Аксональный транспорт
Аксональный транспорт , также называемый аксоплазматическим транспортом или аксоплазматическим потоком , представляет собой клеточный процесс, ответственный за перемещение митохондрий , липидов , синаптических пузырьков , белков и других органелл обратно и нейрона к телу клетки через цитоплазму его аксона , называемую аксоплазмой. . [1] Поскольку длина некоторых аксонов составляет порядка метров, нейроны не могут полагаться на диффузию для переноса продуктов ядра и органелл к концам своих аксонов. Аксональный транспорт также отвечает за перемещение молекул, предназначенных для деградации, из аксона обратно в тело клетки, где они расщепляются лизосомами . [2]
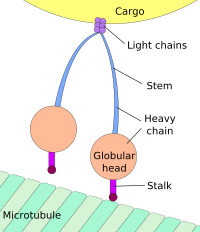
Движение к телу клетки называется ретроградным транспортом , а движение к синапсу — антероградным транспортом . [3] [4]
Механизм
[ редактировать ]
Подавляющее большинство аксональных белков синтезируется в теле нейрональных клеток и транспортируется по аксонам. Некоторая трансляция мРНК была продемонстрирована внутри аксонов. [5] [6] Аксональный транспорт происходит на протяжении всей жизни нейрона и необходим для его роста и выживания. Микротрубочки (состоящие из тубулина ) проходят по всей длине аксона и обеспечивают основные цитоскелетные «дорожки» для транспортировки. Кинезин и динеин — это моторные белки , которые перемещают грузы в антероградном (вперед от сомы к кончику аксона) и ретроградном (назад к соме (телу клетки) направлениях соответственно. Моторные белки связывают и транспортируют несколько различных грузов, включая митохондрии , цитоскелетные полимеры . , аутофагосомы и синаптические пузырьки , содержащие нейромедиаторы .
Аксональный транспорт может быть быстрым или медленным, антероградным (от тела клетки) или ретроградным (переносит материалы от аксона к телу клетки).
Быстрый и медленный транспорт
[ редактировать ]Везикулярные грузы перемещаются относительно быстро (50–400 мм/день), тогда как транспорт растворимых (цитозольных) и цитоскелетных белков занимает гораздо больше времени (перемещение менее 8 мм/день). [7] Основной механизм быстрого аксонального транспорта был понятен на протяжении десятилетий, но механизм медленного аксонального транспорта стал ясен только недавно благодаря передовым методам визуализации . [8] Методы флуоресцентной маркировки (например, флуоресцентная микроскопия ) позволили напрямую визуализировать транспорт в живых нейронах.
Недавние исследования показали, что движение цитоскелетных «медленных» грузов на самом деле происходит быстро, но, в отличие от быстрых грузов, они часто останавливаются, что значительно замедляет общую скорость транзита. Этот механизм известен как модель медленного аксонального транспорта «Stop and Go» и был тщательно проверен на транспорте цитоскелетных белковых нейрофиламентов. [9] Перемещение растворимых (цитозальных) грузов более сложное, но, по-видимому, имеет аналогичную основу: растворимые белки организуются в мультибелковые комплексы, которые затем передаются посредством временных взаимодействий с более быстро движущимися грузами, движущимися в быстром аксональном транспорте. [10] [11] [12] Аналогия — разница в тарифах на проезд между пригородными поездами и экспрессами метро. Хотя оба типа поездов движутся между станциями с одинаковой скоростью, пригородному поезду требуется гораздо больше времени, чтобы добраться до конца линии, поскольку он останавливается на каждой станции, тогда как экспресс делает лишь несколько остановок в пути.
Антероградный транспорт
[ редактировать ]Антероградный (также называемый «ортоградным») транспорт — это движение молекул/органелл наружу, от тела клетки (также называемой сомой ) к синапсу или клеточной мембране .
Антероградное перемещение отдельных грузов (в транспортных везикулах ) как быстрых, так и медленных компонентов по микротрубочкам. [4] опосредуется кинезинами . [2] Некоторые кинезины участвуют в медленном транспорте. [8] однако механизм возникновения «пауз» при транзите медленных составных грузов до сих пор неизвестен.
Выделяют два класса медленного антероградного транспорта: медленный компонент а (SCa), переносящий преимущественно микротрубочки и нейрофиламенты со скоростью 0,1-1 миллиметра в день, и медленный компонент b (SCb), переносящий более 200 разнообразных белков и актин со скоростью до 6 миллиметров в день. [8] Медленный компонент b, который также несет актин, транспортируется со скоростью 2-3 миллиметра в день по аксонам клеток сетчатки.
Во время реактивации из латентного периода вирус простого герпеса (ВПГ) вступает в свой литический цикл и использует механизмы антероградного транспорта для миграции из нейронов дорсальных корешков ганглиев в кожу или слизистую оболочку, на которые он впоследствии влияет. [13]
Груз-рецептор для моторов антероградного транспорта, кинезины, был идентифицирован как белок-предшественник амилоида (APP), родительский белок, который производит сенильные бляшки, обнаруженные при болезни Альцгеймера. [14] Пептид из 15 аминокислот на карбоксильном конце цитоплазмы АРР связывается с высоким сродством к обычному кинезину-1 и опосредует транспорт экзогенного груза в гигантский аксон кальмара. [15]
Марганец, контрастный агент для Т 1 -взвешенной МРТ, после стереотаксической инъекции перемещается в мозг экспериментальных животных путем антероградного транспорта и тем самым выявляет схемы при МРТ всего мозга у живых животных, как это впервые сделали Робия Паутлер, Элейн Бирер и Расс Джейкобс. Исследования на мышах с нокаутом легкой цепи кинезина-1 показали, что Mn 2+ перемещается посредством транспорта на основе кинезина в зрительном нерве и в мозге. Транспорт как в проекциях гиппокампа, так и в зрительном нерве также зависит от АРР. [16] Транспортировка из гиппокампа в передний мозг снижается с возрастом, а место назначения меняется из-за наличия бляшек болезни Альцгеймера. [17]
Ретроградный транспорт
[ редактировать ]Ретроградный транспорт переносит молекулы/органеллы от концов аксона к телу клетки . Ретроградный аксональный транспорт опосредуется цитоплазматическим динеином и используется, например, для отправки химических сообщений и продуктов эндоцитоза, направляющихся к эндолизосомам из аксона обратно в клетку. [2] Работая со средней скоростью in vivo примерно 2 мкм/сек, [18] [19] быстрый ретроградный транспорт может преодолевать 10-20 сантиметров в день. [2]
Быстрый ретроградный транспорт возвращает использованные синаптические пузырьки и другие материалы в сому и информирует сому о состоянии окончаний аксонов. Ретроградный транспорт переносит сигналы выживания из синапса обратно в тело клетки, такие как TRK, рецептор фактора роста нервов. [20] Некоторые патогены используют этот процесс для проникновения в нервную систему. Они входят в дистальные кончики аксона и перемещаются в сому ретроградным транспортом. Примеры включают столбнячный токсин и вирусы простого герпеса, бешенства и полиомиелита. При таких инфекциях задержка между заражением и появлением симптомов соответствует времени, необходимому возбудителям для достижения соматических клеток. [21] Вирус простого герпеса перемещается по аксонам в обоих направлениях в зависимости от его жизненного цикла, при этом ретроградный транспорт доминирует в полярности поступающих капсидов. [22]
Последствия прерывания
[ редактировать ]Всякий раз, когда аксональный транспорт ингибируется или прерывается, нормальная физиология становится патофизиологией, что может привести к накоплению аксоплазмы, называемой аксональным сфероидом . Поскольку транспорт аксонов может быть нарушен множеством способов, аксональные сфероиды можно наблюдать при многих различных классах заболеваний, включая генетические, травматические, ишемические, инфекционные, токсические, дегенеративные и специфические заболевания белого вещества , называемые лейкоэнцефалопатиями . Некоторые редкие нейродегенеративные заболевания связаны с генетическими мутациями моторных белков, кинезина и динеина , и в этих случаях вполне вероятно, что аксональный транспорт является ключевым игроком в опосредовании патологии. [23] [24] Дисфункциональный аксональный транспорт также связан со спорадическими (распространенными) формами нейродегенеративных заболеваний, таких как болезнь Альцгеймера и Паркинсона . [8] Это главным образом связано с многочисленными наблюдениями о том, что большие скопления аксонов неизменно наблюдаются в пораженных нейронах и что гены, которые, как известно, играют роль в семейных формах этих заболеваний, также предполагаемо играют роль в нормальном аксональном транспорте. Однако прямых доказательств участия аксонального транспорта в последних заболеваниях мало, и другие механизмы (такие как прямая синаптотоксичность) могут иметь большее значение.
Остановка аксоплазматического потока на краю ишемизированных участков при сосудистых ретинопатиях приводит к набуханию нервных волокон, в результате чего образуются мягкие экссудаты или ватные пятна.
Поскольку аксон зависит от аксоплазматического транспорта жизненно важных белков и материалов, повреждение, такое как диффузное повреждение аксона , которое прерывает транспорт, приведет к дегенерации дистального аксона в процессе, называемом валлеровской дегенерацией . Противораковые препараты , которые препятствуют росту рака путем изменения микротрубочек (которые необходимы для деления клеток ), повреждают нервы, поскольку микротрубочки необходимы для аксонального транспорта.
Инфекция
[ редактировать ]Вирус бешенства достигает центральной нервной системы ретроградным аксоплазматическим током. [25] Столбнячный нейротоксин интернализуется в нервно-мышечном соединении посредством связывания белков нидогена и ретроградно транспортируется к соме в сигнальных эндосомах. [26] Нейротропные вирусы, такие как вирусы герпеса, перемещаются внутри аксонов с помощью механизмов клеточного транспорта, как было показано в работе группы Элейн Бирер. [27] [28] Другие инфекционные агенты также подозреваются в использовании аксонального транспорта. [29] Сейчас считается, что такие инфекции способствуют развитию болезни Альцгеймера и других нейродегенеративных неврологических расстройств. [30] [31]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Сабри Дж., О'Коннор Т.П., Киршнер М.В. (июнь 1995 г.). «Аксональный транспорт тубулина в пионерских нейронах Ti1 in situ» . Нейрон . 14 (6): 1247–56. дои : 10.1016/0896-6273(95)90271-6 . ПМИД 7541635 .
- ^ Jump up to: а б с д Озтас Э (2003). «Трассировка нейронов» (PDF) . Нейроанатомия . 2 : 2–5. Архивировано (PDF) из оригинала 25 октября 2005 г.
- ^ Карп Г., ван дер Гир П. (2005). Клеточная и молекулярная биология: концепции и эксперименты (4-е изд.). Джон Уайли. п. 344 . ISBN 978-0-471-46580-5 .
- ^ Jump up to: а б Медведь М.Ф., Коннорс Б.В., Парадсо М.А. (2007). Нейронаука: исследование мозга (3-е изд.). Липпинкотт Уильямс и Уилкинс. п. 41 . ISBN 978-0-7817-6003-4 .
- ^ Джустетто М., Хегде А.Н., Си К., Касадио А., Инокучи К., Пей В., Кандель Э.Р., Шварц Дж.Х. (ноябрь 2003 г.). «Аксональный транспорт эукариотической мРНК фактора элонгации трансляции 1альфа связывает транскрипцию в ядре с долговременным облегчением в синапсе» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (23): 13680–5. Бибкод : 2003PNAS..10013680G . дои : 10.1073/pnas.1835674100 . ПМЦ 263873 . ПМИД 14578450 .
- ^ Да К, Джустетто Си К., Джустетто М., Эткин А., Сюй Р., Янисевич AM, Миниачи MC, Ким Дж. Х., Чжу Х., Кандель Э. Р. (декабрь 2003 г.). «Нейрональная изоформа CPEB регулирует локальный синтез белка и стабилизирует специфическое для синапсов долговременное облегчение аплизии» . Клетка . 115 (7): 893–904. дои : 10.1016/s0092-8674(03)01021-3 . ПМИД 14697206 . S2CID 15552012 .
- ^ Мадей, Сандра; Двенадцать деревьев, Элисон Э.; Мугамян, Армен Дж.; Хольцбаур, Эрика Л.Ф. (октябрь 2014 г.). «Аксональный транспорт: специфические для груза механизмы подвижности и регуляции» . Нейрон . 84 (2): 292–309. дои : 10.1016/j.neuron.2014.10.019 . ПМК 4269290 . ПМИД 25374356 .
- ^ Jump up to: а б с д Рой С., Чжан Б., Ли В.М., Трояновский JQ (январь 2005 г.). «Дефекты аксонального транспорта: общая тема нейродегенеративных заболеваний». Акта Нейропатологика . 109 (1): 5–13. дои : 10.1007/s00401-004-0952-x . ПМИД 15645263 . S2CID 11635065 .
- ^ Браун А. (март 2003 г.). «Аксональный транспорт мембранных и немембранных грузов: единый взгляд» . Журнал клеточной биологии . 160 (6): 817–21. дои : 10.1083/jcb.200212017 . ПМК 2173776 . ПМИД 12642609 .
- ^ Скотт Д.А., Дас Ю, Тан Й, Рой С. (май 2011 г.). «Механистическая логика, лежащая в основе аксонального транспорта цитозольных белков» . Нейрон . 70 (3): 441–54. дои : 10.1016/j.neuron.2011.03.022 . ПМК 3096075 . ПМИД 21555071 .
- ^ Рой С., Уинтон М.Дж., Блэк М.М., Трояновски Дж.К., Ли В.М. (март 2007 г.). «Быстрый и прерывистый котранспорт медленных белков компонента b» . Журнал неврологии . 27 (12): 3131–8. doi : 10.1523/JNEUROSCI.4999-06.2007 . ПМК 6672457 . ПМИД 17376974 .
- ^ Кузнецов А.В. (2011). «Аналитическое решение уравнений, описывающих медленный аксональный транспорт, на основе гипотезы остановки и движения» . Центральноевропейский физический журнал . 9 (3): 662–673. Бибкод : 2011CEJPh...9..662K . дои : 10.2478/s11534-010-0066-0 .
- ^ Холланд DJ, Миранда-Уитнесс М., Боадл Р.А., Армати П., Каннингем А.Л. (октябрь 1999 г.). «Антероградный транспорт белков вируса простого герпеса в аксонах периферических нейронов плода человека: исследование с помощью иммуноэлектронной микроскопии» . Журнал вирусологии . 73 (10): 8503–11. doi : 10.1128/JVI.73.10.8503–8511.1999 . ПМК 112870 . ПМИД 10482603 .
- ^ Сатпут-Кришнан П., ДеДжоргис Дж.А., Конли, член парламента, Джанг М., Бирер Э.Л. (октябрь 2006 г.). «Почтовый индекс пептида, достаточный для антероградного транспорта внутри белка-предшественника амилоида» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (44): 16532–7. Бибкод : 2006PNAS..10316532S . дои : 10.1073/pnas.0607527103 . ПМК 1621108 . ПМИД 17062754 .
- ^ Симстер П.Е., Левенберг М., Паскаль Дж., Шовьер А., Гонсалес А., Кристини В., Бирер Э.Л. (октябрь 2012 г.). «Количественные измерения и моделирование грузо-моторных взаимодействий при быстром транспорте в живом аксоне» . Физическая биология . 9 (5): 055005. Бибкод : 2012PhBio...9e5005S . дои : 10.1088/1478-3975/9/5/055005 . ПМЦ 3625656 . ПМИД 23011729 .
- ^ Галлахер Дж.Дж., Чжан X, Зиомек Г.Дж., Джейкобс Р.Э., Бирер Э.Л. (апрель 2012 г.). «Дефицит аксонального транспорта в схемах гиппокампа и зрительных путях у животных с нокаутом APP, подтвержденный МРТ с усилением марганца» . НейроИмидж . 60 (3): 1856–66. doi : 10.1016/j.neuroimage.2012.01.132 . ПМЦ 3328142 . ПМИД 22500926 .
- ^ Bearer EL, Manifold-Wheeler BC, Medina CS, Gonzales AG, Chaves FL, Jacobs RE (октябрь 2018 г.). «Изменения функциональных схем стареющего мозга и влияние мутированной экспрессии АРР» . Нейробиология старения . 70 : 276–290. doi : 10.1016/j.neurobiolaging.2018.06.018 . ПМК 6159914 . ПМИД 30055413 .
- ^ Гиббс К.Л., Кальмар Б., Сани Дж.Н., Гринсмит Л., Скьяво Г. (январь 2016 г.). «Визуализация аксонального транспорта in vivo в моторных и сенсорных нейронах мышей» . Журнал методов нейробиологии . 257 : 26–33. doi : 10.1016/j.jneumeth.2015.09.018 . ПМК 4666412 . ПМИД 26424507 .
- ^ Сани Дж., Скьяво Дж. (2016). «Старее, но не медленнее: старение не меняет динамику аксонального транспорта сигнальных эндосом in vivo» . Имеет значение . 2 (6). дои : 10.19185/matters.201605000018 .
- ^ Цуй Б, Ву С, Чен Л, Рамирес А, Бирер Э.Л., Ли В.П., Мобли В.К. , Чу С. (август 2007 г.). «Поочередное отслеживание аксонального транспорта NGF в реальном времени с использованием квантовых точек» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (34): 13666–71. Бибкод : 2007PNAS..10413666C . дои : 10.1073/pnas.0706192104 . ЧВК 1959439 . ПМИД 17698956 .
- ^ Саладин, Кеннет. Анатомия и физиология: единство формы и функции. Шестое. Нью-Йорк: McGraw-Hill, 2010. 445. Печать.
- ^ Бирер Э.Л., Брейкфилд XO, Шубак Д., Риз Т.С., ЛаВейл Дж.Х. (июль 2000 г.). «Ретроградный аксональный транспорт вируса простого герпеса: доказательства единого механизма и роли тегумента» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (14): 8146–50. Бибкод : 2000PNAS...97.8146B . дои : 10.1073/pnas.97.14.8146 . ПМК 16684 . ПМИД 10884436 .
- ^ Мэдай С., Твелветрис А.Э., Мугамян А.Дж., Хольцбаур Э.Л. (октябрь 2014 г.). «Аксональный транспорт: специфичные для груза механизмы подвижности и регуляции» . Нейрон . 84 (2): 292–309. дои : 10.1016/j.neuron.2014.10.019 . ПМК 4269290 . ПМИД 25374356 .
- ^ Сани Дж.Н., Россор А.М., Феллоуз А.Д., Тосолини А.П., Скьяво Дж. (декабрь 2019 г.). «Аксональный транспорт и неврологические заболевания» . Нат преподобный Нейрол . 15 (12): 691–703. дои : 10.1038/s41582-019-0257-2 . PMID 31558780 . S2CID 203437348 .
- ^ Митрабхакди Э., Шуангшоти С., Ваннакрайрот П., Льюис Р.А., Сусуки К., Лаотаматас Дж., Хемачуда Т. (ноябрь 2005 г.). «Различия нейропатогенетических механизмов при бешенстве человека и паралитическом бешенстве». Журнал неврологических наук . 238 (1–2): 3–10. дои : 10.1016/j.jns.2005.05.004 . ПМИД 16226769 . S2CID 25509462 .
- ^ Берцени К., Шмиг Н., Брайсон Дж.Б., Уоллес М., Каччин П., Голдинг М., Занотти Г., Гринсмит Л., Ништ Р., Скьяво Г. (ноябрь 2014 г.). «Вход столбнячного токсина. Нидогены являются терапевтическими мишенями для профилактики столбняка» (PDF) . Наука . 346 (6213): 1118–23. дои : 10.1126/science.1258138 . ПМИД 25430769 . S2CID 206560426 .
- ^ Сатпут-Кришнан П., ДеДжоргис Дж.А., Бирер Э.Л. (декабрь 2003 г.). «Быстрый антероградный транспорт вируса простого герпеса: роль белка-предшественника амилоида болезни Альцгеймера» . Стареющая клетка . 2 (6): 305–18. дои : 10.1046/j.1474-9728.2003.00069.x . ПМЦ 3622731 . ПМИД 14677633 .
- ^ Ченг С.Б., Ферланд П., Вебстер П., Бирер Э.Л. (март 2011 г.). «Вирус простого герпеса танцует с белком-предшественником амилоида при выходе из клетки» . ПЛОС ОДИН . 6 (3): e17966. Бибкод : 2011PLoSO...617966C . дои : 10.1371/journal.pone.0017966 . ПМК 3069030 . ПМИД 21483850 .
- ^ Носитель Э.Л., Сатпуте-Кришнан П. (сентябрь 2002 г.). «Роль цитоскелета в жизненном цикле вирусов и внутриклеточных бактерий: дорожки, моторы и машины полимеризации» . Текущие цели по борьбе с наркотиками. Инфекционные заболевания . 2 (3): 247–64. дои : 10.2174/1568005023342407 . ПМЦ 3616324 . ПМИД 12462128 .
- ^ Ицхаки Р.Ф., Лате Р., Балин Б.Дж., Болл М.Дж., Бирер Э.Л., Браак Х. и др. (2016). «Микробы и болезнь Альцгеймера» . Журнал болезни Альцгеймера . 51 (4): 979–84. дои : 10.3233/JAD-160152 . ПМЦ 5457904 . ПМИД 26967229 .
- ^ «Нет места лучше асфальта для этих выносливых микробов». Новый учёный . 206 (2757): 15. 2010. doi : 10.1016/s0262-4079(10)60991-8 .