Фоторецепторная клетка
| Фоторецепторная клетка | |
|---|---|
 | |
| Идентификаторы | |
| МеШ | D010786 |
| НейроЛекс ID | сао226523927 |
| ФМА | 85613 86740, 85613 |
| Анатомические термины нейроанатомии | |
Фоторецепторная клетка — это специализированный тип нейроэпителиальных клеток сетчатки , который способен осуществлять зрительную фототрансдукцию . Большое биологическое значение фоторецепторов заключается в том, что они преобразуют свет (видимое электромагнитное излучение ) в сигналы, способные стимулировать биологические процессы. Если быть более конкретным, белки-фоторецепторы в клетке поглощают фотоны клетки , вызывая изменение мембранного потенциала .
В настоящее время в глазах млекопитающих известно три типа фоторецепторных клеток: палочки , колбочки и внутренне светочувствительные ганглиозные клетки сетчатки . Двумя классическими фоторецепторными клетками являются палочки и колбочки, каждая из которых вносит информацию, используемую зрительной системой для формирования изображения окружающей среды, зрения . Палочки в первую очередь обеспечивают скотопическое зрение (тусклые условия), тогда как колбочки в первую очередь обеспечивают фотопическое зрение (яркие условия), но процессы в каждом из них, поддерживающие фототрансдукцию, аналогичны. [ 1 ] Внутренне светочувствительные ганглиозные клетки сетчатки были открыты в 1990-х годах. [ 2 ] Считается, что эти клетки не способствуют зрению напрямую, но играют роль в поддержании циркадного ритма и зрачкового рефлекса .
Светочувствительность
[ редактировать ]
Каждый фоторецептор поглощает свет в соответствии со своей спектральной чувствительностью (поглощением), которая определяется белками фоторецептора, экспрессируемыми в этой клетке. У людей есть три класса колбочек (L, M, S), каждый из которых отличается спектральной чувствительностью и «предпочитает» фотоны разной длины волны (см. график). Например, пиковая длина волны спектральной чувствительности S-конуса составляет примерно 420 нм (нанометры, мера длины волны), поэтому вероятность поглощения фотона при длине волны 420 нм выше, чем при любой другой длине волны. Свет с большей длиной волны также может дать такой же отклик от S-конуса, но для этого он должен быть ярче.
В соответствии с принципом унивариантности выходной сигнал фоторецептора пропорционален только количеству поглощенных фотонов. Фоторецепторы не могут измерять длину волны света, который они поглощают, и поэтому не определяют цвет самостоятельно. Скорее, это соотношение реакций трех типов колбочек, которые могут оценить длину волны и, следовательно, обеспечить цветовое зрение .
Гистология
[ редактировать ]![Анатомия палочки[4]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/bb/Rod%26Cone.jpg/179px-Rod%26Cone.jpg)

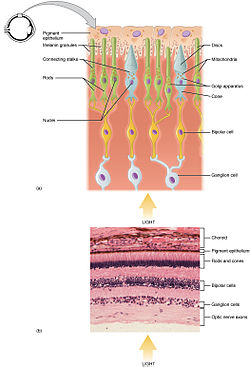
Фоторецепторы палочек и колбочек расположены на внешнем слое сетчатки ; они оба имеют одинаковую базовую структуру. Ближе всего к полю зрения (и дальше всего от мозга) находится окончание аксона , которое высвобождает нейротрансмиттер, называемый глутаматом, в биполярные клетки . Еще дальше находится тело клетки клетки , содержащее органеллы . Еще дальше находится внутренний сегмент, специализированная часть клетки , полная митохондрий . Основная функция внутреннего сегмента — обеспечение АТФ (энергии) для натриево-калиевого насоса . Наконец, ближе всего к мозгу (и дальше всего от поля зрения) находится внешний сегмент — часть фоторецептора, поглощающая свет . Внешние сегменты на самом деле представляют собой модифицированные реснички. [ 5 ] [ 6 ] которые содержат диски, заполненные опсином , молекулой, которая поглощает фотоны, а также потенциалзависимые натриевые каналы .
Мембранный фоторецепторный белок опсин содержит молекулу пигмента, называемую ретиналем . В палочках они вместе называются родопсином . В колбочках присутствуют различные типы опсинов, которые в сочетании с ретиналем образуют пигменты, называемые фотопсинами . Три разных класса фотопсинов в колбочках реагируют на разные диапазоны частоты света, и такая избирательность позволяет зрительной системе преобразовывать цвета . Функция фоторецепторной клетки заключается в преобразовании световой информации фотона в форму информации, передаваемой нервной системе и легко используемой организмом: это преобразование называется трансдукцией сигнала .
Опсин, обнаруженный в светочувствительных ганглиозных клетках сетчатки, называется меланопсином . Эти клетки участвуют в различных рефлекторных реакциях мозга и тела на присутствие (дневного) света, таких как регуляция циркадных ритмов , зрачковый рефлекс и другие невизуальные реакции на свет. Меланопсин функционально напоминает опсины беспозвоночных.
Мозаика сетчатки
[ редактировать ]

Большинство фоторецепторов позвоночных расположены в сетчатке. Распределение палочек и колбочек (и их классов) в сетчатке называется мозаикой сетчатки . человека Каждая сетчатка содержит около 6 миллионов колбочек и 120 миллионов палочек. [ 8 ] В «центре» сетчатки (точке непосредственно за хрусталиком) находится ямка (или центральная ямка), содержащая только колбочки; и является ли областью, способной обеспечить самую высокую остроту зрения или самое высокое разрешение . В остальной части сетчатки палочки и колбочки переплетены. фоторецепторов не обнаружено . В слепом пятне (области, где волокна ганглиозных клеток собираются в зрительный нерв и выходят из глаза) [ 9 ] Распределение классов колбочек (L, M, S) также неоднородно: S-колбочки в ямке отсутствуют, а соотношение L-конусов и М-колбочек у разных людей различается.
Количество и соотношение палочек и колбочек варьируется у разных видов в зависимости от того, ведет ли животное преимущественно дневной или ночной образ жизни . Некоторые совы, например, ночная неясыть , [ 10 ] имеют огромное количество палочек в сетчатке. У других позвоночных также будет разное количество классов колбочек: от монохроматов до пентахроматов .
Сигнализация
[ редактировать ]
Путь зрительного сигнала описывается каскадом фототрансдукции — механизмом, с помощью которого энергия фотона сигнализирует механизму в клетке, который приводит к ее электрической поляризации. Эта поляризация в конечном итоге приводит либо к пропусканию, либо к подавлению нервного сигнала, который будет поступать в мозг через зрительный нерв . Шаги, которые применимы к пути фототрансдукции от фоторецепторов палочек/колбочек позвоночных:
- в Зрительный опсин позвоночных мембране диска внешнего сегмента поглощает фотон, изменяя конфигурацию сетчатки основания Шиффа кофактора внутри белка с цис-формы на транс-форму, вызывая изменение формы сетчатки.
- В результате образуется ряд нестабильных промежуточных продуктов, последний из которых сильнее связывается с белком G в мембране , называемым трансдуцином , и активирует его. Это первый этап амплификации: каждый фотоактивируемый опсин запускает активацию около 100 трансдуцинов.
- Затем каждый трансдуцин активирует фермент цГМФ-специфическую фосфодиэстеразу (ФДЭ).
- Затем ФДЭ катализирует гидролиз цГМФ до 5'-ГМФ. Это второй этап амплификации, на котором один ФДЭ гидролизует около 1000 молекул цГМФ.
- Чистая концентрация внутриклеточного цГМФ снижается (из-за его превращения в 5'-ГМФ посредством ФДЭ), что приводит к закрытию циклических нуклеотид-управляемых Na. + ионные каналы, расположенные в мембране наружного сегмента фоторецептора.
- В результате ионы натрия больше не могут проникать в клетку, а мембрана наружного сегмента фоторецептора становится гиперполяризованной , поскольку заряд внутри мембраны становится более отрицательным.
- Это изменение мембранного потенциала клетки приводит к закрытию потенциалзависимых кальциевых каналов. Это приводит к уменьшению притока ионов кальция в клетку и, следовательно, к падению внутриклеточной концентрации ионов кальция.
- Снижение внутриклеточной концентрации кальция означает, что меньше глутамата высвобождается посредством кальций-индуцированного экзоцитоза в биполярную клетку (см. ниже). (Снижение уровня кальция замедляет высвобождение нейромедиатора глутамата , который возбуждает постсинаптические биполярные клетки и горизонтальные клетки .)
- АТФ, обеспечиваемая внутренним сегментом, приводит в действие натриево-калиевый насос. Этот насос необходим для сброса исходного состояния внешнего сегмента, забирая ионы натрия, попавшие в клетку, и откачивая их обратно.
Гиперполяризация
[ редактировать ]В отличие от большинства сенсорных рецепторных клеток, фоторецепторы фактически становятся гиперполяризованными при стимуляции ; и наоборот, деполяризуются, когда их не стимулируют. Это означает, что глутамат высвобождается непрерывно, когда клетка не стимулируется, а стимул останавливает высвобождение. В темноте клетки имеют относительно высокую концентрацию циклического гуанозин-3'-5'-монофосфата (цГМФ), который открывает цГМФ-управляемые ионные каналы . Эти каналы неспецифичны и в открытом состоянии позволяют перемещать ионы натрия и кальция. Движение этих положительно заряженных ионов в клетку (движимое соответствующим электрохимическим градиентом ) деполяризует мембрану и приводит к высвобождению нейромедиатора глутамата .
Нестимулированные (в темноте) каналы, управляемые циклическими нуклеотидами во внешнем сегменте, открыты, поскольку циклический ГМФ с ними связан (цГМФ). Следовательно, положительно заряженные ионы (а именно натрия ионы ) попадают в фоторецептор, деполяризуя его примерно до -40 мВ ( потенциал покоя в других нервных клетках обычно составляет -65 мВ). Этот ток деполяризации часто называют темновым током.
Биполярные клетки
[ редактировать ]Фоторецепторы ( палочки и колбочки ) передают сигнал биполярным клеткам, которые затем передают сигнал ганглиозным клеткам сетчатки. Аксоны ганглиозных клеток сетчатки вместе образуют зрительный нерв , через который они проецируются в мозг. [ 8 ]
Фоторецепторы палочек и колбочек сигнализируют о поглощении фотонов посредством уменьшения высвобождения нейромедиатора глутамата в биполярные клетки на окончаниях их аксонов. Поскольку фоторецептор деполяризуется в темноте, в биполярных клетках в темноте выделяется большое количество глутамата. Поглощение фотона гиперполяризует фоторецептор и, следовательно, приводит к высвобождению меньшего количества глутамата на пресинаптическом терминале биполярной клетки.
Каждый фоторецептор палочки или колбочки высвобождает один и тот же нейромедиатор — глутамат. Однако эффект глутамата различается в биполярных клетках в зависимости от типа рецептора, встроенного в мембрану этой клетки . Когда глутамат связывается с ионотропным рецептором , биполярная клетка деполяризуется (и, следовательно, гиперполяризуется светом, поскольку выделяется меньше глутамата). С другой стороны, связывание глутамата с метаботропным рецептором приводит к гиперполяризации, поэтому эта биполярная клетка будет деполяризоваться к свету по мере высвобождения меньшего количества глутамата.
По сути, это свойство позволяет одной популяции биполярных клеток возбуждаться светом, а другой популяции — тормозиться им, хотя все фоторецепторы демонстрируют одинаковую реакцию на свет. Эта сложность становится одновременно важной и необходимой для определения цвета , контраста , краев и т. д.
Преимущества
[ редактировать ]Фототрансдукция в палочках и колбочках несколько необычна тем, что стимул (в данном случае свет) снижает реакцию клетки или скорость срабатывания, в отличие от большинства других сенсорных систем, в которых стимул увеличивает реакцию клетки или скорость срабатывания. Это различие имеет важные функциональные последствия:
- Классический фоторецептор (палочка или колбочка) деполяризуется в темноте, а это означает, что в клетку поступает много ионов натрия. Таким образом, случайное открытие или закрытие натриевых каналов не повлияет на мембранный потенциал клетки; только закрытие большого количества каналов за счет поглощения фотона повлияет на него и сигнализирует о том, что свет находится в поле зрения. Эта система может иметь меньше шума по сравнению со схемой сенсорной трансдукции, которая увеличивает скорость нервных импульсов в ответ на стимулы, такие как прикосновение и обоняние .
- на двух стадиях классической фототрансдукции происходит большая амплификация: один пигмент активирует множество молекул трансдуцина , а один ФДЭ расщепляет множество цГМФ. Это усиление означает, что даже поглощение одного фотона повлияет на мембранный потенциал и подаст сигнал мозгу о том, что свет находится в поле зрения. Это основная особенность, которая отличает палочковидные фоторецепторы от колбочек. Палочки чрезвычайно чувствительны и способны регистрировать один фотон света, в отличие от колбочек. С другой стороны, известно, что колбочки имеют очень быструю кинетику с точки зрения скорости усиления фототрансдукции, в отличие от палочек.
Разница между палочками и колбочками
[ редактировать ]Сравнение человеческих палочек и колбочек от Эрика Канделя и др. в принципах нейронауки . [ 11 ]
| Стержни | Конусы |
|---|---|
| Используется для скотопического зрения (зрения в условиях низкой освещенности). | Используется для фотопического зрения (зрение в условиях высокой освещенности). |
| Очень светочувствительный ; чувствителен к рассеянному свету | Не очень светочувствителен; чувствителен только к прямому свету |
| Потеря вызывает куриную слепоту | Убыток вызывает юридическую слепоту |
| Низкая острота зрения | Высокая острота зрения; лучшее пространственное разрешение |
| Нет в фовеа | Сосредоточено в фовеа |
| Медленная реакция на свет, раздражители добавляются со временем. | Быстрая реакция на свет, способность воспринимать более быстрые изменения стимулов. |
| Имеют больше пигмента, чем колбочки, поэтому могут обнаруживать более низкие уровни освещенности. | Имеют меньше пигмента, чем палочки, требуют больше света для обнаружения изображений. |
| Стопки покрытых мембраной дисков не прикреплены непосредственно к клеточной мембране. | Диски прикреплены к внешней мембране. |
| Около 120 миллионов палочек распределены по сетчатке. [ 8 ] | В каждой сетчатке распределено около 6 миллионов колбочек. [ 8 ] |
| Один тип светочувствительного пигмента | Три типа светочувствительного пигмента у человека |
| Даруют ахроматическое зрение | Дайте цветовое зрение |
Разработка
[ редактировать ]Ключевые события, опосредующие дифференцировку палочек, S-конусов и M-конусов, индуцируются несколькими факторами транскрипции, включая RORbeta, OTX2, NRL, CRX, NR2E3 и TRbeta2. Судьба S-конуса представляет собой программу фоторецепторов по умолчанию; однако дифференциальная транскрипционная активность может приводить к образованию палочек или М-конусов. L-конусы присутствуют у приматов, однако об их программе развития мало что известно, поскольку в исследованиях использовались грызуны. Существует пять этапов разработки фоторецепторов: пролиферация мультипотентных клеток-предшественников сетчатки (RPC); ограничение компетенции ПКР; спецификация судьбы клеток; экспрессия генов фоторецепторов; и, наконец, рост аксонов, образование синапсов и рост внешнего сегмента.
Ранняя передача сигналов Notch поддерживает цикличность предшественников. Предшественники фоторецепторов возникают за счет ингибирования передачи сигналов Notch и повышения активности различных факторов, включая гомолог 1 ахета-щитка. Активность OTX2 передает клеткам судьбу фоторецептора. CRX дополнительно определяет специфичную для фоторецепторов панель экспрессируемых генов. Экспрессия NRL приводит к судьбе палочки. NR2E3 дополнительно ограничивает клетки судьбой палочки путем репрессии генов колбочек. RORbeta необходим как для развития стержней, так и для развития колбочек. TRbeta2 опосредует судьбу М-конуса. Если какая-либо из функций ранее упомянутых факторов удалена, фоторецептором по умолчанию является S-конус. Эти события происходят в разные периоды времени для разных видов и включают в себя сложную картину действий, вызывающую целый спектр фенотипов. Если эти регуляторные сети нарушаются, это может привести к пигментному ретиниту , дегенерации желтого пятна или другим нарушениям зрения. [ 12 ]
Фоторецепторы ганглиозных клеток
[ редактировать ]Внутренне светочувствительные ганглиозные клетки сетчатки (ipRGC) представляют собой подмножество (≈1–3%) ганглиозных клеток сетчатки , в отличие от других ганглиозных клеток сетчатки, по своей природе светочувствительны из-за присутствия меланопсина , светочувствительного белка. Следовательно, они составляют третий класс фоторецепторов, помимо палочек и колбочек . [ 13 ]
У людей ipRGC участвуют в функциях, не формирующих изображение, таких как циркадные ритмы, поведение и зрачковый световой рефлекс . [ 14 ] Пиковая спектральная чувствительность рецептора составляет от 460 до 482 нм. [ 14 ] Однако они также могут способствовать развитию рудиментарного зрительного пути, обеспечивающего сознательное зрение и определение яркости. [ 14 ] Классические фоторецепторы (палочки и колбочки) также участвуют в работе новой зрительной системы, что может способствовать постоянству цвета. ipRGC могут сыграть важную роль в понимании многих заболеваний, включая основные причины слепоты во всем мире, такие как глаукома, заболевание, поражающее ганглиозные клетки, а изучение рецептора открывает потенциал в качестве нового направления для поиска методов лечения слепоты.
ipRGC были окончательно обнаружены у людей только во время знаковых экспериментов в 2007 году на людях без стержней и шишек. [ 15 ] [ 16 ] Как было обнаружено у других млекопитающих, у людей непалочковидный неколбочковый фоторецептор представляет собой ганглиозную клетку во внутренней сетчатке. Исследователи выследили пациентов с редкими заболеваниями, которые уничтожают функцию классических фоторецепторов палочек и колбочек, но сохраняют функцию ганглиозных клеток. [ 15 ] [ 16 ] Несмотря на отсутствие палочек или колбочек, у пациентов продолжали наблюдаться циркадные фотоэнцефалические изменения, циркадные поведенческие модели, подавление меланопсина и реакции зрачков, при этом пиковая спектральная чувствительность к окружающему и экспериментальному свету соответствовала таковой для фотопигмента меланопсин. Их мозг также мог ассоциировать зрение со светом этой частоты.
Нечеловеческие фоторецепторы
[ редактировать ]Палочки и колбочки фоторецепторы являются общими почти для всех позвоночных. Шишковидная и парапинеальная железы светочувствительны у позвоночных, не относящихся к млекопитающим, но не у млекопитающих. У птиц есть фотоактивные нейроны, контактирующие со спинномозговой жидкостью (СМЖ) в паравентрикулярном органе, которые реагируют на свет в отсутствие сигнала от глаз или нейротрансмиттеров. [ 17 ] Фоторецепторы беспозвоночных в таких организмах, как насекомые и моллюски, различаются как по своей морфологической организации, так и по лежащим в их основе биохимическим путям. В этой статье описаны фоторецепторы человека .
См. также
[ редактировать ]- Визуальная фототрансдукция
- Рецептор, связанный с G-белком
- Сенсорная система
- Светочувствительный
- Фоточувствительная ганглиозная клетка
- Горизонтальная ячейка
- Биполярная ячейка
- Амакриновая клетка
Ссылки
[ редактировать ]- ^ "глаз, человек." Британская энциклопедия. Полный справочный комплект Британской энциклопедии. Чикаго: Британская энциклопедия, 2010.
- ^ Фостер, Р.Г.; Провенсио, И.; Хадсон, Д.; Фиске, С.; Грип, В.; Менакер, М. (1991). «Циркадный фоторецепция у мышей с дегенерацией сетчатки (rd/rd)». Журнал сравнительной физиологии А. 169 (1): 39–50. дои : 10.1007/BF00198171 . ПМИД 1941717 . S2CID 1124159 .
- ^ Боумейкер Дж.К. и Дартналл HJA (1980). «Зрительные пигменты палочек и колбочек сетчатки человека» . Дж. Физиол . 298 : 501–511. дои : 10.1113/jphysicalol.1980.sp013097 . ПМЦ 1279132 . ПМИД 7359434 .
- ^ Физиология человека и механизмы болезней Артура К. Гайтона (1992) ISBN 0-7216-3299-8 стр. 373
- ^ Ричардсон, ТМ (1969). «Цитоплазматические и ресничные связи между внутренними и внешними сегментами зрительных рецепторов млекопитающих». Исследование зрения . 9 (7): 727–731. дои : 10.1016/0042-6989(69)90010-8 . ПМИД 4979023 .
- ^ Луви, А.; Гроув, Э.А. (2011). «Реснички в ЦНС: тихие органеллы занимают центральное место» . Нейрон . 69 (6): 1046–1060. дои : 10.1016/j.neuron.2011.03.002 . ПМК 3070490 . ПМИД 21435552 .
- ^ Основы видения , Брайан А. Ванделл
- ^ Перейти обратно: а б с д Шактер, Дэниел Л. (2011). Психология Второе издание . Нью-Йорк: Издательство Worth. стр. 136–137 . ISBN 978-1-4292-3719-2 .
- ^ Гольдштейн, Э. Брюс (2007). Ощущение и восприятие (7-е изд.). Томсон и Уодсвот.
- ^ «Информация о совиных глазах» . совы.орг . Всемирный фонд сов. Архивировано из оригинала 16 февраля 2018 года . Проверено 1 мая 2017 г.
- ^ Свеча, скорая помощь; Шварц, Дж. Х.; Джесселл, ТМ (2000). Принципы нейронауки (4-е изд.). Нью-Йорк: МакГроу-Хилл. стр. 100-1 507–513 . ISBN 0-8385-7701-6 .
- ^ Сваруп, Ананд; Дуглас Ким; Дуглас Форрест (август 2010 г.). «Транкрипционная регуляция развития фоторецепторов и гомеостаз в сетчатке млекопитающих» . Обзоры природы Неврология . 11 (8): 563–576. дои : 10.1038/nrn2880 . ПМИД 20648062 . S2CID 6034699 .
- ^ До МТ, Яу К.В. (октябрь 2010 г.). «Самостоятельно светочувствительные ганглиозные клетки сетчатки» . Физиологические обзоры . 90 (4): 1547–81. doi : 10.1152/physrev.00013.2010 . ПМЦ 4374737 . ПМИД 20959623 .
- ^ Перейти обратно: а б с Заиди Ф.Х. и др. (2007). «Коротковолновая световая чувствительность циркадного, зрачкового и зрительного восприятия у людей, лишенных внешней сетчатки» . Современная биология . 17 (24): 2122–8. Бибкод : 2007CBio...17.2122Z . дои : 10.1016/j.cub.2007.11.034 . ПМК 2151130 . ПМИД 18082405 .
- ^ Перейти обратно: а б Коглан А. Слепые люди «видят» восход и закат солнца . New Scientist, 26 декабря 2007 г., выпуск 2635.
- ^ Перейти обратно: а б Медицинские новости сегодня. Нормальные реакции на невизуальные эффекты света, удерживаемые слепыми людьми, у которых нет палочек и колбочек. Архивировано 6 февраля 2009 г. в Wayback Machine . 14 декабря 2007 г.
- ^ «Ученые зарегистрировали светочувствительные глаза птиц в мозгу птиц» . www.birdnews.com . Новости птиц. Архивировано из оригинала 2 июля 2017 года . Проверено 20 июля 2017 г.
Библиография
[ редактировать ]- Кэмпбелл, Нил А. и Рис, Джейн Б. (2002). Биология . Сан-Франциско: Бенджамин Каммингс. стр. 1064–1067 . ISBN 0-8053-6624-5 .
- Фриман, Скотт (2002). Биологические науки (2-е изд.). Энглвуд Клиффс, Нью-Джерси: Прентис Холл. стр. 835–837 . ISBN 0-13-140941-7 .
Внешние ссылки
[ редактировать ] СМИ, связанные с фоторецепторными клетками , на Викискладе?
СМИ, связанные с фоторецепторными клетками , на Викискладе? - Поиск NIF – фоторецепторная клетка через информационную структуру нейробиологии

| Базы данных органов управления : Национальные |
|---|