Бета -шпилька

Бета -шпилька (иногда также называемая бета-высокопоставленной или бета-бета-единицей ) представляет собой простой белка структурный мотив с участием двух бета-нитей , которые похожи на шпильку . Мотив состоит из двух цепей, которые прилегают к первичной структуре , ориентированные в антипараллельном направлении ( N-конце одного листа примыкает к С-конце следующего) и связан короткой петлей из двух-пяти аминокислот Полем Бета -шпильки могут происходить в изоляции или как часть серии связанных с водородными прядями, которые в совокупности составляют бета -лист .
Исследователи, такие как Франциско Бланко и соавт. использовали белок ЯМР, чтобы показать, что бета-лаирпины могут быть образованы из изолированных коротких пептидов в водном растворе, что позволяет предположить, что шпильки могут образовывать сайты нуклеации для складывания белка . [ 1 ]
Классификация
[ редактировать ]Бета-шпильки первоначально были классифицированы исключительно по количеству аминокислотных остатков в своих последовательностях петли, так что они были названы одним рестораном, двумя ресторанами и т. Д. [ 2 ] Эта система, однако, несколько неоднозначна, поскольку она не учитывает, являются ли остатки, которые сигнализируют конец шпильки, по отдельности или вдвойне связаны с водородом друг с другом. С тех пор Milner-White и поэт предложил улучшенное средство классификации. [ 3 ] Бета -шпильки разбиваются на четыре различных класса, как показано на рисунке публикации 1. Каждый класс начинается с наименьшего возможного количества остатков петли и постепенно увеличивает размер петли, удаляя водородные связи в бета -листе. Основная шпилька класса 1-это петля с одним разрешением, где связанные остатки имеют две водородные связи. Одна водородная связь затем удаляется, чтобы создать петлю из трех ресурсов, которая является вторичной шпилькой класса 1. Одиночные остатки подсчитываются в последовательности петли, но также сигнализируют о конце петли, таким образом определяя эту шпильку как три-остаток петля. Эта отдельная водородная связь затем удаляется, чтобы создать третичную шпильку; петля из пяти ресурсов с двойным связанными остатками. Этот рисунок продолжается бесконечно и определяет все бета -шпильки в классе. Класс 2 следует той же схеме, начиная с цикла с двумя остатками с завершающими остатками, которые имеют две водородные связи. Класс 3 начинается с трех остановок и 4 класса с четырьмя остановками. Класс 5 не существует, поскольку первичная шпилька уже определена в классе 1. Пик Эта схема классификации не только учитывает различные степени водородной связи, но и говорит что -то о биологическом поведении шпильки. Одиночные аминокислотные замены могут разрушить конкретную водородную связь, но не развернут шпильку или изменять свой класс. С другой стороны, аминокислотные вставки и делеции должны будут разворачиваться и реформировать все Beta Strand , чтобы избежать бета -выпуклости во вторичной структуре. Это изменит класс шпильки в процессе. Поскольку замена являются наиболее распространенными аминокислотными мутациями, белок может потенциально подвергаться конверсии, не влияя на функциональность бета -шпильки. [ 3 ]
Динамика складывания и привязки
[ редактировать ]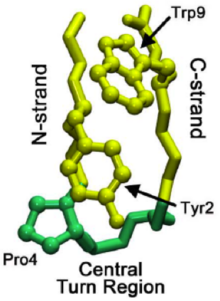

Понимание механизма, посредством которого микродомены могут помочь пролить свет на складывание складывания целых белков . Исследования бета-шпильки, называемой хиньолином (см. Чиньолин на протеопедии ), обнаружили пошаговый процесс складывания, который способствует складыванию бета-волос. Эта шпилька имеет особенности последовательности, аналогичные более чем 13 000 известных шпильков, и, таким образом, может служить более общей моделью для образования бета -шпильки. Образование нативной области поворота сигнализирует о складном каскаде для начала, где нативный поворот - это то, что присутствует в последней сложенной структуре.
При складывании общих белков поворот может возникнуть не в области нативного поворота, а в C-потоке бета-волосы. Затем этот поворот распространяется через C-цепь (бета-нить, ведущая к C-конце), пока не достигнет области нативного поворота. Иногда взаимодействия остатков , приведшие к местной области поворота, слишком сильны, вызывая обратное распространение. Однако, как только нативный поворот формируется, взаимодействие между пролинами и остатками триптофана (наблюдаемые на изображении справа) в области помогают стабилизировать поворот, предотвращая «откат» или растворение.
Исследователи полагают, что повороты не возникают в N-странице из-за повышенной жесткости (часто вызванной пролином, ведущим к области нативного поворота) и менее конформационными вариантами. Первоначальная образование поворота происходит примерно в 1 мкс. После того, как был установлен первоначальный поворот, были предложены два механизма относительно того, как остальные складки бета-волосы: гидрофобный коллапс с перестройками на уровне боковой цепи или более принятым механизмом, похожим на молнию. [ 4 ]
Мотив петли β-волосы можно найти во многих макромолекулярных белках. Тем не менее, небольшие и простые β-волос могут существовать самостоятельно. Чтобы ясно видеть это, белок домена PIN1 показан слева в качестве примера.
Белки, которые богаты β-листами, также называемые доменами WW , функционируют путем прилипания к богатым пролину и/или фосфорилированными пептидами для опосредованного белка-белкового взаимодействия . «WW» относится к двум остаткам триптофана (W), которые сохраняются в последовательности и помогают в складывании β-листов для получения небольшого гидрофобного ядра. [ 5 ] Эти остатки триптофана можно увидеть ниже (справа) в красном.
Этот фермент связывает свой лиганд через силы Ван-дер-Ваальса консервативных триптофанов и богатых пролином областей лиганда. Другие аминокислоты могут затем ассоциироваться с гидрофобным ядром структуры β-волосы для обеспечения безопасного связывания. [ 6 ]
Также распространено находить остатки пролина в фактической части петли β-волосы, поскольку эта аминокислота жесткая и способствует формированию «поворота». Эти остатки пролина можно рассматривать как красные боковые цепи на изображении домена PIN1 WW внизу (слева).
 |
 |
Искусственно спроектированная бета-волоса
[ редактировать ]Конструкция пептидов, которые применяют структуру β-волосы (не полагаясь на связывание металлов, необычные аминокислоты или дисульфидные сшивки), добилась значительного прогресса и дала представление о динамике белка. В отличие от α-списков , β-волос не стабилизируются обычной паттерной водородной связи. В результате ранние попытки требовали не менее 20–30 аминокислотных остатков для достижения стабильных третичных складок β-волос. Тем не менее, этот нижний предел был уменьшен до 12 аминокислот благодаря получению стабильности, обеспечиваемым путем включения паров поперечного потока триптофана-трипптофана. Было показано, что две негидрогеновые триптофанные пары переключаются в мотив, похожий на молнию, стабилизируя структуру β-волосы, в то же время позволяя ему оставаться растворимым в воде . Структура ЯМР β-пептида β - пептида триптофана (TRPZIP) демонстрирует стабилизирующий эффект благоприятного взаимодействия между соседними индольные кольца. [ 7 ]

Синтез пептидов Trpzip β-Hairpin включает в себя Photogatches, которые облегчают точный контроль над складыванием. Несколько аминокислот в ходе поворота заменяются азобензолом , который может быть вызван для перехода от транс на конформацию CIS на свет при 360 нм. Когда фрагмент азобензола находится в конформации CIS, аминокислотные остатки правильно выравниваются, чтобы принять образование β-волосы. Однако транс-конформация не имеет надлежащей геометрии поворота для β-волосы. [ 8 ] Это явление можно использовать для изучения пептидной конформационной динамики с фемтосекундной спектроскопией поглощения. [ 8 ]
Ссылки
[ редактировать ]- ^ Бланко, FJ; Ривас, Г.; Серрано Л. (1994). «Короткий линейный пептид, который складывается в нативную стабильную бета-волос в водном растворе». Nat Struct Biol . 1 (9): 584–590. doi : 10.1038/nsb0994-584 . PMID 7634098 . S2CID 35065527 .
- ^ Сибанда, BL; Blundell, TL; Тортон, JM (1985). «Конформации бета-волос в белковых структурах». Природа (Лондон) 316 170–174.
- ^ Jump up to: а беременный Milner-White, J.; Поэт, Р. (1986). «Четыре класса бета-волос в белках». Биохимический журнал 240 289–292.
- ^ Jump up to: а беременный Энемар, Сёрен; Kurniawan, Nicholas A.; Раджагопалан, Радж (11 сентября 2012 г.). «β-волос формируется путем свертывания с C-концевого: топологическое руководство динамики раннего складывания» . Научные отчеты . 2 : 649. Bibcode : 2012natsr ... 2e.649e . doi : 10.1038/srep00649 . PMC 3438464 . PMID 22970341 .
- ^ Джагер, Маркус; Deechongkit, Songpon; Koepf, Edward K.; Нгуен, Хуби; Гао, Цзянмин; Powers, Evan T.; Gruebele, Martin; Келли, Джеффри В. (2008). «Понимание механизма складывания β-листа с химической и биологической точки зрения». Биополимеры . 90 (6): 751–758. doi : 10.1002/bip.21101 . PMID 18844292 .
- ^ Кей, BK; Уильямсон, депутат; Судол М. Журнал FASEB. 2000, 14, 231–241.
- ^ Cochran, Andrea G.; Скелтон, Николас Дж.; Starovasnik, Melissa A. (8 мая 2001 г.). «Триптофан-молнии: стабильные, мономерные β-волокны» . Труды Национальной академии наук . 98 (10): 5578–5583. Bibcode : 2001pnas ... 98.5578c . doi : 10.1073/pnas.091100898 . ISSN 0027-8424 . PMC 33255 . PMID 11331745 .
- ^ Jump up to: а беременный Донг, Шоу-Лян; Löweneck, Markus; Schrader, Tobias E.; Schreier, Wolfgang J.; Зинт, Вольфганг; Мородер, Луис; Реннер, Кристиан (23 января 2006 г.). «Фотоконтролируемый β-волос пептид». Химия - европейский журнал . 12 (4): 1114–1120. doi : 10.1002/chem.200500986 . ISSN 1521-3765 . PMID 16294349 .