , если доступен ко-субстрат, такой как глюкоза, обеспечивающий восстанавливающую способность. [8] [35] [36] [37] Анаэробный citT рост возможен благодаря экспрессии трансмембранного гена цитрат-сукцинатного антипортера, , который был впервые идентифицирован в 1998 году. Этот ген регулируется совместно с другими генами, участвующими в ферментации цитрата, обнаруженными в опероне cit , который включается. только при отсутствии кислорода. [35] [38]
Неспособность аэробно расти на цитрате, называемом Cit. − фенотип считается определяющей характеристикой E. coli как вида и является ценным средством дифференциации E. coli от патогенной сальмонеллы . Хотя цит. + штаммы E. coli были выделены из образцов окружающей среды и сельскохозяйственных культур, в каждом таком случае было обнаружено, что этот признак обусловлен наличием плазмиды, несущей чужеродный переносчик цитрата. [39] Одиночный, спонтанный Cit + О мутанте E. coli сообщил Холл в 1982 году. [40] Этот мутант был выделен в ходе длительной селекции на рост на другом новом веществе в питательной среде, которая также содержала цитрат. Генетический анализ Холла показал, что основная мутация была сложной, но в конечном итоге он не смог определить точные изменения или задействованные гены, что привело его к гипотезе об активации загадочного гена-переносчика. [40] Области генома, к которым Холлу удалось сузить расположение изменений, не соответствуют известному местоположению гена citT , выявленному 16 лет спустя, равно как и физиологические характеристики в транспортных анализах Холла Cit. + мутанты соответствуют ожидаемым в отношении аэробной экспрессии транспортера CitT. [39] [41]
Цит + развивается в LTEE [ править ]
В 2008 году команда Ленски под руководством Закари Д. Блаунта сообщила, что способность к аэробному росту на цитрате развилась у одной популяции. Примерно в поколении 33 127 резкое увеличение мутности наблюдалось в популяции, обозначенной как Ара-3. Они обнаружили, что популяция содержит клоны, способные аэробно расти на цитрате (Cit. + ). Эта метаболическая способность позволила популяции вырасти в несколько раз больше, чем раньше, из-за большого количества цитрата, присутствующего в среде. Исследование замороженных образцов ископаемых популяций показало, что Cit + клоны могут быть изолированы уже в 31 500 поколениях. Город + Было обнаружено, что варианты в популяции обладают рядом генетических маркеров, уникальных для популяции Ara-3; это наблюдение исключило возможность того, что клоны были контаминантами, а не спонтанными мутантами. В серии экспериментов, «переигравших» ленту эволюции Ара-3 от Цита − клонов, изолированных из образцов, замороженных в различные моменты истории популяции, они продемонстрировали, что способность аэробного роста на цитрате с большей вероятностью будет повторно развиваться в подмножестве генетически чистых, эволюционировавших клонов. В этих экспериментах они наблюдали 19 новых независимых экземпляров Cit. + реэволюция, но только если начинать с клонов, изолированных после 20 000 поколения. Флуктуационные тесты показали, что клоны этого поколения и более поздние демонстрируют скорость мутации Cit. + признак, который был значительно выше, чем предковый уровень. Даже в этих более поздних клонах скорость мутации Cit + было порядка одного случая на триллион клеточных делений. [8]
Ленски и его коллеги пришли к выводу, что эволюция Cit + Функция в этой популяции возникла благодаря одной или нескольким более ранним, возможно, неадаптивным, «потенцирующим» мутациям, которые увеличили скорость мутаций до доступного уровня. Данные свидетельствуют о том, что использование цитрата вызвало как минимум две мутации, последовавшие за этими «потенциирующими» мутациями. В более общем плане авторы предполагают, что эти результаты, следуя аргументу Стивена Джея Гулда , указывают на то, что «историческая случайность может иметь глубокое и продолжительное влияние» на ход эволюции. [8] Эти открытия стали считаться важным примером влияния исторических обстоятельств на эволюцию . [17] [42] [43]
Геномный анализ Cit + черта и последствия эволюционных для инноваций
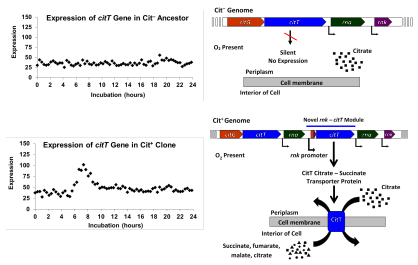
В 2012 году Ленски и его команда сообщили о результатах геномного анализа Cit. + черта, которая проливает свет на генетическую основу и историю эволюции этой черты. Исследователи секвенировали полные геномы двадцати девяти клонов, выделенных в разные моменты истории популяции Ара-3. Они использовали эти последовательности для реконструкции филогенетической истории популяции; эта реконструкция показала, что популяция разделилась на три клады за 20 000 поколений. Город + варианты развились в одной из них, которую они назвали кладой 3. Клоны, потенцирование которых было обнаружено в более ранних исследованиях, были распределены по всем трем кладам, но были чрезмерно представлены в кладе 3. Это привело исследователей к выводу, что было как минимум две потенцирующие мутации, участвующие в Cit + эволюция. [9]
Исследователи также обнаружили, что все Cit + клоны имели мутации, при которых сегмент ДНК из 2933 пар оснований дублировался или амплифицировался. Дублированный сегмент содержал ген citT белка-переносчика цитрата, используемого при анаэробном росте на цитрате. Дублирование происходит тандемно, в результате чего копии располагаются лицом к хвосту по отношению друг к другу. Эта новая конфигурация помещает копию ранее молчащего, неэкспрессируемого citT под контроль промотора соседнего гена rnk , который управляет экспрессией в присутствии кислорода. Этот новый модуль rnk-citT создал новый механизм регуляции citT , активируя экспрессию переносчика цитрата при наличии кислорода и тем самым обеспечивая аэробный рост цитрата. [9]
Перемещение этого модуля rnk-citT в геном потенцированного Cit. − Было показано, что клона достаточно для производства Cit + фенотип. Однако первоначальный Cit + фенотип, полученный в результате дупликации, был очень слабым и давал улучшение приспособленности только на ~ 1%. Исследователи обнаружили, что количество копий модуля rnk-citT необходимо увеличить, чтобы усилить Cit. + Этого признака достаточно, чтобы позволить бактериям хорошо расти на цитрате. Дальнейшие мутации после Cit + бактерии стали доминировать в популяции, продолжали накапливать улучшенный рост на цитрате. [ нужна ссылка ]
Исследователи пришли к выводу, что эволюция Cit + признак возник в три отдельные фазы: (1) накопление мутаций, которые увеличили скорость мутации в Cit + , (2) сам признак проявился в слабой форме и (3) признак был улучшен за счет более поздних мутаций. Блаунт и др. предположил, что эта закономерность может быть типичной для развития новых черт в целом, и предложил трехэтапную модель эволюционных инноваций:
- Потенцирование : развивается генетический фон, в котором признак становится доступным для мутации, что делает возможной эволюцию признака.
- Актуализация : происходит мутация, которая производит признак, проявляя его, хотя, вероятно, в слабой форме.
- Уточнение : как только признак существует, если он обеспечивает селективную выгоду, будут накапливаться мутации, которые улучшат признак, делая его эффективным. Эта фаза бессрочна и будет продолжаться до тех пор, пока возникают уточняющие мутации и признак остается полезным. [9] [17]
Эта модель нашла признание в эволюционной биологии. В 2015 году палеонтолог Дуглас Эрвин предложил модификацию четырехэтапной модели, чтобы лучше отразить возможное различие между эволюционной новизной и эволюционными инновациями, а также подчеркнуть важность условий окружающей среды: потенциация, генерация новых фенотипов (актуализация), адаптивное усовершенствование и эксплуатация (превращение новинки в инновацию, поскольку она становится важной для экологического истеблишмента обладающих организмов). [45]
Исследование потенциации [ править ]
В 2014 году исследовательская группа под руководством Эрика Квандта в лаборатории Джеффри Баррика в Техасском университете в Остине описала применение нового метода под названием «Рекурсивная полногеномная рекомбинация и секвенирование» (REGRES) для выявления потенцирующих мутаций среди 70 присутствующих в Ara. -3 линии, которые развили Цит + . [46] В этом методе использовалось несколько этапов процесса, в котором F-плазмиды на основе конъюгация между Cit из 33 000 поколений + клон, CZB154 и Cit − основатель клона LTEE для очистки мутаций, не необходимых ни для проявления слабой, ни для сильной формы Cit + черта, последняя называется Cit ++ . Они обнаружили, что модуль rnk-citT, ответственный за фенотипическое переключение на Cit + было достаточно, чтобы создать слабый Cit + фенотип у предка. Они также идентифицировали мутацию, которая произошла в линии, ведущей к CZB154, после первоначальной эволюции Cit. + что дало сильный Cit ++ фенотипе у предка отсутствовала какая-либо мутация, кроме модуля rnk-citT . Эта мутация, обнаруженная в регуляторной области гена dctA , вызвала значительное увеличение экспрессии транспортера DctA , который импортирует C 4 -дикарбоксилаты в клетку. Они обнаружили, что повышенная экспрессия DctA позволила Cit + клетки для повторного поглощения сукцината , малата и фумарата, высвобождаемых в среду транспортером CitT во время импорта цитрата. Они выявили аналогичную мутацию у Cit ++ клоны в популяции Ara-3, которые увеличили экспрессию DctA за счет восстановления функции регулирующего ее гена, dcuS , который был деактивирован в предковом клоне. Квандт и др. пришли к выводу, что мутация dctA участвует не в потенцировании, а в усовершенствовании. Это привело их к предположению, что эволюция Cit + в популяции Ara-3 могло зависеть от генетического фона и специфичной для популяции экологии, которые позволили ранним, слабым Cit + варианты сохраняются в популяции достаточно долго, чтобы возникли уточняющие мутации и сделали рост на цитрате достаточно сильным, чтобы обеспечить значительную пользу для фитнеса. [ нужна ссылка ]
Позже Квандт и его коллеги опубликовали результаты, окончательно идентифицирующие мутацию, которая действительно усиливала Cit. + эволюция. [47] Эта мутация произошла в гене gltA , который кодирует цитратсинтазу , фермент, участвующий в поступлении углерода в цикл лимонной кислоты . Он имел эффект увеличения активности цитратсинтазы, и они показали, что это способствует улучшению роста на ацетате . Более того, при gltA мутации модуль rnk-citT , вызывающий Cit + Черта оказывает нейтральный или слегка полезный эффект на приспособленность, в то время как без нее модуль был сильно вреден. Таким образом, мутация gltA , по-видимому, позволила ранним, слабым Cit + варианты сохраняются в популяции до тех пор, пока не возникнут более поздние уточняющие мутации, что согласуется с их более ранними выводами. После сильного цитирования ++ фенотип эволюционировал, повышенная активность цитратсинтазы стала вредной. Исследователи обнаружили, что более поздние мутации gltA противодействуют первой мутации, снижая активность цитратсинтазы и еще больше улучшая рост цитрата. Они пришли к выводу, что серия мутаций gltA сначала усиливает, а затем улучшает рост цитрата. Они также предположили, что линия, в которой находится Cit + возник, возможно, занял нишу в Ara-3 в результате роста на ацетате, и что потенцирующие мутации, которые привели к эволюции Cit + в Ara-3 изначально были адаптированы к использованию ацетата. [ нужна ссылка ]
Расследование постцита + экология и устойчивое разнообразие [ править ]
Небольшая субпопуляция Cit − клетки, не способные расти на цитрате и принадлежащие к отдельной кладе, персистировали в популяции после Cit + клетки стали доминантными. Ранние результаты показали, что это разнообразие было частично связано с Cit − клетки лучше растут на глюкозе в среде. [8] Тернер и др. позже выяснилось, что еще одним фактором сосуществования было то, что Cit − клетки развили способность перекрестно питаться Cit + большинство. Они показали, что Цит. + клетки выделяют сукцинат , малат и фумарат во время роста на цитрате, поскольку транспортер CitT выкачивает эти вещества из клетки, одновременно перекачивая цитрат в клетку. Город − клетки быстро развили способность расти на этих веществах благодаря мутации, которая восстановила экспрессию соответствующего белка-переносчика, который молчал у предка. [48]
Город − субпопуляция в конечном итоге вымерла в популяции между 43 500 и 44 000 поколениями. Было показано, что это вымирание произошло не из-за Cit. + большинство развивается, чтобы иметь возможность вторгнуться в нишу, занимаемую Cit − меньшинство. Действительно, цит. − клоны могут вторгнуться в Цит + популяций после вымирания. Более того, в эксперименте, в котором они перезапустили двадцать повторений популяции Ara-3 из образца, замороженного за 500 поколений до вымирания, Turner et al. обнаружил, что Цит. − субпопуляция не вымерла ни в одной из повторностей после 500 поколений эволюции. Одна из этих реплик продолжалась на протяжении 2500 поколений, в течение которых Цит. − продолжали сосуществовать. Исследователи пришли к выводу, что исчезновение Cit − произошло из-за какого-то неизвестного «редкого возмущения окружающей среды», подобного тому, которое может повлиять на природные популяции. [49] Последний экземпляр был интегрирован в основной эксперимент LTEE, став тринадцатой популяцией, Ara-7. [50]
Различные интерпретации результатов [ править ]
Другие исследователи экспериментировали с развитием аэробной цитратной кишечной палочки . Дастин Ван Хофвеген и др. смогли изолировать 46 независимых мутантов E. coli , использующих цитрат , всего за 12–100 поколений, используя длительную селекцию в условиях голодания, в ходе которой бактерии быстрее получали больше мутаций. [51] В их исследовании секвенирование геномной ДНК выявило амплификацию локусов citT и dctA , а реаранжировка ДНК представляла собой тот же класс мутаций, которые были выявлены в эксперименте Ричарда Ленски и его команды. Они пришли к выводу, что редкость мутанта, использующего цитрат, в исследовании Ленски, скорее всего, была результатом выборочных экспериментальных условий, использованных его командой, а не уникальным событием эволюционного видообразования. [51]
Джон Рот и Софи Мэнье-Патен рассмотрели подходы как к отсроченным мутациям группы Ленски, так и к быстрым мутациям группы Ван Хофвегена в E. coli . Они утверждают, что обе команды наблюдали одну и ту же последовательность усиления, актуализации и совершенствования, ведущую к одинаковым Cit. + варианты. [52] По их словам, период Ленски продолжительностью менее суток, в течение которого использование цитрата будет подвергаться отбору, за которым последует 100-кратное разведение, и период роста на глюкозе, который не будет отбирать использование цитрата; в конечном итоге снизила вероятность того, что E. coli сможет накапливать ранние адаптивные мутации от одного периода селекции к другому. [52] Напротив, команда Ван Хофвегена предусмотрела непрерывный период отбора в 7 дней, что привело к более быстрому развитию E. coli , использующей цитрат . Рот и Менье-Патен предполагают, что серийное разведение E. coli и короткий период отбора на использование цитрата в условиях LTEE постоянно препятствовали достижению каждым поколением E. coli следующих стадий аэробного использования цитрата. [52]
Ленски утверждает, что проблема не в экспериментах или данных, а в интерпретациях, сделанных Ван Хофвегеном и др. и Менье-Патен и Рот. [53] По его словам, быстрая эволюция Cit + не обязательно было неожиданным, поскольку его команда также смогла создать множество Cit + мутантов за несколько недель во время повторных экспериментов. [54] Он утверждает, что LTEE не был разработан для изоляции мутантов, использующих цитрат, или для борьбы с видообразованием, которое является процессом, а не событием. Более того, он утверждал, что эволюция Cit + в LTEE зависело от мутаций, накопившихся ранее. [54]
См. также [ править ]
Ссылки [ править ]
- ^ Пенниси, Элизабет (14 ноября 2013 г.). «Человек, который закупорил эволюцию». Наука . 342 (6160): 790–793. Бибкод : 2013Sci...342..790P . дои : 10.1126/science.342.6160.790 . ПМИД 24233702 .
- ^ Jump up to: Перейти обратно: а б «Фотографии» . Лаборатория Блаунта . Проверено 28 мая 2016 г.
- ^ Jump up to: Перейти обратно: а б Баррик, Джеффри (21 июня 2022 г.). «Начало LTEE в UT в Остине» . Эксперимент долгосрочной эволюции . Проверено 22 июня 2022 г.
- ^ Jump up to: Перейти обратно: а б Ленски, Ричард Э. (2000). «Источник основополагающего штамма» . Домашняя страница Ричарда Э. Ленски . Мичиганский государственный университет. Архивировано из оригинала 31 мая 2018 г. Проверено 18 июня 2008 г.
- ^ «Некоторые морщины времени» . Возвращение к Теллиамеду . 13 марта 2017 г. Проверено 13 марта 2017 г.
- ^ Ленски, Ричард Э. (09 марта 2020 г.). «Мы прерываем этот ужасный вирус хорошими новостями о бактериях» . Возвращение к Теллиамеду . Проверено 27 мая 2021 г.
- ^ «Они вернулись!» . Возвращение к Теллиамеду . 22 сентября 2020 г. Проверено 17 июня 2021 г.
- ^ Jump up to: Перейти обратно: а б с д и ж Блаунт, Закари Д.; Борланд, Кристина З.; Ленски, Ричард Э. (2008). «Историческая случайность и эволюция ключевой инновации в экспериментальной популяции Escherichia coli » . Труды Национальной академии наук . 105 (23): 7899–906. Бибкод : 2008PNAS..105.7899B . дои : 10.1073/pnas.0803151105 . JSTOR 25462703 . ПМЦ 2430337 . ПМИД 18524956 .
- ^ Jump up to: Перейти обратно: а б с д и Блаунт З.Д., Баррик Дж.Э., Дэвидсон С.Дж., Ленски Р.Э. (27 сентября 2012 г.). «Геномный анализ ключевой инновации в экспериментальной популяции Escherichia coli » . Природа . 489 (7417): 513–518. Бибкод : 2012Natur.489..513B . дои : 10.1038/nature11514 . ПМЦ 3461117 . ПМИД 22992527 .
- ^ «Долгосрочные исследования в области биологии окружающей среды (LTREB)» . new.nsf.gov . Национальный научный фонд. 9 декабря 2020 г. Проверено 10 июня 2024 г.
- ^ «Еще пять лет» . Возвращение к Теллиамеду . 04.05.2020 . Проверено 9 мая 2020 г.
- ^ «Твиттер/Твиттер» . Твиттер.com . Проверено 21 июня 2022 г.
- ^ Jump up to: Перейти обратно: а б с д и Ленски, Ричард Э. (2010). «Фенотипическая и геномная эволюция в ходе эксперимента с 20 000 поколений бактерии Escherichia coli » . Обзоры селекции растений . стр. 225–265. дои : 10.1002/9780470650288.ch8 . ISBN 978-0-471-46892-9 . S2CID 82586203 .
- ^ Фокс, Джереми В.; Ленски, Ричард Э. (23 июня 2015 г.). «Отныне и в вечность — теория и практика действительно длительного эксперимента» . ПЛОС Биология . 13 (6): e1002185. дои : 10.1371/journal.pbio.1002185 . ПМЦ 4477892 . ПМИД 26102073 .
- ^ Блаунт, Закари Д. (25 марта 2015 г.). «Неисчерпаемый потенциал кишечной палочки» . электронная жизнь . 4 . doi : 10.7554/eLife.05826 . ПМЦ 4373459 . ПМИД 25807083 .
- ^ Jump up to: Перейти обратно: а б «Жидкая среда ДМ25» . Lenski.mmg.msu.edu . Проверено 6 мая 2022 г.
- ^ Jump up to: Перейти обратно: а б с д и Блаунт, Закари Д. (август 2016 г.). «Пример эволюционных непредвиденных обстоятельств» . Исследования по истории и философии науки. Часть C: Исследования по истории и философии биологических и биомедицинских наук . 58 : 82–92. дои : 10.1016/j.shpsc.2015.12.007 . ПМИД 26787098 .
- ^ Фицджеральд, Джордж; Уильямс, Лютер С. (апрель 1975 г.). «Модифицированная процедура обогащения пенициллином для отбора бактериальных мутантов» . Журнал бактериологии . 122 (1): 345–346. дои : 10.1128/JB.122.1.345-346.1975 . ПМК 235679 . ПМИД 1091629 .
- ^ Вагегг, В; Браун, В. (январь 1981 г.). «Транспорт цитрата железа в Escherichia coli требует белка рецептора внешней мембраны fecA» . Журнал бактериологии . 145 (1): 156–163. дои : 10.1128/JB.145.1.156-163.1981 . ПМК 217256 . ПМИД 7007312 .
- ^ Ленски, Ричард Э. (2000). «Обзор E. coli эксперимента по долгосрочной эволюции » . Домашняя страница Ричарда Э. Ленски . Мичиганский государственный университет. Архивировано из оригинала 28 августа 2008 г. Проверено 18 июня 2008 г.
- ^ Ленски, Ричард (2010). «Обзор E. coli эксперимента по долгосрочной эволюции » . myxo.css.msu.edu . Архивировано из оригинала 14 ноября 2021 г. Проверено 29 июня 2023 г.
- ^ Уайзер, MJ; Рибек, Н.; Ленский, Р.Э. (14 ноября 2013 г.). «Многолетняя динамика адаптации асексуальных популяций» . Наука . 342 (6164): 1364–1367. Бибкод : 2013Sci...342.1364W . дои : 10.1126/science.1243357 . ПМИД 24231808 . S2CID 15341707 .
- ^ Шарпинг, Натаниэль (16 декабря 2015 г.). «Может ли эволюция когда-либо создать «идеальный» организм?» . Откройте для себя журнал . Архивировано из оригинала 20 декабря 2015 года . Проверено 18 декабря 2015 г.
- ^ Ленски, Ричард Э; и др. (2015). «Устойчивый рост физической формы и изменчивость траекторий физической формы в долгосрочном эксперименте по эволюции Escherichia coli » . Труды Королевского общества B: Биологические науки . 282 (1821): 20152292. doi : 10.1098/rspb.2015.2292 . ПМЦ 4707762 . ПМИД 26674951 .
- ^ Казначеев, Артем (май 2019). «Вычислительная сложность как окончательное ограничение эволюции» . Генетика . 212 (1): 245–265. doi : 10.1534/genetics.119.302000 . ПМК 6499524 . ПМИД 30833289 .
- ^ Сниговский, Пол Д.; Джерриш, Филип Дж.; Ленски, Ричард Э. (июнь 1997 г.). «Эволюция высоких частот мутаций в экспериментальных популяциях кишечной палочки» . Природа . 387 (6634): 703–705. Бибкод : 1997Natur.387..703S . дои : 10.1038/42701 . ПМИД 9192894 . S2CID 4351382 .
- ^ Баррик, Дж. Э.; Ленски, Р.Э. (23 сентября 2009 г.). «Общегеномное мутационное разнообразие в развивающейся популяции Escherichia coli» . Симпозиумы Колд-Спринг-Харбор по количественной биологии . 74 : 119–129. дои : 10.1101/sqb.2009.74.018 . ПМК 2890043 . ПМИД 19776167 .
- ^ Баррик, Джеффри Э.; Ю, Донг Су; Юн, Сон Хо; Чон, Хэён; О, Тэ Кван; Шнайдер, Доминик; Ленски, Ричард Э.; Ким, Джихён Ф. (18 октября 2009 г.). «Эволюция и адаптация генома в долгосрочном эксперименте с Escherichia coli». Природа . 461 (7268): 1243–1247. Бибкод : 2009Natur.461.1243B . дои : 10.1038/nature08480 . ПМИД 19838166 . S2CID 4330305 .
- ^ Jump up to: Перейти обратно: а б Филипп, Надеж; Пелоси, Людовик; Ленски, Ричард Э.; Шнайдер, Доминик (2008). «Эволюция концентрации пенициллинсвязывающего белка 2 и формы клеток в ходе длительного эксперимента с Escherichia coli » . Журнал бактериологии . 191 (3): 909–21. дои : 10.1128/JB.01419-08 . ПМК 2632098 . ПМИД 19047356 .
- ^ Купер, Вон С.; Ленски, Ричард Э. (октябрь 2000 г.). «Популяционная генетика экологической специализации в развитии популяций Escherichia coli». Природа . 407 (6805): 736–739. Бибкод : 2000Natur.407..736C . дои : 10.1038/35037572 . ПМИД 11048718 . S2CID 205009743 .
- ^ Лейби, Николас; Маркс, Кристофер Дж.; Моран, Нэнси А. (18 февраля 2014 г.). «Метаболическая эрозия, главным образом за счет накопления мутаций, а не компромиссов, приводит к ограниченной эволюции специфичности субстрата у Escherichia coli» . ПЛОС Биология . 12 (2): e1001789. дои : 10.1371/journal.pbio.1001789 . ПМЦ 3928024 . ПМИД 24558347 .
- ^ Jump up to: Перейти обратно: а б Розен, Дэниел Э.; Ленски, Ричард Э. (январь 2000 г.). «Долгосрочная экспериментальная эволюция Escherichia coli . VIII. Динамика сбалансированного полиморфизма». Американский натуралист . 155 (1): 24–35. дои : 10.1086/303299 . ПМИД 10657174 . S2CID 4440384 .
- ^ Розен, Дэниел Э.; Шнайдер, Доминик; Ленски, Ричард Э. (27 июня 2005 г.). «Долгосрочная экспериментальная эволюция Escherichia coli. XIII. Филогенетическая история сбалансированного полиморфизма». Журнал молекулярной эволюции . 61 (2): 171–180. Бибкод : 2005JMolE..61..171R . дои : 10.1007/s00239-004-0322-2 . ПМИД 15999245 . S2CID 6970967 .
- ^ «Об эволюции использования цитрата» . Возвращение к Теллиамеду . 20 февраля 2016 г. Проверено 26 мая 2016 г.
- ^ Jump up to: Перейти обратно: а б с «Клеточная биология: использование цитрата» . ЭВО-ЭД . Мичиганский университет.
- ^ Лара, ФДЖС; Стоукс, Дж. Л. (1952). «Окисление цитрата кишечной палочкой » . Журнал бактериологии . 63 (3): 415–420. дои : 10.1128/JB.63.3.415-420.1952 . ПМК 169284 . ПМИД 14927574 .
- ^ Лютгенс, М.; Готшалк, Г. (1 июля 1980 г.). «Почему для анаэробного роста Escherichia coli на цитрате необходим ко-субстрат» . Микробиология . 119 (1): 63–70. дои : 10.1099/00221287-119-1-63 . ПМИД 6997437 .
- ^ Пос, Клаас Мартинус; Димрот, Питер; Ботт, Майкл (август 1998 г.). « Носитель цитрата CitT Escherichia coli : член нового семейства эубактериальных транспортеров, родственный транслокатору 2-оксоглутарат/малат из хлоропластов шпината» . Журнал бактериологии . 180 (16): 4160–4165. дои : 10.1128/JB.180.16.4160-4165.1998 . ПМЦ 107412 . ПМИД 9696764 .
- ^ Jump up to: Перейти обратно: а б Рейнольдс, Швейцария; Сильвер, S (декабрь 1983 г.). «Утилизация цитрата Escherichia coli: системы, кодируемые плазмидами и хромосомами» . Журнал бактериологии . 156 (3): 1019–1024. дои : 10.1128/JB.156.3.1019-1024.1983 . ПМК 217945 . ПМИД 6358185 .
- ^ Jump up to: Перейти обратно: а б Холл, Б.Г. (июль 1982 г.). «Хромосомная мутация использования цитрата Escherichia coli K-12» . Журнал бактериологии . 151 (1): 269–273. дои : 10.1128/JB.151.1.269-273.1982 . ПМК 220237 . ПМИД 7045076 .
- ^ « Эшерихия coli К-12 субстр. MG1655 citT» . ecocyc.org . Проверено 23 мая 2016 г.
- ^ Дежарден, Эрик (1 января 2011 г.). «Историчность и экспериментальная эволюция» . Биология и философия . 26 (3): 339–364. дои : 10.1007/s10539-011-9256-4 . S2CID 83908986 .
- ^ Битти, Джон; Каррера, Изабель (1 января 2011 г.). «Когда то, что должно было случиться, не должно было произойти: история, шанс, повествование, эволюция» . Журнал философии истории . 5 (3): 471–495. дои : 10.1163/187226311x599916 .
- ^ «Фотографии» . Лаборатория Блаунта . Проверено 6 октября 2017 г.
- ^ Эрвин, Дуглас Х. (октябрь 2015 г.). «Новизна и инновации в истории жизни» . Современная биология . 25 (19): 930–940 р. дои : 10.1016/j.cub.2015.08.019 . ПМИД 26439356 .
- ^ Квандт, Эрик М.; Детередж, Дэниел Э.; Эллингтон, Эндрю Д.; Георгиу, Джордж; Баррик, Джеффри Э. (11 февраля 2014 г.). «Рекурсивная полногеномная рекомбинация и секвенирование открывают ключевой шаг в эволюции метаболических инноваций в Escherichia coli » . Труды Национальной академии наук . 111 (6): 2217–2222. Бибкод : 2014PNAS..111.2217Q . дои : 10.1073/pnas.1314561111 . ПМЦ 3926077 . ПМИД 24379390 .
- ^ Квандт, Эрик М; Голлихар, Джимми; Блаунт, Закари Д.; Эллингтон, Эндрю Д.; Георгиу, Джордж; Баррик, Джеффри Э. (14 октября 2015 г.). «Точная настройка потока цитратсинтазы усиливает и совершенствует метаболические инновации в эксперименте по эволюции Ленски» . электронная жизнь . 4 . doi : 10.7554/eLife.09696 . ПМЦ 4718724 . ПМИД 26465114 .
- ^ Тернер, Кэролайн Б.; Блаунт, Закари Д.; Митчелл, Дэниел Х.; Ленски, Ричард Э. (17 июня 2015 г.). «Эволюция и сосуществование в ответ на ключевое нововведение в долгосрочном эксперименте по эволюции Escherichia coli ». bioRxiv 10.1101/020958 .
- ^ Саэй, Тина Хесман (9 сентября 2015 г.). «Эксперимент показал, что вымирание в лабораторной бутылке было случайностью» . Новости науки . Проверено 4 июня 2016 г.
- ^ Тернер, Кэролайн Б.; Блаунт, Закари Д.; Ленски, Ричард Э.; Коэн, Фредерик М. (18 ноября 2015 г.). «Воспроизведение эволюции для проверки причины вымирания одного экотипа в экспериментально развитой популяции» . ПЛОС ОДИН . 10 (11): e0142050. Бибкод : 2015PLoSO..1042050T . дои : 10.1371/journal.pone.0142050 . ПМЦ 4651540 . ПМИД 26581098 .
- ^ Jump up to: Перейти обратно: а б Ван Хофвеген, Дастин Дж.; Ховде, Кэролайн Дж.; Миннич, Скотт А.; Силхави, Ти Джей (1 апреля 2016 г.). «Быстрая эволюция использования цитрата Escherichia coli путем прямого отбора требует citT и dctA» . Журнал бактериологии . 198 (7): 1022–1034. дои : 10.1128/JB.00831-15 . ПМЦ 4800869 . ПМИД 26833416 .
- ^ Jump up to: Перейти обратно: а б с Рот, Джон Р.; Менье-Патен, Софи; Силхави, Ти Джей (1 апреля 2016 г.). «Переосмысление долгосрочных экспериментов по эволюции: является ли отложенная адаптация примером исторической случайности или следствием прерывистого отбора?» . Журнал бактериологии . 198 (7): 1009–1012. дои : 10.1128/JB.00110-16 . ПМЦ 4800865 . ПМИД 26883821 .
- ^ «Похожие данные, разные выводы | Журнал Scientist Magazine®» . Ученый . Проверено 21 мая 2016 г.
- ^ Jump up to: Перейти обратно: а б Ленски, Ричард (20 февраля 2016 г.). «Об эволюции использования цитрата» . Возвращение к Теллиамеду .
Дальнейшее чтение [ править ]
- Докинз, Ричард (2009). «Сорок пять тысяч поколений эволюции в лаборатории» . Величайшее шоу на Земле: Доказательства эволюции . Нью-Йорк: Свободная пресса. стр. 116–33 . ISBN 978-1-4165-9478-9 .
- Джон Тиммер (17 ноября 2013 г.). «Спустя 50 000 поколений бактерии все еще развивают большую приспособленность» . Арс Техника .
Внешние ссылки [ править ]
- E. coli Сайт проекта долгосрочной экспериментальной эволюции
- Бактерии совершают серьезный эволюционный сдвиг в лаборатории. Боб Холмс New Scientist, 9 июня 2008 г.
- Эволюция: прошлое, настоящее и будущее. Архивировано 18 июля 2017 г. в Wayback Machine Ричард Ленски.
- Список публикаций по эксперименту
- Интернет-публикация статьи о быстрой эволюции использования цитрата.