Базальный (филогенетика)
В филогенетике укорененного филогенетического базальным называется направление основания ( или корня) дерева или кладограммы . Этот термин может более строго применяться только к узлам, прилегающим к корню, или более широко применяться к узлам, считающимся близкими к корню. Обратите внимание, что существующие таксоны, лежащие на ветвях, соединяющихся непосредственно с корнем, не более тесно связаны с корнем, чем любые другие существующие таксоны. [ 1 ] [ 2 ] [ 3 ]
Хотя всегда должно быть две или более одинаково «базальных» клад, вырастающих из корня каждой кладограммы, эти клады могут сильно различаться по таксономическому рангу . [ н 1 ] видовое разнообразие , или и то, и другое. [ н 2 ] Если C — базальная клада внутри D , имеющая самый низкий ранг из всех базальных клад внутри D , [ н 3 ] C можно описать как базальный этого таксон ранга в D. пределах [ н 4 ] Концепция « ключевой инновации » подразумевает некоторую степень корреляции между эволюционными инновациями и диверсификацией . [ 4 ] [ 5 ] [ 6 ] [ n 5 ] Однако такая корреляция не делает данный случай предсказуемым, поэтому предковые признаки не следует приписывать членам менее богатой видами базальной клады без дополнительных доказательств. [ 1 ] [ 2 ] [ 7 ] [ 8 ] [ n 6 ]
В общем, клада A является более базальной, чем клада B, B является подгруппой сестринской группы A если или группы A. самой [ н 7 ] В контексте больших групп термин «базальный» часто используется в широком смысле для обозначения позиций, более близких к корню, чем большинство, и в таких случаях могут появляться такие выражения, как «очень базальный». «Основная клада» относится к группе, которая включает в себя все составляющие клады, за исключением базальной клады самого низкого ранга в более крупной кладе, примером которой являются основные эвдикоты . Ни один из существующих таксонов не находится ближе к корню, чем любой другой.
Использование
[ редактировать ]Базальная группа в более строгом смысле образует сестринскую группу по отношению к остальной части более крупной клады. [ нужна ссылка ] как в следующем случае:
|
Базальная клада №2 |
Хотя на такой кладограмме легко идентифицировать базальную кладу, уместность такой идентификации зависит от точности и полноты диаграммы. В этом примере часто предполагается, что конечные ветви кладограммы изображают все существующие таксоны данного ранга внутри клады; это одна из причин, по которой термин «базальный» очень обманчив, поскольку отсутствие дополнительных видов в одной кладе воспринимается как свидетельство морфологического родства с предковыми таксонами. Кроме того, эта квалификация не гарантирует, что будет представлено разнообразие вымерших таксонов (которые могут быть плохо изучены). [ нужна ссылка ]
В филогенетике термин базальный не может быть объективно применен к кладам организмов, но имеет тенденцию применяться избирательно и более спорно к группам или линиям. [ н 8 ] Считается, что они обладают наследственными признаками или самими такими предполагаемыми наследственными чертами . При описании персонажей «предковые» или « плезиоморфные » предпочтительнее «базальных» или « примитивных », последний из которых может нести ложный оттенок неполноценности или отсутствия сложности. [ 1 ] Термины «глубокое ветвление» или «раннее ветвление» схожи по значению и в равной степени могут искажать существующие таксоны, которые лежат на ветвях, соединяющихся непосредственно с корневым узлом, как имеющие больше состояний предкового признака. [ 1 ] [ 2 ]
Несмотря на повсеместное использование базального , систематики стараются избегать его использования, поскольку его применение к существующим группам является ненужным и вводит в заблуждение. [ 3 ] Этот термин чаще применяется, когда одна ветвь (которая считается «базальной») менее разнообразна, чем другая ветвь (это ситуация, в которой можно было бы ожидать найти базальный таксон более низкого минимального ранга). Этот термин может быть двусмысленным, поскольку он также относится к направлению корня дерева, который представляет собой гипотетического предка; следовательно, это может неточно означать, что сестринская группа более богатой видами клады демонстрирует наследственные черты. [ 8 ] Существующая базальная группа может напоминать или не напоминать последнего общего предка более крупной клады в большей степени, чем другие группы, и отделена от этого предка на такое же время, как и все другие существующие группы. Однако бывают случаи, когда необычно маленький размер сестринской группы действительно коррелирует с необычным количеством наследственных признаков, как у Amborella (см. Ниже). Вероятно, это является источником неправильного использования этого термина. Другими известными примерами этого явления являются яйцекладущее без сосков размножение и лактация у однопроходных , клады млекопитающих. [ 11 ] всего пять видов и архаичная анатомия туатары , [ 12 ] базальная клада лепидозавров с одним видом.
Примеры
[ редактировать ]Цветущие растения
[ редактировать ]
Семейство цветковых растений Amborellaceae , распространенное в Новой Каледонии в юго-западной части Тихого океана. [ n 9 ] представляет собой базальную кладу современных покрытосеменных растений . [ 13 ] состоит из большинства видов, родов, семейств и отрядов внутри группы, родственных всем другим покрытосеменным (из общего числа около 250 000 видов покрытосеменных). Считается, что черты Amborella trichopoda дают важное представление об эволюции цветковых растений; например, у него есть «самая примитивная древесина (состоящая только из трахеид ) среди всех живых покрытосеменных растений», а также «простые отдельные части цветка в неопределенном количестве и незапечатанные плодолистики». [ 14 ] Однако эти черты представляют собой смесь архаичных и апоморфных (производных) особенностей, которые удалось выявить только путем сравнения с другими покрытосеменными и их положением на филогенетическом дереве (летопись окаменелостей потенциально также могла бы быть полезной в этом отношении, но она отсутствует в этот случай). [ 14 ] Приведенная ниже кладограмма основана на Рамирес-Бараона и др. (2020), [ 15 ] с подсчетом видов, взятым из указанного источника.
|
Великие обезьяны
[ редактировать ]Этот раздел включает список общих ссылок , но в нем отсутствуют достаточные соответствующие встроенные цитаты . ( Ноябрь 2021 г. ) |
Среди обезьян человекообразных гориллы (восточные и западные) являются родственной группой шимпанзе , бонобо и человека . Эти пять видов образуют кладу, подсемейство Homininae (африканские обезьяны), базальным родом которого гориллу называют . Однако, если анализ не ограничивается родами, клада Homo plus Pan также является базальной.

| Люди |
| ||||||||||||
Более того, орангутанги являются сестринской группой Homininae и являются базальным родом семейства человекообразных обезьян Hominidae в целом.
| Гоминиды |
| ||||||
Подсемейства Homininae и Ponginae являются базальными внутри Hominidae, но, учитывая, что в кладограмме нет небазальных подсемейств, маловероятно, что этот термин будет применяться к любому из них. В общем, утверждение о том, что одна группа (например, орангутанги) является базовой или первой разветвляется внутри другой группы (например, Hominidae), может не иметь смысла, если только соответствующий таксономический уровень(и) (в данном случае род) ) указано. Если этот уровень невозможно указать (т. е. если рассматриваемая клада не имеет ранга), может потребоваться более подробное описание соответствующих сестринских групп. Как можно видеть, этот термин не отражает наследственные состояния или близость к общему предку современных видов.
В этом примере орангутанги отличаются от других родов своим азиатским ареалом. Этот факт, а также их базальный статус намекают на то, что самый недавний общий предок современных человекообразных обезьян, возможно, был евразийцем (см. ниже), и это предположение согласуется с другими свидетельствами. [ 19 ] (Конечно, малые обезьяны полностью азиатские.) Однако орангутанги также отличаются от африканских обезьян более древесным образом жизни - черта, которая обычно считается у обезьян наследственной. [ 20 ] [ 21 ]
Актуальность для биогеографической истории
[ редактировать ]Учитывая, что самый глубокий филогенетический раскол в группе, вероятно, произошел в начале ее истории, идентификация наиболее базального субклада (ов) в широко распространенном таксоне или кладе может дать ценную информацию о регионе его происхождения; однако отсутствие дополнительных видов в кладе не является свидетельством того, что она несет предковое состояние по большинству признаков. Наиболее обманчиво то, что люди часто полагают, что можно также предположить направление миграции в сторону от места происхождения (как в случаях с Amaurobioides и Noctilionoidea ниже). Как и в случае со всеми другими признаками, филогеографическое расположение одной клады, которая соединяется с корнем, не дает информации о предковом состоянии. Примеры, когда могли быть сделаны такие необоснованные выводы, включают:
- Пауки рода Amaurobioides обитают в Южной Африке, Австралии, Новой Зеландии и Чили. [ 22 ] [ 23 ] Самая базальная клада - южноафриканская; Данные последовательности ДНК указывают на то, что после того, как их южноамериканские предки достигли Южной Африки, они рассеялись на восток вплоть до Южной Америки в течение примерно 8 миллионов лет. [ 23 ]
- Ящерицы -игуаниды ( sensu lato ) распространены по всей Америке, на Мадагаскаре , а также на Фиджи и Тонге в западной части южной части Тихого океана. Ранее считалось, что малагасийские формы (Opluridae) являются базальными, с предполагаемой датой отклонения от остальных примерно 162 миллиона лет, незадолго до момента отделения Мадагаскара от Африки. [ 24 ] Это позволило предположить, что игуаниды когда-то имели широкое распространение в Гондване ; после того, как малагасийские и новосветские представители были разделены викариантностью , менее изолированные игуаниды Старого Света вымерли в результате конкуренции с другими группами ящериц (например, агамидами ). Напротив, игуаниды западной части Тихого океана глубоко гнездятся внутри американских игуанид. [ 25 ] очевидно, они колонизировали свой изолированный ареал после эпического рафтинга длиной 10 000 км . [ 26 ] [ 27 ] Однако исследование 2022 года показало, что оплюриды тесно связаны с американскими игуанами Leiosauridae , которые разошлись всего 60 миллионов лет назад после вероятного собственного рафтинга. [ 28 ] В связи с этим ни одна из «игуанид» Старого Света не считается базовой линией.
- Коралловые змеи насчитывают около 16 видов в Азии и более 65 видов в Америке. Однако ни одна из американских клад не является базальной, а это означает, что предки группы происходили из Старого Света. [ 29 ]
- Сохранившиеся австралидельфийские сумчатые насчитывают около 240 видов в Австралазии и один вид ( monito del monte ) в Южной Америке. Тот факт, что monito del monte занимает базальное положение (самый базальный вид, род, семейство и отряд) в надотряде Australidelphia, является важным ключом к тому, что его происхождение произошло из Южной Америки. Этот вывод согласуется с тем фактом, что южноамериканский отряд Didelphimorphia является базальным внутри инфракласса Marsupialia ; т.е. современные сумчатые в целом, по-видимому, также произошли из Южной Америки. [ 18 ] [ n 10 ] [ n 11 ] [ n 12 ]
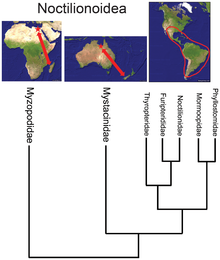
- В то время как надсемейство летучих мышей Noctilionoidea насчитывает более 200 видов в Неотропах, два в Новой Зеландии и два на Мадагаскаре , базальное положение малагасийского семейства [ 36 ] в сочетании с летописью окаменелостей и следующим по базальному расположению новозеландской семьи предполагает, что это надсемейство зародилось в Африке, а затем мигрировало на восток, в Южную Америку, размножаясь там, но выживая в Старом Свете только в рефугиумах . [ 37 ]
- Род Urocyon (серая и островная лисицы) является базальным в подсемействе собачьих . [ 38 ] что позволяет предположить североамериканское происхождение этой почти всемирной группы. Это согласуется с ископаемыми свидетельствами, указывающими на североамериканское происхождение семейства псовых в целом (два других подсемейства псовых, вымершие Borophaginae [ 39 ] и геспероционины , [ 40 ] последние, будучи базальными у Canidae, оба были эндемиками Северной Америки).
Примечания
[ редактировать ]- ^ Имеется в виду низшие таксономические ранги соответствующих клад; высшие ранги должны быть одинаковыми (при условии, что они имеют рейтинг).
- ^ См Amborella . пример базальная клада представляет собой единственный сохранившийся вид (который также является единственным живым представителем отряда Amborellales ) , в котором одна . Между тем, другая (без ранга) сестринская базальная клада насчитывает около 250 000 видов.
- ^ Например, C может быть родом , а другие базальные клады могут иметь более высокие ранги подсемейства или семейства .
- ^ В примере с человекообразными обезьянами горилла является базальным родом подсемейства Homininae, а понго — базальным родом семейства Hominidae. Две базальные клады последнего имеют высший ранг подсемейства, т. е. Homininae и Ponginae.
- ^ Большая диверсификация клады также может быть связана с колонизацией нового земельного массива, особенно если он больше или менее конкурентоспособен, чем исконный земельный массив; см. примеры коралловых змей, сумчатых и ночных летучих мышей .
- ^ Например, гигантская панда представляет собой наиболее основной из существующих видов, родов и подсемейств Ursidae . [ 9 ] но его специализация на бамбуковой диете не является наследственным характером урсид. [ 10 ]
- ^ То есть на диаграмме ниже обе базальные клады №1 и №2 являются более базальными, чем небазальная клада №1, которая, в свою очередь, более базальная, чем небазальные клады №2 и №3.
- ^ Поскольку родословная представляет собой линейную цепочку происхождения, все родословные внутри клады можно проследить не только до корня, но и до происхождения жизни . Таким образом, с филогенетической точки зрения представление о том, что линия является базовой, бессмысленно. Однако в генетике базальная линия относится к линии, соединяющей общего предка с одним вариантным аллелем с предком ветви с двумя вариантами потомков.
- ^ Новая Каледония рассматривается как рефугиум ; т. е. в этом случае считается, что географическое положение базальной клады не свидетельствует о месте происхождения покрытосеменных.
- ^ Эти выводы были подтверждены находкой эоценовых ископаемых останков микробиотерийского вида Woodburnodon Casei в Антарктиде . [ 30 ] который, предположительно, служил промежуточной станцией на пути миграции в Австралию перед окончательным распадом Гондваны .
- ^ Точно так же среди австралобатрахических лягушек южноамериканское семейство Calyptocephalellidae с 5 современными видами (живущими в том же вальдивском лесу, что и monito del monte) является базальным для австралийских семейств Limnodynastidae и Myobatrachidae , [ 31 ] насчитывает около 120 существующих видов, что позволяет предположить южноамериканское происхождение группы. [ 32 ] Это согласуется с находкой окаменелости южноамериканского семейства в Антарктиде. [ 33 ]
- ^ Ратиты , возможно, также путешествовали по суше из Южной Америки, чтобы колонизировать Австралию; [ 34 ] ископаемый бескилевой известен из Антарктиды, [ 35 ] и южноамериканские реи являются более базальными внутри группы, чем австралийско-тихоокеанские бескилевые. [ 34 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Смит, Стейси (19 сентября 2016 г.). «Из любви к деревьям: Предков нет среди нас» . Из любви к деревьям . Проверено 7 марта 2022 г.
- ^ Перейти обратно: а б с Крисп, Майкл Д.; Кук, Лин Дж. (2005). «Означают ли ранние ветвящиеся линии наследственные черты?» . Тенденции в экологии и эволюции . 20 (3): 122–128. дои : 10.1111/j.0307-6970.2004.00262.x . ПМИД 16701355 . S2CID 82371239 .
- ^ Перейти обратно: а б Крелл, Фрэнк-Т; Крэнстон, Питер С. (2004). «Какая сторона дерева более базальная?». Систематическая энтомология . 29 (3): 279–281. дои : 10.1016/j.tree.2004.11.010 . ПМИД 16701355 . S2CID 12285373 .
- ^ Слышал, СБ; Хаузер, Д.Л. (1995). «Ключевые эволюционные инновации и их экологические механизмы». Историческая биология . 10 (2): 151–173. дои : 10.1080/10292389509380518 .
- ^ Энгель, М.С.; Гримальди, Д.А. (2004). «Новый свет, пролитый на старейшее насекомое». Природа . 427 (6975): 627–630. Бибкод : 2004Natur.427..627E . дои : 10.1038/nature02291 . ПМИД 14961119 . S2CID 4431205 .
- ^ Фернандес-Масуэкос, М.; Бланко-Пастор, JL; Хуан, А.; Карнисеро, П.; Форрест, А.; Аларкон, М.; Варгас, П.; Гловер, Би Джей (2019). «Макроэволюционная динамика нектарных шпор, ключевое эволюционное нововведение». Новый фитолог . 222 (2): 1123–1138. дои : 10.1111/nph.15654 . hdl : 10045/89954 . ПМИД 30570752 . S2CID 58567195 .
- ^ Баум, Д.А. (4 ноября 2013 г.). «Филогенетика и история жизни» . Принстонский путеводитель по эволюции . Издательство Принстонского университета. п. 57. ИСБН 978-1-4008-4806-5 . ОСЛК 861200134 .
- ^ Перейти обратно: а б Дженнер, Рональд А. (2006). «Освобождение эво-дево: наследственные влечения, модельные организмы и базальная чушь». Гены развития и эволюция . 216 (7–8): 385–394. дои : 10.1007/s00427-006-0084-5 . ПМИД 16733736 . S2CID 23140594 .
- ^ Краузе, Дж.; Унгер, Т.; Носон, А.; Маласпинас, А.; Колокотронис, С.; Стиллер, М.; Сойбельзон, Л.; Сприггс, Х.; Дорогой, ПХ; Бриггс, AW; Брей, SCE; О'Брайен, SJ; Рабедер, Г.; Матеус, П.; Купер, А.; Слаткин М.; Паабо, С.; Хофрейтер, М. (2008). «Митохондриальные геномы обнаруживают взрывную радиацию вымерших и современных медведей вблизи границы миоцена и плиоцена» . Эволюционная биология BMC . 8 (220): 220. дои : 10.1186/1471-2148-8-220 . ПМК 2518930 . ПМИД 18662376 .
- ^ Маклеллан, Брюс; Райнер, Дэвид К. (1994). «Обзор эволюции медведей». Медведи: их биология и управление . 9 (1): 85–96. дои : 10.2307/3872687 . JSTOR 3872687 .
- ^ Ван Рид, Теун (2005). «Утконос на своем месте: ядерные гены и индели подтверждают родство сестринской группы однопроходных и терианцев» . Молекулярная биология и эволюция . 23 (3): 587–597. дои : 10.1093/molbev/msj064 . ПМИД 16291999 .
- ^ Эррера-Флорес, Дж.А.; Стаббс, ТЛ; Бентон, MJ; Рута, М. (2017). «Макроэволюционные закономерности ринхоцефалии: является ли туатара ( Sphenodon punctatus ) живым ископаемым?» . Палеонтология . 60 (3): 319–328. Бибкод : 2017Palgy..60..319H . дои : 10.1111/пала.12284 .
- ^ Ли, Х.-Т.; Йи, Т.-С.; Гао, Л.-М.; Ма, П.-Ф.; Чжан, Т.; Ян, Ж.-Б.; Гитценданнер, Массачусетс; Фрич, П.В.; Кай, Дж.; Луо, Ю.; Ван, Х.; ван дер Банк, М.; Чжан, С.-Д.; Ван, Ц.-Ф.; Ван, Дж.; Чжан, З.-Р.; Фу, К.-Н.; Ян, Дж.; Холлингсворт, премьер-министр; Чейз, штат Вашингтон; Солтис, Делавэр; Солтис, П.С.; Ли, Д.-З. (2019). «Происхождение покрытосеменных растений и загадка юрского разрыва». Природные растения . 5 (5): 461–470. дои : 10.1038/s41477-019-0421-0 . ПМИД 31061536 . S2CID 146118264 .
- ^ Перейти обратно: а б Эссиг, ФБ (1 июля 2014 г.). «Что такого примитивного в Амборелле ?» . Профессор ботаники . Проверено 4 октября 2014 г.
- ^ Рамирес-Бараона, С.; Соке, Х.; Магальон, С. (2020). «Замедленная и географически неоднородная диверсификация семейств цветковых растений». Экология и эволюция природы . 4 (9): 1232–1238. дои : 10.1038/s41559-020-1241-3 . ПМИД 32632260 . S2CID 220375828 .
- ^ Перейти обратно: а б с д и Палмер, доктор медицинских наук; Солтис, Делавэр; Чейз, М.В. (2004). «Растительное древо жизни: обзор и некоторые точки зрения» . Американский журнал ботаники . 91 (10): 1437–1445. дои : 10.3732/ajb.91.10.1437 . ПМИД 21652302 .
- ^ Перейти обратно: а б с Кристенхуш, MJM; Бинг, JW (2016). «Число известных видов растений в мире и его ежегодное увеличение» . Фитотаксы . 261 (3): 201–217. дои : 10.11646/phytotaxa.261.3.1 .
- ^ Перейти обратно: а б Нильссон, Массачусетс; Чураков Г.; Соммер, М.; Ван Тран, Н.; Земанн, А.; Брозиус, Дж.; Шмитц, Дж. (27 июля 2010 г.). Пенни, Дэвид (ред.). «Отслеживание эволюции сумчатых с помощью вставок архаичных геномных ретропозонов» . ПЛОС Биология . 8 (7): e1000436. дои : 10.1371/journal.pbio.1000436 . ПМК 2910653 . ПМИД 20668664 .
- ^ Моя-Сола, С.; Альба, DM; Альмеция, С.; Казановас-Вилар, И.; Колер, М.; Де Эстебан-Тривиньо, С.; Роблес, Дж. М.; Галиндо, Дж.; Фортуни, Дж. (16 июня 2009 г.). «Уникальный европейский гуманоид среднего миоцена и происхождение великих обезьян и людей» . ПНАС . 106 (24): 9601–9606. Бибкод : 2009PNAS..106.9601M . дои : 10.1073/pnas.0811730106 . ПМК 2701031 . ПМИД 19487676 . .
- ^ МаКлатчи, Л.; Гебо, Д.; Китио, Р.; Пилбим, Д. (2000). «Посткраниальная функциональная морфология Morotopithecus Bishopi , имеющая значение для эволюции передвижения современных обезьян». Журнал эволюции человека . 39 (2): 159–183. дои : 10.1006/jhev.2000.0407 . ПМИД 10968927 .
- ^ Торп, СКС; Кромптон, Р.Х. (2006). «Позиционное поведение орангутанга и характер древесного передвижения Hominoidea» . Американский журнал физической антропологии . 131 (3): 384–401. дои : 10.1002/ajpa.20422 . ПМИД 16617429 .
- ^ Куксо, Ф. (08 ноября 2016 г.). «Мореходные пауки облетели весь мир за 8 миллионов лет» . Научный американец . Проверено 10 ноября 2016 г.
- ^ Перейти обратно: а б Кунтнер, М.; Чеккарелли, Ф.С.; Опель, Б.Д.; Хаддад, ЧР; Рэйвен, Роберт Дж.; Сото, Э.М.; Рамирес, MJ (12 октября 2016 г.). «Вокруг света за восемь миллионов лет: историческая биогеография и эволюция паука зоны распыления Amaurobioides (Araneae: Anyphaenidae)» . ПЛОС ОДИН . 11 (10): e0163740. Бибкод : 2016PLoSO..1163740C . дои : 10.1371/journal.pone.0163740 . ПМК 5061358 . ПМИД 27732621 .
- ^ Окадзима, Ю.; Кумадзава, Ю. (15 июля 2009 г.). «Митогеномные взгляды на филогению и биогеографию игуанид: гондванское замещение происхождения мадагаскарских оплюринов». Джин . 441 (1–2): 28–35. дои : 10.1016/j.gene.2008.06.011 . ПМИД 18598742 .
- ^ Шульте, Дж. А.; Валладарес, Япония; Ларсон, А. (сентябрь 2003 г.). «Филогенетические отношения внутри игуанид, выявленные с использованием молекулярных и морфологических данных и филогенетической таксономии игуанских ящериц». Герпетологика . 59 (3): 399–419. дои : 10.1655/02-48 . S2CID 56054202 .
- ^ Гиббонс, JRH (31 июля 1981 г.). «Биогеография Brachylophus (Iguanidae), включая описание нового вида B. vitiensis с Фиджи». Журнал герпетологии . 15 (3): 255–273. дои : 10.2307/1563429 . JSTOR 1563429 .
- ^ Кио, Дж.С.; Эдвардс, Д.Л.; Фишер, Р.Н.; Харлоу, PS (2008). «Молекулярный и морфологический анализ находящихся под угрозой исчезновения фиджийских игуан раскрывает загадочное разнообразие и сложную биогеографическую историю» . Философские труды Королевского общества B: Биологические науки . 363 (1508): 3413–3426. дои : 10.1098/rstb.2008.0120 . ПМК 2607380 . ПМИД 18782726 .
- ^ Вельт, Рэйчел С.; Раксуорси, Кристофер Дж. (01 февраля 2022 г.). «Рассредоточение, а не наместничество объясняет биогеографическое происхождение игуан на Мадагаскаре» . Молекулярная филогенетика и эволюция . 167 : 107345. doi : 10.1016/j.ympev.2021.107345 . ISSN 1055-7903 . PMID 34748875 . S2CID 243821392 .
- ^ Словински, Дж. Б. ; Баунди, Дж.; Лоусон, Р. (июнь 2001 г.). «Филогенетические отношения азиатских коралловых змей (Elapidae: Calliophis и Maticora) на основе морфологических и молекулярных признаков». Герпетологика . 57 (2): 233–245. JSTOR 3893186 .
- ^ Гоин, Ф.Дж.; Зимич, Н.; Регеро, Массачусетс; Сантильяна, Сан-Франциско; Маренси, ЮАР; Моли, Джей Джей (2007). «Новые сумчатые (млекопитающие) из эоцена Антарктиды, а также происхождение и родство микробиотерий » Журнал Аргентинской геологической ассоциации . 62 (4): 597–603 . Получено 1 июля 2016 г.
- ^ Пайрон, РА; Винс, Джей-Джей (2011). «Крупномасштабная филогения земноводных, включающая более 2800 видов, и пересмотренная классификация современных лягушек, саламандр и червяг» . Молекулярная филогенетика и эволюция . 61 (2): 543–583. дои : 10.1016/j.ympev.2011.06.012 . ПМИД 21723399 .
- ^ Фэн, Ю.-Дж.; Блэкберн, округ Колумбия; Лян, Д.; Хиллис, DM; Уэйк, ДБ; Каннателла, округ Колумбия; Чжан, П. (2017). «Филогеномика обнаруживает быструю и одновременную диверсификацию трех основных клад гондванских лягушек на границе мела и палеогена» . Труды Национальной академии наук . 114 (29): E5864–E5870. Бибкод : 2017PNAS..114E5864F . дои : 10.1073/pnas.1704632114 . ПМК 5530686 . ПМИД 28673970 .
- ^ Мёрс, Т.; Регеро, М.; Васильян, Д. (2020). «Первая ископаемая лягушка из Антарктиды: последствия для климатических условий эоцена в высоких широтах и гондванского космополитизма Австралобатрахии» . Научные отчеты . 10 (1): 5051. Бибкод : 2020НатСР..10.5051М . дои : 10.1038/s41598-020-61973-5 . ПМК 7181706 . ПМИД 32327670 .
- ^ Перейти обратно: а б Ёнедзава, Т.; Сегава, Т.; Мори, Х.; Кампос, ПФ; Хонго, Ю.; Эндо, Х.; Акиёси, А.; Кохно, Н.; Нисида, С.; Ву, Дж.; Джин, Х.; Адачи, Дж.; Кишино, Х.; Курокава, К.; Ноги, Ю.; Танабэ, Х.; Мукояма, Х.; Ёсида, К.; Расоамиараманана, А.; Ямагиши, С.; Хаяши, Ю.; Ёсида, А.; Койке, Х.; Акисинономия, Ф.; Виллерслев, Э.; Хасэгава, М. (15 декабря 2016 г.). «Филогеномика и морфология вымерших палеогнат раскрывают происхождение и эволюцию бескилевых» . Современная биология . 27 (1): 68–77. дои : 10.1016/j.cub.2016.10.029 . ПМИД 27989673 .
- ^ Тамбусси, CP; Норьега, Дж.И.; Газджицкий А.; Татур, А.; Регеро, Массачусетс; Вискаино, Сан-Франциско (1994). «Ратитовая птица из палеогеновой формации Ла-Плато, остров Сеймур, Антарктида» (PDF ) Польские полярные исследования . 15 (1–2): 15–2 Архивировано из оригинала (PDF) 2 декабря 2019 г. Получено 28 декабря.
- ^ Тилинг, ЕС; Спрингер, М.; Мэдсен, О.; Бейтс, П.; О'Брайен, С.; Мерфи, В. (28 января 2005 г.). «Молекулярная филогения летучих мышей освещает биогеографию и летопись окаменелостей». Наука . 307 (5709): 580–584. Бибкод : 2005Sci...307..580T . дои : 10.1126/science.1105113 . ПМИД 15681385 . S2CID 25912333 .
- ^ Ганнелл, Г.Ф.; Симмонс, Северная Каролина; Зайферт, ER (04 февраля 2014 г.). «Новые Myzopodidae (Chiroptera) из позднего палеогена Египта: уточненный диагноз семейства и биогеографическое происхождение Noctilionoidea» . ПЛОС ОДИН . 9 (2): е86712. Бибкод : 2014PLoSO...986712G . дои : 10.1371/journal.pone.0086712 . ПМЦ 3913578 . ПМИД 24504061 .
- ^ Линдблад-То, К.; Уэйд, CM; Миккельсен, Т.С.; Карлссон, ЕК; Яффе, Д.Б.; Камаль, М.; Клэмп, М.; Чанг, Дж.Л.; Кулбокас, Э.Дж.; Зоди, MC; Маусели, Э.; Се, X.; Брин, М.; Уэйн, РК; Острандер, Э.А.; Понтинг, КП; Галиберт, Ф.; Смит, доктор медицинских наук; Деджонг, ПиДжей; Киркнесс, Э.; Альварес, П.; Бьяджи, Т.; Брокман, В.; Батлер, Дж.; Чин, К.-В.; Кук, А.; Кафф, Дж.; Дейли, MJ; Декаприо, Д.; и др. (2005). «Последовательность генома, сравнительный анализ и структура гаплотипов домашней собаки» . Природа . 438 (7069): 803–819. Бибкод : 2005Natur.438..803L . дои : 10.1038/nature04338 . ПМИД 16341006 .
- ^ Ван, Сяомин; Тедфорд, Ричард; Тейлор, Берил (17 ноября 1999 г.). «Филогенетическая систематика Borophaginae (Carnivora, Canidae)». Бюллетень Американского музея естественной истории . 243 . hdl : 2246/1588 .
- ^ Ван, X.; Тедфорд, Р.Х. (2008). Собаки, их ископаемые родственники и история эволюции . Издательство Колумбийского университета. стр. 23–31. ISBN 978-0-231-13528-3 .
Внешние ссылки
[ редактировать ]- «Интерпретация древовидной диаграммы или списка подгрупп на странице Древа жизни» . Веб-проект «Древо жизни» .
| Соответствующие поля | ||
|---|---|---|
| Основные понятия | ||
| Методы вывода | ||
| Текущие темы | ||
| Групповые черты | ||
| Типы групп | ||
| Номенклатура | ||