Нейронное кодирование звука
Эта статья нуждается в дополнительных цитатах для проверки . ( январь 2023 г. ) |
| Часть серии о | ||||||
| Фонетика | ||||||
|---|---|---|---|---|---|---|
| Часть серии «Лингвистика». | ||||||
| Субдисциплины | ||||||
| Артикуляция | ||||||
| ||||||
| Акустика | ||||||
| ||||||
| Восприятие | ||||||
| ||||||
| Лингвистический портал | ||||||
Нейронное кодирование звука представляет собой представление слуховых ощущений и восприятия в нервной системе . [1] Сложности современной нейробиологии постоянно пересматриваются. Таким образом, то, что известно о слуховой системе, постоянно меняется. Кодирование звуков включает преобразование звуковых волн в электрические импульсы ( потенциалы действия ) по волокнам слухового нерва и дальнейшую обработку в мозге.
Базовая физика звука
[ редактировать ]Звуковые волны — это то, что физики называют продольными волнами , которые состоят из распространяющихся областей высокого давления (сжатия) и соответствующих областей низкого давления (разрежения).
Форма волны
[ редактировать ]Форма волны — это описание общей формы звуковой волны. Формы сигналов иногда описываются суммой синусоид с помощью анализа Фурье .
Амплитуда
[ редактировать ]
Амплитуда представляет собой величину (величину) изменений давления в звуковой волне и в первую очередь определяет громкость, с которой воспринимается звук. В синусоидальной функции, такой как , C представляет собой амплитуду звуковой волны.

Частота и длина волны
[ редактировать ]Частота звука определяется как количество повторений его формы волны в секунду и измеряется в герцах ; частота обратно пропорциональна длине волны (в среде с одинаковой скоростью распространения, например звук в воздухе). Длина волны звука — это расстояние между любыми двумя последовательными совпадающими точками на форме волны. Диапазон слышимых частот для молодых людей составляет от 20 Гц до 20 кГц. Слышимость более высоких частот снижается с возрастом, ограничиваясь примерно 16 кГц для взрослых и даже 3 кГц для пожилых людей. [ нужна ссылка ]
Анатомия уха
[ редактировать ]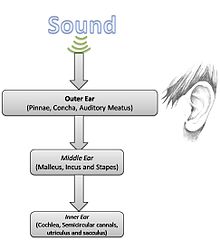
Учитывая простую физику звука, анатомию и физиологию слуха можно изучить более подробно.
Наружное ухо
[ редактировать ]Наружное ухо состоит из ушной раковины или ушной раковины (видимых частей, включая мочки и раковины) и слухового прохода (прохода для звука). Основная функция этой части уха — собирать звуковую энергию и доставлять ее к барабанной перепонке . Резонансы наружного уха избирательно усиливают звуковое давление с частотой в диапазоне 2–5 кГц. [2]
Благодаря своей асимметричной структуре ушная раковина способна давать дополнительные подсказки о высоте, с которой произошел звук. Вертикальная асимметрия ушной раковины выборочно усиливает звуки более высокой частоты с большой высоты, обеспечивая тем самым пространственную информацию благодаря своей механической конструкции. [2] [3]
Среднее ухо
[ редактировать ]
Среднее ухо играет решающую роль в слуховом процессе, поскольку оно, по сути, преобразует изменения давления воздуха в возмущения жидкостей внутреннего уха. Другими словами, именно механическая передаточная функция позволяет эффективно передавать собранную звуковую энергию между двумя разными средами. [2] За этот сложный процесс отвечают три маленькие кости: молоточек , наковальня и стремечко , известные под общим названием ушные косточки . [4] [5] Согласование импеданса осуществляется посредством соотношения рычагов и соотношения площадей барабанной перепонки и подножия стремени, создавая механизм, подобный трансформатору . [4] Кроме того, косточки устроены таким образом, чтобы резонировать на частоте 700–800 Гц, одновременно защищая внутреннее ухо от чрезмерной энергии. [5] Определенная степень нисходящего контроля присутствует на уровне среднего уха, главным образом, за счет двух мышц, присутствующих в этой анатомической области: напрятеля барабанной перепонки и стремени . Эти две мышцы могут удерживать косточки, чтобы уменьшить количество энергии, передаваемой во внутреннее ухо в громкой обстановке. [3] [4]
Внутреннее ухо
[ редактировать ]
Улитка . внутреннего уха, чудо физиологической инженерии, действует как частотный анализатор и нелинейный акустический усилитель [2] Улитка имеет более 32 000 волосковых клеток . Наружные волосковые клетки в первую очередь обеспечивают усиление бегущих волн, вызываемых звуковой энергией, тогда как внутренние волосковые клетки обнаруживают движение этих волн и возбуждают нейроны (типа I) слухового нерва .
Базальный конец улитки, куда звуки поступают из среднего уха, кодирует верхний конец диапазона слышимых частот, а апикальный конец улитки кодирует нижний конец диапазона частот. Эта тонотопия играет решающую роль в слухе, поскольку позволяет осуществлять спектральное разделение звуков. Поперечный разрез улитки покажет анатомическую структуру с тремя основными камерами ( лестница преддверия , средняя лестница и барабанная лестница ). [5] На верхушечном конце улитки, в отверстии, известном как геликотрема, лестница преддверия сливается с барабанной лестницей. Жидкость, находящаяся в этих двух камерах улитки, представляет собой перилимфу , а средняя лестница, или улитковый проток , заполнена эндолимфой . [3]
Трансдукция
[ редактировать ]Слуховые волосковые клетки
[ редактировать ]Слуховые волосковые клетки улитки лежат в основе особых функций слуховой системы (аналогичные волосковые клетки расположены в полукружных каналах ). Их основная функция — механотрансдукция или преобразование механических и нервных сигналов. Относительно небольшое количество слуховых волосковых клеток удивляет по сравнению с другими сенсорными клетками, такими как палочки и колбочки зрительной системы . Таким образом, потеря меньшего количества (порядка тысяч) слуховых волосковых клеток может иметь разрушительные последствия, в то время как потеря большего количества клеток сетчатки (порядка сотен тысяч) не будет такой страшной с сенсорной точки зрения. [6]
Волосковые клетки улитки организованы как внутренние волосковые клетки и внешние волосковые клетки; внутренний и внешний относятся к относительному положению относительно оси улитковой спирали. Внутренние волосковые клетки являются первичными сенсорными рецепторами, и значительная часть сенсорной информации, поступающей в слуховую кору, поступает от этих волосковых клеток. С другой стороны, наружные волосковые клетки усиливают механический сигнал, используя электромеханическую обратную связь. [6]
Механотрансдукция
[ редактировать ]Апикальная поверхность каждой волосковой клетки улитки содержит пучок волос . Каждый пучок волос содержит около 300 тонких выступов, известных как стереоцилии , образованных актиновыми элементами цитоскелета. [7] Стереоцилии в пучке волос расположены в несколько рядов разной высоты. Помимо стереоцилий, существует настоящая цилиарная структура, известная как киноцилия , которая, как полагают, играет роль в дегенерации волосковых клеток, вызванной воздействием высоких частот. [2] [7]
Стереоцилия способна изгибаться в месте прикрепления к апикальной поверхности волосковой клетки. Актиновые фибрином нити, образующие ядро стереоцилии, сильно взаимосвязаны и перекрестно связаны с и поэтому являются жесткими и негибкими в положениях, отличных от основания. Когда стереоцилии в самом высоком ряду отклоняются в направлении положительного стимула, более короткие ряды стереоцилий также отклоняются. [7] Эти одновременные отклонения происходят из-за нитей, называемых кончиковыми звеньями, которые прикрепляют сторону каждой более высокой стереоцилии к вершине более короткой стереоцилии в соседнем ряду. Когда самые высокие стереоцилии отклоняются, в кончиковых звеньях возникает напряжение, вызывающее также отклонение стереоцилий в других рядах. На нижнем конце каждого кончика звена находится один или несколько каналов механо-электрической трансдукции (МЕТ), которые открываются под действием напряжения в кончиках звеньев. [8] Эти МЕТ-каналы представляют собой катион-селективные каналы трансдукции, которые позволяют ионам калия и кальция проникать в волосковую клетку из эндолимфы, омывающей ее апикальный конец.
Приток катионов, особенно калия, через открытые каналы МЕТ вызывает деполяризацию мембранного потенциала волосковой клетки. Эта деполяризация открывает потенциалзависимые кальциевые каналы, обеспечивая дальнейший приток кальция. Это приводит к увеличению концентрации кальция, что запускает экзоцитоз везикул нейромедиаторов в ленточных синапсах на базолатеральной поверхности волосковой клетки. Высвобождение нейромедиатора в ленточном синапсе, в свою очередь, генерирует потенциал действия в соединенном с ним волокне слухового нерва. [7] Важна также гиперполяризация волосковой клетки, возникающая при выходе калия из клетки, поскольку она останавливает приток кальция и, следовательно, останавливает слияние везикул в ленточных синапсах. Таким образом, как и в других частях тела, трансдукция зависит от концентрации и распределения ионов. [7] Перилимфа, обнаруженная в барабанной лестнице, имеет низкую концентрацию калия, тогда как эндолимфа, обнаруженная в средней лестнице, имеет высокую концентрацию калия и электрический потенциал около 80 милливольт по сравнению с перилимфой. [2] Механотрансдукция стереоцилиями очень чувствительна и способна обнаруживать даже малейшие возмущения, такие как колебания жидкости размером 0,3 нанометра, и может преобразовать эту механическую стимуляцию в электрический нервный импульс примерно за 10 микросекунд. [ нужна ссылка ]
Нервные волокна от улитки
[ редактировать ]обнаруживаются два типа афферентных нейронов В улитковом нерве : тип I и тип II. Каждый тип нейрона обладает определенной клеточной избирательностью в улитке. [9] Механизм, определяющий селективность каждого типа нейрона для конкретной волосковой клетки, был предложен двумя диаметрально противоположными теориями в нейробиологии, известными как гипотеза периферической инструкции и гипотеза автономной клеточной инструкции. Гипотеза периферической инструкции утверждает, что фенотипическая дифференциация между двумя нейронами не происходит до тех пор, пока эти недифференцированные нейроны не прикрепятся к волосковым клеткам, что, в свою очередь, будет определять путь дифференцировки. Гипотеза клеточной автономной инструкции утверждает, что дифференцировка в нейроны типа I и типа II происходит после последней фазы митотического деления, но предшествует иннервации. [9] Оба типа нейронов участвуют в кодировании звука для передачи в мозг.
Нейроны I типа
[ редактировать ]Нейроны I типа иннервируют внутренние волосковые клетки. Конвергенция этого типа нейронов к базальному концу значительно больше, чем к апикальному концу. [9] Пучок радиальных волокон действует как посредник между нейронами I типа и внутренними волосковыми клетками. Соотношение иннервации между нейронами типа I и внутренними волосковыми клетками составляет 1:1, что приводит к высокой точности и разрешению передачи сигнала. [9]
Нейроны II типа
[ редактировать ]С другой стороны, нейроны типа II иннервируют внешние волосковые клетки. Однако наблюдается значительно большая конвергенция этого типа нейронов к вершинному концу по сравнению с базальным концом. Соотношение иннервации между нейронами типа II и внешними волосковыми клетками составляет 1:30-60, что, в свою очередь, делает эти нейроны идеальными для электромеханической обратной связи. [9] Нейронами типа II можно физиологически манипулировать для иннервации внутренних волосковых клеток при условии, что внешние волосковые клетки были разрушены либо в результате механического повреждения, либо в результате химического повреждения, вызванного такими лекарствами, как гентамицин . [9]
Ствол мозга и средний мозг
[ редактировать ]Слуховая нервная система включает в себя множество этапов обработки информации между ухом и корой головного мозга .
Слуховая кора
[ редактировать ]Первичные слуховые нейроны переносят потенциалы действия из улитки в путь передачи, показанный на соседнем изображении. Несколько ретрансляционных станций действуют как центры интеграции и обработки. Сигналы достигают первого уровня корковой обработки в первичной слуховой коре (А1), в верхней височной извилине доли височной . [6] Большинство областей до A1 включительно картированы тонотопически (то есть частоты сохраняются в упорядоченном расположении). Однако A1 участвует в кодировании более сложных и абстрактных аспектов слуховых стимулов, не кодируя должным образом частотный состав, включая наличие отдельного звука или его отголосков. [10] Как и нижние области, эта область мозга имеет комбинационно-чувствительные нейроны, которые имеют нелинейные реакции на стимулы. [6]
Недавние исследования, проведенные на летучих мышах и других млекопитающих, показали, что способность обрабатывать и интерпретировать частотную модуляцию в первую очередь возникает в верхней и средней височных извилинах височной доли. [6] Латерализация функций мозга существует в коре головного мозга, при этом обработка речи происходит в левом полушарии головного мозга , а звуков окружающей среды — в правом полушарии слуховой коры. Музыка, оказывающая влияние на эмоции, также обрабатывается в правом полушарии слуховой коры. Хотя причина такой локализации не совсем понятна, латерализация в этом случае не подразумевает исключительности, поскольку в обработке участвуют оба полушария, но одно полушарие имеет тенденцию играть более значительную роль, чем другое. [6]
Последние идеи
[ редактировать ]- По мере продвижения по слуховой коре наблюдалось изменение механизмов кодирования. Кодирование отклоняется от синхронных ответов в ядре улитки и позже становится зависимым от скорости кодирования в нижних холмиках . [11]
- Несмотря на достижения в генной терапии, которые позволяют изменять экспрессию генов, влияющих на слух, таких как ATOH1 , и использование вирусных векторов для этой цели, микромеханические и нервные сложности, окружающие волосковые клетки внутреннего уха, искусственная регенерация in vitro остается далекой реальностью. [12]
- Недавние исследования показывают, что слуховая кора, возможно, не так вовлечена в обработку информации сверху вниз, как считалось ранее. В исследованиях, проведенных на приматах для решения задач, требующих распознавания акустического трепетания, Лемус обнаружил, что слуховая кора играет только сенсорную роль и не имеет никакого отношения к познанию поставленной задачи. [13]
- В связи с наличием тонотопических карт в слуховой коре в раннем возрасте предполагалось, что корковая реорганизация мало связана с формированием этих карт, но эти карты подвержены пластичности. [14] Судя по всему, кора выполняет более сложную обработку, чем спектральный анализ или даже спектрально-временной анализ. [10]
Ссылки
[ редактировать ]- ^ Леонард, Мэтью К.; Гвильямс, Лаура; Селлерс, Кристин К.; Чанг, Джейсон Э.; Сюй, Дуо; Мишлер, Гэвин; Месгарани, Нима; Велкенхейзен, Марлен; Дутта, Барундеб; Чанг, Эдвард Ф. (15 февраля 2024 г.). «Крупномасштабное кодирование речевого звука одним нейроном на глубине коры головного мозга человека» . Природа . 626 (7999): 593–602. дои : 10.1038/s41586-023-06839-2 . ISSN 0028-0836 . ПМЦ 10866713 .
- ^ Перейти обратно: а б с д и ж Хадспет, Эй.Дж. (октябрь 1989 г.). «Как работает ухо». Природа . 341 (6241): 397–404. Бибкод : 1989Natur.341..397H . дои : 10.1038/341397a0 . ПМИД 2677742 . S2CID 33117543 .
- ^ Перейти обратно: а б с Хадспет, Эй.Дж. (2001). «Как работает ухо: механоэлектрическая трансдукция и усиление волосковыми клетками внутреннего уха». Харви Лект . 97 : 41–54. ПМИД 14562516 .
- ^ Перейти обратно: а б с Худде, Х.; Вайстенхофер, К. (2006). «Основные особенности среднего уха человека». ORL J Оториноларингол Relat Spec . 68 (6): 324–328. дои : 10.1159/000095274 . PMID 17065824 . S2CID 42550955 .
- ^ Перейти обратно: а б с Хадспет, ЭйДжей; Кониси, М. (октябрь 2000 г.). «Слуховая нейробиология: развитие, трансдукция и интеграция» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (22): 11690–1. дои : 10.1073/pnas.97.22.11690 . ПМК 34336 . ПМИД 11050196 .
- ^ Перейти обратно: а б с д и ж Каас, Дж. Х.; Хакетт, штат Калифорния; Трамо, MJ. (апрель 1999 г.). «Слуховая обработка в коре головного мозга приматов» (PDF) . Современное мнение в нейробиологии . 9 (2): 164–170. дои : 10.1016/S0959-4388(99)80022-1 . ПМИД 10322185 . S2CID 22984374 .
- ^ Перейти обратно: а б с д и Феттиплейс, Р.; Хакни, CM. (январь 2006 г.). «Сенсорная и моторная роль слуховых волосковых клеток». Nat Rev Neurosci . 7 (1): 19–29. дои : 10.1038/nrn1828 . ПМИД 16371947 . S2CID 10155096 .
- ^ Берг, М.; Феттиплейс, Р.; Нам, Дж. Х.; Риччи, А.Дж. (май 2009 г.). «Локализация каналов механопреобразователей внутренних волосковых клеток с использованием высокоскоростной визуализации кальция» . Природная неврология . 12 (5): 553–558. дои : 10.1038/nn.2295 . ПМЦ 2712647 . ПМИД 19330002 .
- ^ Перейти обратно: а б с д и ж Рубель, EW.; Фрич, Б. (2002). «Развитие слуховой системы: первичные слуховые нейроны и их мишени». Ежегодный обзор неврологии . 25 : 51–101. дои : 10.1146/annurev.neuro.25.112701.142849 . ПМИД 12052904 .
- ^ Перейти обратно: а б Чечик, Гал; Нелькен (2012). «Слуховая абстракция от спектро-временных характеристик к кодированию слуховых объектов» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (44): 18968–73. Бибкод : 2012PNAS..10918968C . дои : 10.1073/pnas.1111242109 . ПМЦ 3503225 . ПМИД 23112145 .
- ^ Фризина, РД. (август 2001 г.). «Механизмы подкоркового нейронного кодирования слуховой временной обработки». Исследование слуха . 158 (1–2): 1–27. дои : 10.1016/S0378-5955(01)00296-9 . ПМИД 11506933 . S2CID 36727875 .
- ^ Бриганда, СП; Хеллер, С. (июнь 2009 г.). «Quo vadis, регенерация волосковых клеток?» . Природная неврология . 12 (6): 679–685. дои : 10.1038/nn.2311 . ПМК 2875075 . ПМИД 19471265 .
- ^ Лемус, Л.; Эрнандес, А.; Ромо, Р. (июнь 2009 г.). «Нейронные коды перцептивного распознавания акустического трепетания в слуховой коре приматов» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (23): 9471–9476. Бибкод : 2009PNAS..106.9471L . дои : 10.1073/pnas.0904066106 . ПМЦ 2684844 . ПМИД 19458263 .
- ^ Кандлер, К.; Клауз, А.; Но, Дж. (июнь 2009 г.). «Тонотопическая реорганизация развивающихся слуховых стволовых цепей» . Природная неврология . 12 (6): 711–7. дои : 10.1038/nn.2332 . ПМК 2780022 . ПМИД 19471270 .