Химическая защита

Химическая защита – это стратегия, используемая многими организмами, чтобы избежать потребления путем производства токсичных или репеллентных метаболитов или химических предупреждений, которые провоцируют защитные изменения в поведении. [1] [2] Производство защитных химических веществ происходит у растений, грибов и бактерий, а также у беспозвоночных и позвоночных животных. [3] [4] Класс химических веществ, вырабатываемых организмами, которые считаются защитными, можно рассматривать в строгом смысле как применимый только к тем, которые помогают организму избежать травоядности или хищничества . [1] Однако различие между типами химического взаимодействия является субъективным, и защитные химические вещества также можно рассматривать как средство защиты от снижения пригодности со стороны вредителей , паразитов и конкурентов . [5] [6] [7] Репеллентными, а не токсичными метаболитами являются алломоны, подкатегория сигнальных метаболитов, известная как семиохимические вещества . Многие химические вещества, используемые в защитных целях, представляют собой вторичные метаболиты, полученные из первичных метаболитов , которые служат физиологическим целям в организме. [1] Вторичные метаболиты, вырабатываемые растениями, потребляются и изолируются различными членистоногими, а токсины, обнаруженные у некоторых земноводных, змей и даже птиц, можно отнести к добыче членистоногих. [8] [9] Существует множество особых случаев, когда антихищническую адаптацию млекопитающих можно рассматривать как химическую защиту. [10]
Прокариоты и грибы [ править ]
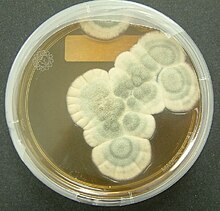
Бактерии родов Chromobacterium , Janthinobacterium и Pseudoalteromonas производят токсичный вторичный метаболит виолацеин для сдерживания хищничества простейших. Виолацеин выделяется при употреблении бактерий, убивая простейших. Другая бактерия, Pseudomonas aeruginosa , агрегирует в биопленки, чувствительные к кворуму , которые могут способствовать скоординированному высвобождению токсинов для защиты от нападения простейших. Жгутиконосцам дали возможность вырасти, и они присутствовали в биопленке P. aeruginosa, выращенной в течение трех дней, но через семь дней жгутиков не было обнаружено. Это позволяет предположить, что концентрированное и скоординированное высвобождение внеклеточных токсинов биопленками имеет больший эффект, чем одноклеточные выделения. [11] Рост бактерий ингибируется не только бактериальными токсинами, но и вторичными метаболитами, продуцируемыми грибами. [4] [7] Самый известный из них, впервые обнаруженный и опубликованный Александром Флемингом в 1929 году, описывает антибактериальные свойства «сока плесени», выделенного из Penicillium notatum . Он назвал вещество пенициллином, и оно стало первым в мире антибиотиком широкого спектра действия. [4] [12] Многие грибы являются либо патогенными сапрофитами , либо живут внутри растений, не причиняя им вреда в качестве эндофитов , и было документально подтверждено, что многие из них производят химические вещества с антагонистическим действием против различных организмов, включая грибы, бактерии и простейшие. [4] Исследования копрофильных грибов обнаружили противогрибковые агенты, которые снижают приспособленность конкурирующих грибов. [7] Кроме того, склероции Aspergillus flavus содержали ряд ранее неизвестных афлавининов , которые были гораздо более эффективны в уменьшении хищничества грибовидного жука Carpophilus hemipterus , чем афлатоксины , которые также продуцирует A. flavus. Было высказано предположение, что алкалоиды спорыньи, микотоксины , продуцируемые Claviceps purpurea , возможно, эволюционировал, чтобы препятствовать травоядности растения-хозяина. [7]
Lichen[editЛишайник
Лишайники демонстрируют химическую защиту, аналогичную упомянутой выше. Их защита действует против травоядных животных и патогенов, включая бактериальные, вирусные и грибковые разновидности. [13] [14] лишайника производит различные химические вещества С этой целью микобионт лишайника через углеводороды, вырабатываемые фотобионтом . [15] [16] Однако одно защитное химическое вещество может служить нескольким целям. Усниновая кислота , например, оказывает антибактериальное, вирусное и грибковое действие. [17] [18] Такие защитные химические вещества могут храниться в различных типах тканей слоевища лишайника или накапливаться на гифах микобионтов в виде внеклеточных кристаллов. [15]
Кислоты, продуцируемые микобионтами, включая, помимо прочего, эверниновую , стиктиновую и скваматовую кислоты, проявляют аллелопатию, более конкретно, защитные химические вещества лишайников могут ингибировать основной путь метаболизма в конкурирующих лишайниках, мхах , микроорганизмах и сосудистых растениях . [13] [15] Задокументированные аллелопатические мишени включают сосну обыкновенную, ель белую и садовые сорта томата, капусты, салата и перца. [15] Антимикробная активность лишайников также опосредована различными кислотами, вырабатываемыми микобионтами, такими как леканоровая и гирофоровая . Было обнаружено, что подобные защитные химикаты подавляют травоядных животных и насекомых. Некоторые из этих защитных соединений от лишайников также обладают фармацевтическим потенциалом. [15] [18]
В 2004 году гибель сотен лосей недалеко от Роулинза, штат Вайоминг, была связана с употреблением в пищу щитового лишайника перекати-поле ( Xanthoparmelia chromorochroa ) . Эта необычайно мощная химическая защита нерегулярна, учитывая, что такие отравления случаются очень редко, а употребление этого лишайника довольно регулярно. [19]
Растения [ править ]
Существует множество литературы по защитной химии вторичных метаболитов, вырабатываемых наземными растениями, и их антагонистическому воздействию на вредителей и патогены, вероятно, из-за того, что человеческое общество зависит от крупномасштабного сельскохозяйственного производства для поддержания глобальной торговли. С 1950-х годов в растениях было зарегистрировано более 200 000 вторичных метаболитов. [20] Эти соединения служат различным физиологическим и аллелохимическим целям и обеспечивают достаточный запас для эволюции защитных химических веществ. Примеры распространенных вторичных метаболитов, используемых растениями в качестве химической защиты, включают алкалоиды , фенолы и терпены . [21] Защитные химические вещества, используемые во избежание употребления в пищу, можно в общих чертах охарактеризовать как токсины или вещества, снижающие пищеварительную способность травоядных животных. Хотя в широком смысле токсины определяются как любое вещество, вырабатываемое одним организмом, которое снижает приспособленность другого организма, в более конкретном смысле токсины — это вещества, которые непосредственно влияют и ослабляют функционирование определенных метаболических путей. [22] [23] Токсины представляют собой второстепенные компоненты (<2% сухого веса), активны в небольших концентрациях и в большем количестве присутствуют в цветах и молодых листьях. С другой стороны, неперевариваемые соединения составляют до 60% сухой массы тканей и преимущественно обнаруживаются в зрелых древесных породах. [23] Многие алкалоиды, пиретрины и фенолы являются токсинами. Танины являются основными ингибиторами пищеварения и представляют собой полифенольные соединения с большой молекулярной массой. Лигнин и целлюлоза являются важными структурными элементами растений и обычно плохо перевариваются. Танины также токсичны в отношении патогенных грибов в естественных концентрациях в различных древесных тканях. [1] Некоторые химические вещества, производимые растениями, не только полезны в качестве сдерживающих факторов для патогенов или потребителей, но и эффективны для подавления конкурентов. Было обнаружено, что два отдельных кустарниковых сообщества в калифорнийском чапарале производят фенольные соединения и летучие терпены, которые накапливаются в почве и препятствуют росту различных трав рядом с кустарниками. Другие растения росли только после того, как пожар уничтожил кусты, но травы впоследствии вымерли после того, как кусты вернулись. [6] Хотя основное внимание уделялось широкомасштабным закономерностям в наземных растениях, Пол и Феникал в 1986 году продемонстрировали множество вторичных метаболитов в морских водорослях, которые предотвращали питание или вызывали смертность у бактерий, грибов, иглокожих, рыб и брюхоногих моллюсков. [24] В природе вредители также представляют собой серьезную проблему для растительных сообществ, что приводит к совместной эволюции химической защиты растений и метаболических стратегий травоядных животных по детоксикации растительной пищи. [25] [14] Растения поедают различные беспозвоночные, но наибольшее внимание уделяется насекомым. Насекомые являются широко распространенными сельскохозяйственными вредителями и иногда встречаются в такой высокой плотности, что могут лишить поля посевов. [26]
Животные [ править ]
Наземные членистоногие [ править ]

Существует множество стратегий, которые наземные членистоногие используют для химической защиты. Первая из этих стратегий включает прямое использование вторичных метаболитов. [27] Многие насекомые неприятны хищникам и выделяют раздражающие вещества или выделяют ядовитые соединения, вызывающие заболевания или смерть при проглатывании. Вторичные метаболиты, полученные из растительной пищи, также могут улавливаться насекомыми и использоваться для производства собственных токсинов. [25] [28] Одним из наиболее известных примеров этого является бабочка-монарх , которая связывает яд, полученный из растения молочая . Среди наиболее успешных отрядов насекомых, использующих эту стратегию, — жуки ( Coleoptera ), кузнечики ( Orthoptera ), а также мотыльки и бабочки ( Lepidoptera ). [29] [30] Насекомые также биосинтезируют уникальные токсины, и хотя секвестрация токсинов из пищевых источников считается энергетически выгодной стратегией, это оспаривается. [25] [31] ), ассоциированные с маракуйей, Бабочки из трибы Heliconiini (подсемейство Heliconiinae либо изолируют, либо синтезируют защитные химические вещества de novo , но мотыльки рода Zygaena (семейство Zygaenidae) развили способность либо синтезировать, либо изолировать свои защитные химические вещества посредством конвергенции. [25] Некоторые жесткокрылые изолируют вторичные метаболиты для использования в качестве защитных химических веществ, но большинство из них биосинтезируют свои собственные de novo . Анатомические структуры развились для хранения этих веществ, некоторые из них циркулируют в гемолимфе и высвобождаются, что связано с явлением, называемым рефлекторным кровотечением . [28]
Использование химической сигнализации и обнаружения является еще одной стратегией химической защиты. Выявление хищников и быстрое и адекватное реагирование является преимуществом и приводит к повышению приспособленности. [2] Эти защитные реакции могут включать (но не ограничиваются ими) реакции избегания и бегства, защиту потомства, агрессивное поведение и применение «прямой защиты» (т.е. токсинов или защитных химикатов, аналогичных стратегии бабочки-монарха, описанной выше). [2] Например, плодовая мушка ( Rhagoletis basiola ) может химически обнаружить ближайшего паразитоида (организм, который действует как паразит и хищник) и остановить его откладку яиц. [32] Отсрочка откладки яиц может снизить риск нападения хищников и подпадает под категорию защиты потомства. [2] Паутинный клещ ( Tetranychus urticae ) может реагировать на летучие вещества хищников в окружающей среде и предпочитает питаться в местах, где нет признаков хищника. [33] Точно так же паутинные клещи также способны чувствовать поврежденные части тела особей того же вида или сородичей и проявлять такое же поведение избегания, как и сигналы хищника. [34] Более того, паутинные клещи демонстрируют такое же поведение при откладке яиц, как и плодовая мушка, и перед откладкой яиц предпочитают перемещаться в места, где нет признаков хищника. Паутинные клещи не будут избегать областей с другими летучими веществами, не являющимися хищниками, а это означает, что эти организмы способны химически отличать угрозу от неопасной. [2] Паразитические осы ( Aphidius uzbekistanicus ) также чувствуют летучие вещества своего хищника, гиперпаразитоида (паразита, хозяином которого является другой паразит), и летают в новые районы, лишенные химических сигналов, демонстрируя такое же поведение избегания, как и паутинный клещ. [35]
С другой стороны, химическое обнаружение хищников или угроз может спровоцировать агрессивное поведение у некоторых наземных членистоногих, а не поведение бегства и избегания. [2] Polybia paulista , веспидовая оса, представляет собой социальный вид, который добывает пищу и защищается в соответствии со сложными социальными структурами. [36] Эти осы научились обнаруживать феромоны в яде представителей одного и того же вида. Обнаружение летучих веществ в яде сородичей позволяет веспидовым осам обнаружить ближайшую угрозу. При обнаружении эти феромоны вызывают атакующее поведение у представителей одного и того же вида. Затем эти осы будут работать вместе, чтобы победить угрозу. [37] Точно так же медоносные пчелы ( Apis mellifera scutellata ) при угрозе выделяют предупреждающий феромон. Эти феромоны усиливают защиту медоносных пчел, увеличивая продолжительность жалящего поведения у всех близлежащих пчел. [38]
Тля, мелкие насекомые, питающиеся соком растений, демонстрируют множество стратегий химической защиты. [39] [40] У тли есть структуры, называемые рожками, вдоль задней стороны брюшка, которые используются для доставки секретов, содержащих как летучие, так и нелетучие соединения. [40] Летучие соединения служат прежде всего сигнальными феромонами . Феромоны — это химические вещества, выделяемые одним человеком и вызывающие реакцию у другого. Нелетучие соединения, такие как воск, используются в качестве ядовитых клеев, которыми тля намазывает своих врагов. Эти мазки используются для смертельного связывания ротового аппарата, антенн, ног и т. д. хищников, а это означает, что эти соединения обычно используются больше для физической защиты, чем для химической защиты. [41] [40] Гороховая тля ( Acyrthosiphon pisum ) вырабатывает предупреждающее химическое вещество под названием (E)-β-фарнезен, которое выделяется в виде летучего соединения в присутствии хищников или предполагаемых угроз. [40] Во многих случаях тля в ответ покидает место кормления в поисках места, где нет тревожных феромонов. [2] Кроме того, гороховая тля хорошо реагирует на то, какие хищники находятся в их районе, поскольку они могут химически определять, что представляет собой угрозу, и соответствующим образом корректировать свою реакцию. Например, гороховая тля может идентифицировать Adalia bipunctata , жука-божью коровку, по химическим признакам хищника. Известно, что гороховая тля, почуяв этого хищника, производит больше потомства с крыльями. [42] Крылатое потомство способно лучше избегать хищников; однако крылатые особи менее плодовиты. Этот компромисс между крыльями и плодовитостью показывает успех этой конкретной защитной стратегии. [2] В «расслабленных» условиях или условиях, в которых отсутствуют признаки хищника, рождается больше бескрылых потомков. [42]

Системы химической защиты тли весьма специфичны. (E)-β-фарнезен, феромон тревоги, о котором говорилось выше, используется многими видами тли. [40] При высвобождении (E)-β-фарнезен будет расширяться всего на 2–3 сантиметра в диаметре. [43] Это защищает более дальних особей от сигнального химического вещества, поэтому они не испытывают ненужных пауз в кормлении или ненужной реакции. [40] Более того, эти химические сигналы тревоги обнаруживаются структурами на усиках тли, которые используют специализированные связывающие белки. Предупреждающие химические вещества должны накопиться до определенного минимума в связывающих белках, прежде чем произойдет ответная реакция. [44] [45] Эти факторы используются для подчеркивания специфики систем химической защиты тли. [40] Более того, используемые химические предупреждения также весьма специфичны, а метод распространения тревожного феромона может вызвать различную реакцию. Например, Ceratovacuna lanigera , тростниковая шерстистая тля, имеет два способа распространения тревожных феромонов. При угрозе феромоны тревоги могут выделяться либо в виде капель, либо в виде мазка. Когда сигнал тревоги срабатывает в виде капли из роговицы тли, местные представители вида реагируют индивидуально и либо избегают этого места, либо покидают его. Однако, когда феромоны тревоги распространяются на хищника, другие представители того же вида начинают совместную атаку. [46] Как обсуждалось выше, восковые мазки роговицы обычно используются для физической защиты тли от хищника. Однако в этом случае химические сигналы воска вызывают изменение поведения; следовательно, эту конкретную стратегию можно считать химической защитой. [40]
Другие организмы смогли воспользоваться сложной химической защитой тлей, чтобы повысить свою приспособленность. [40] Химическая мимикрия — мощный инструмент химической защиты. [47] Lysiphlebus fabarum , паразитоид тли, способен имитировать химические выделения конкретных тлей при проникновении в их колонии. Эта мимикрия служит «химическим камуфляжем» и защищает этих паразитоидов, поскольку они остаются незамеченными в колониях тли. [48] Chrysopalossonae , златоглазка , использует воск шерстистой ольховой тли для химической маскировки от муравьев-муравьев (подсемейства Formicinea ), которые научились избегать нападения на тлю. [49] Это означает, что находящиеся поблизости муравьиные муравьи будут игнорировать златоглазку, как и шерстистую ольховую тлю. Это еще один случай, когда восковые выделения используются для химической защиты, а не для физической. [ нужна ссылка ]
Морские беспозвоночные [ править ]
Морские беспозвоночные используют разнообразные стратегии химической защиты. Некоторые из этих стратегий включают: производство вторичных метаболитов, хранение и модификацию вторичных метаболитов другого организма, химические предупреждения, предупреждения о хищниках, фагомимикрию и химическую «одежду». Успех этих стратегий подтверждается количеством видов, обладающих такой химической защитой. [50]

Морские губки из типа Porifera — лишь один пример морских беспозвоночных, которые получают выгоду от производства вторичных метаболитов . [51] Губки обладают способностью производить свои собственные вторичные метаболиты, а не полагаться на хранение и модификацию химической защиты другого организма. [51] Роль некоторых наблюдаемых вторичных метаболитов до сих пор неизвестна; однако есть данные, подчеркивающие тот факт, что большое количество вторичных метаболитов используется в защитных целях. [51] Например, существует обратная зависимость между количеством вторичных метаболитов внутри губки и количеством спикул, присутствующих в самом организме. [52] Спикулы представляют собой острые игольчатые структуры, выступающие из губки и используемые в качестве средства физической защиты. [53] Вторичные метаболиты и спикулы имеют обратную зависимость, поскольку с увеличением количества вторичных метаболитов количество спикул уменьшается. [52] Это приводит к идее, что вторичные метаболиты действительно используются в защитных целях, и губкам больше не приходится полагаться на физическую защиту. [51] Кроме того, многие губки, вырабатывающие вторичные метаболиты, токсичны для потенциальных хищников. [54] Губки, которые производят большее количество вторичных метаболитов, меньше подвергаются нападениям хищников, что подтверждает идею о том, что вторичные метаболиты используются в качестве защитного механизма. [51]
Хранение и модификация вторичных метаболитов являются полезной стратегией для многих морских беспозвоночных. Они способны изолировать ранее существовавшие химические вещества, не тратя энергию на производство самих вторичных метаболитов. [51] Например, голожаберники , также известные как морские слизни, обладают как «пассивной», так и «активной» формой химической защиты. Морские слизни плотоядны, и центральную часть их рациона составляют морские губки, которые, как обсуждалось выше, производят свои собственные защитные вторичные метаболиты. [51] Ключевой особенностью химической защиты морских слизней является их способность хранить и повторно использовать химические вещества, вырабатываемые потребляемыми ими организмами. [55] Например, морская губка производит пигменты, которые придают им яркие цвета. Пигменты губок накапливаются в морских слизняках во время их питания, что позволяет морским слизнякам маскироваться в окружающей среде. [56] Цвет морских слизней зависит от того, какую губку они едят. Например, морской слизень, который кажется розовым, когда его обнаруживают питающимся розовой губкой, может стать зеленым при миграции на зеленую губку. [56] Этот камуфляж можно рассматривать как «случайную» или пассивную форму химической защиты. [51] Более активная форма химической защиты, обнаруженная у морских слизней, — это их способность хранить и использовать защитные вторичные метаболиты, вырабатываемые губками. [51] Морские слизни обладают двумя механизмами хранения защитных химикатов. Первый из этих механизмов — хранение химических веществ внутри спины (или «задней стороны»). [57] Этот механизм хранения выгоден, поскольку защитные химические вещества расположены вблизи поверхности морского слизняка и легко доступны для выделения любой слизи. [51] Второй механизм защитного химического хранения, демонстрируемый морскими слизняками, заключается в сохранении вторичных метаболитов в других областях их тела. Например, некоторые морские слизни хранят вторичные метаболиты в своем пищеварительном тракте. [51] Морские слизни, использующие эту стратегию для хранения вторичных метаболитов, имеют механизмы применения защитных химических веществ, когда это необходимо. Морские слизни филогенетически связаны с морскими улитками . [58] Одним из наиболее отличительных факторов между этими двумя морскими беспозвоночными является то, что у морских улиток есть панцирь, а у морских слизней его нет. Эта потеря панциря дает представление об успехе стратегии химической защиты морских слизней. [51] При использовании защитных химикатов снаряды становятся ненужными и энергетически дорогими, что приводит к потере этих защитных структур. Тот факт, что морские слизни могут эффективно выживать и уклоняться от хищников без использования панциря, подчеркивает успех хранения и модификации вторичных метаболитов в качестве защитного механизма. [51]
Использование химических предупреждений и сигналов тревоги в качестве защитного механизма используется многими морскими беспозвоночными. [50] Этот механизм основан на том, что беспозвоночные выделяют и воспринимают химические сигналы в водной среде и в результате изменяют свое поведение. [50] Например, моллюски научились чувствовать феромоны хищников в окружающей воде и реагировать таким образом, что скрывают свое присутствие от этих хищников. [59] Моллюски , относящиеся ко многим видам моллюсков, питаются насосным способом. «Накачка» происходит, когда моллюски втягивают окружающую воду, питаются присутствующими в ней микроорганизмами и выпускают только что отфильтрованную воду. [59] Хищники моллюсков, а именно голубые панцирные крабы и трубачи, способны идентифицировать свою добычу, ощущая химические сигналы, присутствующие в фильтрованной воде. Моллюски научились химически чувствовать хищников, находящихся вверху по течению. [50] Когда поблизости чувствуется хищник, моллюски меняют свое поведение и прекращают сцеживание, чтобы уменьшить количество потребительских сигналов. Хищникам больше не придется идти по химическому следу в поисках моллюска. [59] Моллюски возобновляют свою работу только тогда, когда потребительские сигналы отсутствуют. [59] В этом сценарии и хищник, и жертва полагаются на наличие вторичных метаболитов. Хищники используют эти химические вещества в качестве механизма охоты, а моллюски используют их в качестве сигнала тревоги, вызывающего их поведенческую реакцию. синие панцирные крабы ( Callinectes sapidus Похожий механизм защиты имеют ), обычные хищники моллюсков; однако вместо химического обнаружения хищников в местной среде они способны улавливать химические предупреждения, излучаемые представителями того же вида. [60] Эти крабы, когда им причиняют вред, излучают химическое предупреждение, специфичное для конкретного вида, то есть эти химические предупреждения обнаруживаются только другими крабами с синим панцирем. Эти предупреждения могут исходить от поврежденных целых крабов или частей тела синих панцирных крабов. [60] Эти химические сигналы предупреждают других избегать зон повышенного риска. [60] Использование химических предупреждений и феромонов сигнализации — это механизм, используемый многими морскими беспозвоночными, моллюски и синие панцирные крабы — лишь два примера этой защитной стратегии. [50]

Морские зайцы используют форму химической защиты, называемую фагомимикрией. [50] В отличие от широко распространенного использования ранее обсуждавшихся стратегий химической защиты, фагомимикрия специфична для морских зайцев. [61] Фагомимикрия, как следует из названия, представляет собой разновидность химической мимикрии. Многие организмы научились использовать мимикрию, поскольку это весьма успешный механизм химической защиты. [47] Морские зайцы при нападении быстро выделяют в окружающую среду туман химических веществ. Химическое облако состоит из двух основных частей: чернил и опала. [61] Чернила, попадая в воду, физически скрывают морского зайца от хищника. Опаловый туман представляет собой смесь химических веществ, которые имитируют сигналы пищи хищника и поэтому действуют как пищевой стимул. [61] Целью опалового химического облака является предоставление более сильного пищевого стимула, чем сам морской заяц. [61] В целом облако работает на то, чтобы подавить и отвлечь хищника. В замешательстве хищник нападет на химическую смесь, а не на самого зайца, давая ему время сбежать. [50]
Некоторые морские беспозвоночные способны приобретать химическую защиту, покрываясь другими организмами, имеющими защитные вторичные метаболиты. Этот защитный механизм описывается как «химическая одежда». [50] Было замечено, что беспозвоночные используют множество различных организмов в качестве одежды. К ним относятся губки, бактерии и морские водоросли. [50] Интересно, что многие морские беспозвоночные, использующие этот механизм защиты, являются травоядными. Эти травоядные животные предпочитают использовать водоросли в качестве одежды, а не еды, то есть они больше ценят водоросли за их защитные способности, а не как потенциальную пищу. [62] В полевых условиях беспозвоночные, такие как атлантический краб-декоратор ( Libinia dubia ), «одетые» в ядовитые водоросли, подвергаются значительно меньшему нападению хищников, чем их раздетые сородичи. [62] Морские беспозвоночные и химически защищенный организм способны образовывать симбиотические отношения, в результате чего морские беспозвоночные приобретают долговременную химическую защиту. [50]
Позвоночные животные [ править ]

Позвоночные животные также могут биосинтезировать защитные химические вещества или выделять их из растений или добычи. [9] [31] Секвестрированные соединения наблюдались у лягушек, натрициновых змей и двух родов птиц, Pitohui и Ifrita . [9] Предполагается, что некоторые хорошо известные соединения, такие как тетродотоксин, вырабатываемый тритонами и рыбой-фугу, [63] происходят от беспозвоночных. Буфадиенолиды , защитные химические вещества, вырабатываемые жабами, были обнаружены в железах натрициновых змей, используемых для защиты. [9]
Земноводные [ править ]
Лягушки приобретают токсины, необходимые для химической защиты, вырабатывая их либо через железы на коже, либо через пищу. Источником токсинов в их рационе являются прежде всего членистоногие , от жуков до многоножек. Когда необходимые пищевые компоненты отсутствуют, например, в неволе, лягушка больше не способна вырабатывать токсины, что делает их неядовитыми. Профиль токсинов может даже меняться в зависимости от сезона, как в случае с восходящей мантеллой , чья диета и пищевое поведение различаются в зависимости от влажного и засушливого сезона. [64]
Эволюционное преимущество производства таких токсинов заключается в отпугивании хищников. Есть данные, позволяющие предположить, что способность вырабатывать токсины развивалась вместе с апосематической окраской , действуя как визуальный сигнал для хищников, чтобы они помнили, какие виды неприятны на вкус. [19]
Хотя токсины, вырабатываемые лягушками, часто называют ядовитыми, дозы токсинов достаточно малы, поэтому они скорее вредны, чем ядовиты. Однако компоненты токсинов, а именно алкалоиды , очень активны в ионных каналах . Таким образом, они нарушают работу нервной системы жертвы, делая их гораздо более эффективными. Внутри самих лягушек токсины накапливаются и доставляются через небольшие специализированные транспортные белки. [65]

Помимо обеспечения защиты от хищников, токсины, выделяемые лягушками, интересуют медицинских исследователей. Ядовитые древолазы семейства Dendrobatidae выделяют батрахотоксин . Этот токсин может действовать как миорелаксант, стимулятор сердца или анестетик. Несколько видов лягушек выделяют эпибатидин, исследование которого дало несколько важных результатов. Было обнаружено, что лягушки сопротивляются отравлению за счет замены одной аминокислоты, которая снижает чувствительность целевых рецепторов к токсину, но при этом сохраняет функцию рецептора. Это открытие дает представление о роли белков, нервной системы и механизма химической защиты, которые способствуют будущим биомедицинским исследованиям и инновациям.
Млекопитающие [ править ]
Некоторые млекопитающие могут выделять неприятный запах жидкости из анальных желез , например панголин. [66] и некоторые представители семейств Mephitidae и Mustelidae, включая скунсов , ласк и хорьков . [67] У однопроходных есть ядовитые шпоры, используемые для защиты от хищников. [68] а медленные лори (Primates: Nycticebus) выделяют яд, который, по-видимому, эффективен для отпугивания как хищников, так и паразитов. [69] Также было продемонстрировано, что физический контакт с медленным лори (без укуса) может вызвать реакцию у людей, действуя как контактный яд. [70]
См. также [ править ]
Ссылки [ править ]
- ^ Jump up to: Перейти обратно: а б с д Беренбаум М.Р. (январь 1995 г.). «Химия защиты: теория и практика» . Труды Национальной академии наук Соединенных Штатов Америки . 92 (1): 2–8. Бибкод : 1995ПНАС...92....2Б . дои : 10.1073/pnas.92.1.2 . ПМК 42807 . ПМИД 7816816 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час Дике, Марсель; Гростал, Пол (2001). «Химическое обнаружение естественных врагов членистоногими: экологическая перспектива» . Ежегодный обзор экологии и систематики . 32 (1): 1–23. doi : 10.1146/annurev.ecolsys.32.081501.113951 . ISSN 0066-4162 .
- ^ Клукас Б (2010). «Защитная химия». В журнале Breed MD, Мур Дж. (ред.). Энциклопедия поведения животных . Оксфорд: Академическая пресса. стр. 481–486. дои : 10.1016/B978-0-08-045337-8.00293-X . ISBN 9780080453378 .
- ^ Jump up to: Перейти обратно: а б с д Келлер Н.П., Тернер Дж., Беннетт Дж.В. (декабрь 2005 г.). «Вторичный метаболизм грибов – от биохимии к геномике». Обзоры природы. Микробиология . 3 (12): 937–47. дои : 10.1038/nrmicro1286 . ПМИД 16322742 . S2CID 23537608 .
- ^ Уолтерс Д. (2011). Защита растений: защита от атак болезнетворных микроорганизмов, травоядных и растений-паразитов . Джон Уайли и сыновья.
- ^ Jump up to: Перейти обратно: а б Уиттакер Р.Х., Фини П.П. (февраль 1971 г.). «Аллелохимика: химические взаимодействия между видами». Наука . 171 (3973). Нью-Йорк, штат Нью-Йорк: 757–70. Бибкод : 1971Sci...171..757W . дои : 10.1126/science.171.3973.757 . JSTOR 1730763 . ПМИД 5541160 .
- ^ Jump up to: Перейти обратно: а б с д Глоер Дж. Б. (1995). «Химия грибкового антагонизма и защиты». Канадский журнал ботаники . 73 (С1): 1265–1274. дои : 10.1139/b95-387 .
- ^ Лесли Э.Н. (1999). «Иметь токсины и есть их тоже: изучение природных источников химической защиты многих животных дает новое понимание природной аптечки» . Бионаука . 49 (12): 945–950. дои : 10.1525/bisi.1999.49.12.945 .
- ^ Jump up to: Перейти обратно: а б с д Савицкий А.Х., Мори А., Хатчинсон Д.А., Сапорито Р.А., Бургхардт Г.М., Лиллиуайт Х.Б., Мейнвальд Дж. (сентябрь 2012 г.). «Секвестрированные защитные токсины у четвероногих позвоночных: принципы, закономерности и перспективы будущих исследований» . Химиоэкология . 22 (3): 141–158. Бибкод : 2012Checo..22..141S . дои : 10.1007/s00049-012-0112-z . ПМЦ 3418492 . ПМИД 22904605 .
- ^ Хетти А., Овегес Б., Мориц А.М., Драхос Л., Капон Р.Дж., Ван Баскирк Дж. и др. (декабрь 2019 г.). «Изменения химической защиты позвоночных, вызванные хищниками» . Журнал экологии животных . 88 (12): 1925–1935. Бибкод : 2019JAnEc..88.1925H дои : 10.1111/1365-2656.13083 . ПМИД 31408536 .
- ^ Мац С., Кьеллеберг С. (июль 2005 г.). «С крючка - как бактерии выживают при выпасе простейших». Тенденции в микробиологии . 13 (7): 302–7. дои : 10.1016/j.tim.2005.05.009 . ПМИД 15935676 .
- ^ Флеминг А (1929). «Об антибактериальном действии культур пенициллий с особым указанием на их использование при выделении B. influenzae» . Британский журнал экспериментальной патологии . 10 (3): 226–236. ПМК 2048009 .
- ^ Jump up to: Перейти обратно: а б Перри, Найджел Б.; Бенн, Майкл Х.; Бреннан, Нерида Дж.; Берджесс, Элейн Дж.; Эллис, Гилл; Галлоуэй, Дэвид Дж.; Лоример, Стивен Д.; Тэнгни, Раймонд С. (1999). «Противомикробная, противовирусная и цитотоксическая активность новозеландских лишайников». Лихенолог . 31 (6): 627–636. дои : 10.1006/lich.1999.0241 . S2CID 86005089 .
- ^ Jump up to: Перейти обратно: а б Лоури, Джеймс Д. (1989). «Вторичные соединения лишайника: доказательства соответствия между противотравоядными и антимикробными функциями». Бриолог . 92 (3): 326–328. дои : 10.2307/3243401 . JSTOR 3243401 .
- ^ Jump up to: Перейти обратно: а б с д и Мольнар К. и Фаркас Э. (2010). Современные результаты биологической активности вторичных метаболитов лишайников: обзор. Журнал естественных исследований C, 65 (3–4), 157–173.
- ^ Бхаттачарья С., Дип П.Р., Сингх С. и Наяк Б. (2016). Вторичные метаболиты лишайника и его биологическая активность. Являюсь. J. PharmTech Res, 6 (6), 1-7.
- ^ Эммерих Р., Гиз И., Ланге О.Л. и Прокш П. (1993). Токсичность и антифидантная активность соединений лишайника в отношении многоядного растительноядного насекомого Spodopteralittoralis. Фитохимия, 33(6), 1389-1394.
- ^ Jump up to: Перейти обратно: а б Фурманек Л., Чарнота П. и Сиворд М.Р. (2022). Обзор потенциала веществ лишайника как противогрибковых средств: влияние экстрактов и вторичных метаболитов лишайника на грибы Fusarium. Архив микробиологии, 204 (8), 1-31.
- ^ Jump up to: Перейти обратно: а б Кук, В.Е., Райсбек, М.Ф., Корниш, Т.Э., Уильямс, Э.С., Браун, Б., Хайатт, Г., и Кригер, Т.Дж. (2007). Парез и смерть лося (Cervus elaphus) из-за интоксикации лишайником в Вайоминге. Журнал болезней диких животных, 43(3), 498-503.
- ^ Хартманн Т. (2007). «От отходов до экохимикатов: пятидесятилетние исследования вторичного метаболизма растений». Фитохимия . 68 (22–24): 2831–46. Бибкод : 2007PChem..68.2831H . doi : 10.1016/j.phytochem.2007.09.017 . ПМИД 17980895 .
- ^ Левин Д.А. (1976). «Химическая защита растений от патогенов и травоядных». Ежегодный обзор экологии и систематики . 7 (1): 121–159. doi : 10.1146/annurev.es.07.110176.001005 .
- ^ Уиттсток Ю, Гершензон Дж (август 2002 г.). «Конститутивные растительные токсины и их роль в защите от травоядных и патогенов». Современное мнение в области биологии растений . 5 (4): 300–7. дои : 10.1016/S1369-5266(02)00264-9 . ПМИД 12179963 .
- ^ Jump up to: Перейти обратно: а б Кейтс Р.Г., Роудс Д.Ф. (1977). «Способы производства средств химической защиты от травоядных животных в растительных сообществах». Биохимическая систематика и экология . 5 (3): 185–193. Бибкод : 1977BioSE...5..185C . дои : 10.1016/0305-1978(77)90003-5 .
- ^ Пол В.Дж., Феникал W (1986). «Химическая защита тропических зеленых водорослей отряда Caulerpales» . Серия «Прогресс в области морской экологии» . 34 : 157–169. Бибкод : 1986MEPS...34..157P . дои : 10.3354/meps034157 .
- ^ Jump up to: Перейти обратно: а б с д Фюрстенберг-Хэгг Дж., Загробельный М., Йоргенсен К., Фогель Х., Мёллер Б.Л., Бак С. (2014). «Химическая защита, сбалансированная секвестрацией и биосинтезом de novo у специалиста по чешуекрылым» . ПЛОС ОДИН . 9 (10): е108745. Бибкод : 2014PLoSO...9j8745F . дои : 10.1371/journal.pone.0108745 . ПМК 4191964 . ПМИД 25299618 .
- ^ Ломер С.Дж., Бейтман Р.П., Джонсон Д.Л., Лангевальд Дж., Томас М. (2001). «Биологическая борьба с саранчой и кузнечиками». Ежегодный обзор энтомологии . 46 (1): 667–702. дои : 10.1146/annurev.ento.46.1.667 . ПМИД 11112183 .
- ^ Эйснер, Томас; Мейнвальд, Джеррольд (16 сентября 1966 г.). «Защитные выделения членистоногих». Наука . 153 (3742): 1341–1350. Бибкод : 1966Sci...153.1341E . дои : 10.1126/science.153.3742.1341 . ISSN 0036-8075 . ПМИД 17814381 .
- ^ Jump up to: Перейти обратно: а б Деттнер К. (1987). «Хемосистематика и эволюция химической защиты жуков». Ежегодный обзор энтомологии . 32 (1): 17–48. doi : 10.1146/annurev.en.32.010187.000313 .
- ^ Шмидт Дж.О. (2008). «Яды и токсины насекомых». Энциклопедия энтомологии . Нидерланды: Спрингер. стр. 4076–4089. дои : 10.1007/978-1-4020-6359-6_3957 . ISBN 978-1-4020-6359-6 .
- ^ Триго-младший (2000). «Химия защиты от хищников с помощью вторичных соединений у неотропических чешуекрылых: факты, перспективы и предостережения» . Журнал Бразильского химического общества . 11 (6): 551–561. doi : 10.1590/S0103-50532000000600002 .
- ^ Jump up to: Перейти обратно: а б Мебс Д. (январь 2001 г.). «Токсичность у животных. Тенденции эволюции?». Токсикон . 39 (1): 87–96. дои : 10.1016/S0041-0101(00)00155-0 . ПМИД 10936625 .
- ^ Хоффмайстер, Т.С.; Ройтберг, Б.Д. (21 марта 1997 г.). «Контрразведка в системе насекомых-травоядных-паразитоидов» . естественные науки . 84 (3): 117–119. Бибкод : 1997NW.....84..117H . дои : 10.1007/s001140050358 . ISSN 0028-1042 . S2CID 43784335 .
- ^ Гростал, П. (1 июля 1999 г.). «Прямые и косвенные признаки риска нападения хищников влияют на поведение и воспроизводство добычи: пример взаимодействия акаринов» . Поведенческая экология . 10 (4): 422–427. дои : 10.1093/beheco/10.4.422 . ISSN 1465-7279 .
- ^ Гростал, П.; Дике, М. (23 марта 2000 г.). «Распознавание врагов: функциональный подход к оценке риска по добыче». Поведенческая экология и социобиология . 47 (4): 258–264. дои : 10.1007/s002650050663 . ISSN 0340-5443 . S2CID 2071560 .
- ^ Хеллер, К.; Миха, С.Г.; Шульц, С.; Франке, В.; Пикетт, Дж. А. (1 февраля 1994 г.). «Вызванное врагом расселение осы-паразита». Эксперименты . 50 (2): 182–185. дои : 10.1007/BF01984961 . ISSN 1420-9071 . S2CID 6380388 .
- ^ О'Доннелл, Шон (январь 1998 г.). «Определение репродуктивной касты у эусоциальных ос (Hymenoptera: Vespidae)» . Ежегодный обзор энтомологии . 43 (1): 323–346. дои : 10.1146/annurev.ento.43.1.323 . ISSN 0066-4170 . ПМИД 15012393 .
- ^ Манзоли-Пальма, МФ; Гобби, Н.; Пальма, MS (1998). «Феромоны СИГНАЛИЗАЦИИ и влияние запаха куколки на агрессивность Polybia paulista (Ihering) (Hymenoptera: Vespidae)» . Журнал ядовитых животных и токсинов . 4 (1): 61–69. дои : 10.1590/s0104-79301998000100006 . ISSN 0104-7930 .
- ^ Нуньес, Дж (31 декабря 1997 г.). «Тревожный феромон вызывает стрессовую аналгезию через опиоидную систему у медоносной пчелы» . Физиология и поведение . 63 (1): 75–80. дои : 10.1016/s0031-9384(97)00391-0 . ISSN 0031-9384 . ПМИД 9402618 . S2CID 8788442 .
- ^ Смит, К. Майкл; Бойко, Елена Владимировна (2007). «Молекулярные основы устойчивости растений и защитных реакций на питание тлей: современное состояние» . Энтомология экспериментальная и прикладная . 122 (1): 1–16. Бибкод : 2007EEApp.122....1S . дои : 10.1111/j.1570-7458.2006.00503.x . ISSN 0013-8703 . S2CID 54875407 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я Мишо, Япония (07 января 2022 г.). «Экологическое значение корней тли и их выделений» . Ежегодный обзор энтомологии . 67 (1): 65–81. doi : 10.1146/annurev-ento-033021-094437 . ISSN 0066-4170 . ПМИД 34995085 . S2CID 263481309 .
- ^ Барри, Адема; Оно, Кадзуро (6 июля 2016 г.). «Выделения роговицы Uroleucon nigrotuberculatum (Homoptera: Aphididae) как последняя пуля против личинок божьих коровок». Энтомологическая наука . 19 (4): 410–415. дои : 10.1111/ens.12221 . ISSN 1343-8786 . S2CID 89225187 .
- ^ Jump up to: Перейти обратно: а б Диксон, Афганистан; Агарвала, БК (7 августа 1999 г.). «Изменения в истории жизни тли, вызванные божьими коровками» . Труды Лондонского королевского общества. Серия Б: Биологические науки . 266 (1428): 1549–1553. дои : 10.1098/rspb.1999.0814 . ISSN 0962-8452 . ПМК 1690176 .
- ^ Миха, Стефан Г.; Висс, Урс (1996). «Феромон тревоги тли (E)-?-фарнезен: хозяин находит кайромон для первичного паразитоида тли Aphidius uzbekistanicus (Hymenoptera: Aphidiinae)» . Химиоэкология . 7 (3): 132–139. Бибкод : 1996Checo...7..132M . дои : 10.1007/bf01245965 . ISSN 0937-7409 . S2CID 28800099 .
- ^ Ван, Линг; Би, Ин-Донг; Лю, Мин; Ли, Вэй; Лю, Мяо; Ди, Шу-Фэн; Ян, Шуай; Фан, Чао; Бай, Лей; Лай, Юн-Кай (25 июля 2019 г.). «Идентификация и анализ профилей экспрессии белков, связывающих запахи, у соевой тли, глицинов aphis (Hemiptera: Aphididae)» . Наука о насекомых . 27 (5): 1019–1030. дои : 10.1111/1744-7917.12709 . ISSN 1672-9609 . ПМИД 31271503 . S2CID 195798550 .
- ^ ван Эмден, Хельмут Ф.; Дингли, Джейн; Дьюхерст, Сара Ю.; Пикетт, Джон А.; Вудкок, Кристин М.; Уодхамс, Лестер Дж. (25 августа 2014 г.). «Влияние искусственной диеты на выработку тревожного феромона Myzus persicae » . Физиологическая энтомология . 39 (4): 285–291. дои : 10.1111/phen.12074 . ISSN 0307-6962 . S2CID 84758675 .
- ^ Аракаки, Норио (1989). «Тревожный феромон, вызывающий реакцию нападения и побега у шерстистой тли сахарного тростника Ceratovacuna lanigera (Homoptera, Pemphigidae)» . Журнал этологии . 7 (2): 83–90. дои : 10.1007/bf02350028 . ISSN 0289-0771 . S2CID 24333580 .
- ^ Jump up to: Перейти обратно: а б Деттнер, К; Липерт, К. (1994). «Химическая мимикрия и камуфляж» . Ежегодный обзор энтомологии . 39 (1): 129–154. doi : 10.1146/annurev.en.39.010194.001021 . ISSN 0066-4170 .
- ^ Расех, Араш; Мишо, JP; Харази-Пакдель, Азиз; Аллахьяри, Хоссейн (2010). «Муравьиная мимикрия паразитоида тли Lysiphlebus fabarum » . Журнал науки о насекомых . 10 (126): 126. дои : 10.1673/031.010.12601 . ISSN 1536-2442 . ПМК 3016887 . ПМИД 20879920 .
- ^ Эйснер, Томас; Хикс, Карен; Эйснер, Мария; Робсон, Дуглас С. (17 февраля 1978 г.). « Стратегия личинки хищного насекомого «Волк в овечьей шкуре» . Наука . 199 (4330): 790–794. Бибкод : 1978Sci...199..790E . дои : 10.1126/science.199.4330.790 . ISSN 0036-8075 . ПМИД 17836295 . S2CID 11558335 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж Хэй, Марк Э. (1 января 2009 г.). «Морская химическая экология: химические сигналы и сигналы, структурирующие морские популяции, сообщества и экосистемы» . Ежегодный обзор морской науки . 1 (1): 193–212. Бибкод : 2009ARMS....1..193H . дои : 10.1146/annurev.marine.010908.163708 . ISSN 1941-1405 . ПМК 3380104 . ПМИД 21141035 .
- ^ Jump up to: Перейти обратно: а б с д и ж г час я дж к л м Фолкнер, диджей; Гизелин, МТ (1983). «Химическая защита и эволюционная экология доридовых голожаберных и некоторых других заднежаберных брюхоногих моллюсков» . Серия «Прогресс в области морской экологии» . 13 : 295–301. Бибкод : 1983MEPS...13..295F . дои : 10.3354/meps013295 . ISSN 0171-8630 .
- ^ Jump up to: Перейти обратно: а б Уокер, Роджер П.; Томпсон, Дженис Э.; Фолкнер, Д. Джон (1980). «Сестертерпены из Spongia idia» . Журнал органической химии . 45 (24): 4976–4979. дои : 10.1021/jo01312a032 . ISSN 0022-3263 .
- ^ Сетманн, Инго; Вёрхайде, Герт (1 апреля 2008 г.). «Структура и состав спикул известковой губки: обзор и сравнение со структурно родственными биоминералами» . Микрон . 39 (3): 209–228. дои : 10.1016/j.micron.2007.01.006 . ISSN 0968-4328 . ПМИД 17360189 .
- ^ Бакус, Джеральд Дж. (30 января 1981 г.). «Механизмы химической защиты на Большом Барьерном рифе, Австралия» . Наука . 211 (4481): 497–499. Бибкод : 1981Sci...211..497B . дои : 10.1126/science.7455691 . ISSN 0036-8075 . ПМИД 7455691 .
- ^ Томпсон, Дженис Э.; Уокер, Роджер П.; Раттен, Стивен Дж.; Фолкнер, Д. Джон (1982). «Механизм химической защиты голожаберного cadlina luteomarginata» . Тетраэдр . 38 (13): 1865–1873. дои : 10.1016/0040-4020(82)80035-5 . ISSN 0040-4020 .
- ^ Jump up to: Перейти обратно: а б Шредль, Майкл; Грау, Хосе Х (2006). «Голожаберные с отдаленных южных чилийских островов Гуамблин и Ипун (архипелаг Чонос, 44–45 ° ю.ш.), с повторным описанием Rostanga pulchra MacFarland, 1905» . Revista Chilena de Historyia Natural . 79 (1). дои : 10.4067/s0716-078x2006000100001 . ISSN 0716-078X .
- ^ Чимино, Г.; Де Роза, С.; Де Стефано, С.; Содано, Г. (1982). «Химическая защита четырех средиземноморских голожаберников» . Сравнительная биохимия и физиология. Часть B: Сравнительная биохимия . 73 (3): 471–474. дои : 10.1016/0305-0491(82)90061-x . ISSN 0305-0491 .
- ^ ГОСЛАЙНЕР, ТМ (1981). «Происхождение и взаимоотношения примитивных представителей Opisthobranchia (Mollusca: Gastropoda)» . Биологический журнал Линнеевского общества . 16 (3): 197–225. дои : 10.1111/j.1095-8312.1981.tb01848.x . ISSN 0024-4066 .
- ^ Jump up to: Перейти обратно: а б с д Сми, Делберт Л.; Вайссбург, Марк Дж. (2006). «Замалчивание: силы окружающей среды уменьшают способность восприятия двустворчатой добычи» . Экология . 87 (6): 1587–1598. doi : 10.1890/0012-9658(2006)87[1587:cuefdt]2.0.co;2 . hdl : 1853/36765 . ISSN 0012-9658 . ПМИД 16869434 .
- ^ Jump up to: Перейти обратно: а б с Фернер, MC; Сми, Д.Л.; Чанг, Ю.П. (2005). «Крабы-каннибалы реагируют на запах раненых особей: опасность или ужин?» . Серия «Прогресс в области морской экологии» . 300 : 193–200. Бибкод : 2005MEPS..300..193F . дои : 10.3354/meps300193 . ISSN 0171-8630 .
- ^ Jump up to: Перейти обратно: а б с д Киклайтер, Синтия Э.; Шабани, Шкельцен; Джонсон, Пол М.; Дерби, Чарльз Д. (2005). «Морские зайцы используют новые средства противохищной химической защиты» . Современная биология . 15 (6): 549–554. дои : 10.1016/j.cub.2005.01.057 . ISSN 0960-9822 . ПМИД 15797024 . S2CID 11562464 .
- ^ Jump up to: Перейти обратно: а б Стахович, Джон Дж.; Хэй, Марк Э. (1999). «Сокращение хищничества посредством химического камуфляжа: косвенное воздействие защиты растений на травоядных животных» . Экология . 80 (2): 495–509. doi : 10.1890/0012-9658(1999)080[0495:rptcmc]2.0.co;2 . hdl : 1853/36756 . ISSN 0012-9658 .
- ^ Фурман Ф.А. (декабрь 1986 г.). «Тетродотоксин, тарихатоксин и хирикитоксин: исторические перспективы». Анналы Нью-Йоркской академии наук . 479 (1 тетродотоксин): 1–14. Бибкод : 1986NYASA.479....1F . дои : 10.1111/j.1749-6632.1986.tb15556.x . ПМИД 3468842 . S2CID 44741246 .
- ^ Московиц Н.А., Роланд А.Б., Фишер Е.К., Ранайворасо Н., Видудес С., Агилар М.Т. и др. (26 декабря 2018 г.). Чавес А.В. (ред.). «Сезонные изменения в рационе питания и химической защите лазающей лягушки Mantella (Mantella laevigata)» . ПЛОС ОДИН . 13 (12): e0207940. Бибкод : 2018PLoSO..1307940M . дои : 10.1371/journal.pone.0207940 . ПМК 6306172 . ПМИД 30586404 .
- ^ Кэти С.Н., Альварес-Буйя А., Берд Г.Д., Видудес С., Роланд А.Б., Тапиа Э.Э. и др. (июнь 2019 г.). «Молекулярная физиология химической защиты ядовитой лягушки» . Журнал экспериментальной биологии . 222 (Часть 12): jeb204149. дои : 10.1242/jeb.204149 . ПМИД 31138640 . S2CID 169034903 .
- ^ «Фолидота (панголины)» . Сеть разнообразия животных .
- ^ Андерсен К.К., Бернштейн Д.Т. (1980). «Соединения серы в куньих». Природные соединения серы . Бостон, Массачусетс: Спрингер. стр. 399–406. дои : 10.1007/978-1-4613-3045-5_35 . ISBN 978-1-4613-3047-9 .
- ^ Хурум Дж.Х., Луо З.Х., Килан-Яворовска З. (2006). «Были ли млекопитающие изначально ядовитыми?» (PDF) . Acta Palaeontologica Polonica . 51 (1): 1–11.
- ^ Некарис К.А., Мур Р.С., Роде Э.Дж., Фрай Б.Г. (сентябрь 2013 г.). «Безумно, плохо и опасно знать: биохимия, экология и эволюция яда медленного лори» . Журнал ядовитых животных и токсинов, включая тропические болезни . 19 (1): 21. дои : 10.1186/1678-9199-19-21 . ПМЦ 3852360 . ПМИД 24074353 .
- ^ Гардинер М., Уэлдон А., Пойндекстер С.А., Гибсон Н., Некарис К.А. (2018). «Опрос специалистов, обращающихся с медленными лори (приматы: Nycticebus): оценка вредного воздействия укусов медленных лори» . Журнал исследований ядов . 9 : 1–7. ПМК 6055083 . ПМИД 30090322 .