Тетрахромация

Тетрахроматия (от греч. «Tetra» , что означает «четыре», и «chroma» , что означает «цвет») — это состояние наличия четырех независимых каналов для передачи цветовой информации или наличие четырех типов колбочек в глазу . Организмы, обладающие тетрахроматией, называются тетрахроматами.
У тетрахроматических организмов сенсорное цветовое пространство является четырехмерным, а это означает, что для сопоставления сенсорного эффекта произвольно выбранных спектров света в их видимом спектре требуется смесь как минимум четырех основных цветов .
Тетрахроматия продемонстрирована у нескольких видов птиц . [2] рыбы , [3] и рептилии . [3] Общим предком всех позвоночных был тетрахромат, но общий предок млекопитающих потерял два из четырех типов колбочек, развив дихроматию , утрату, приписываемую предполагаемому ночному узкому месту . у некоторых приматов развился третий конус. Позже [4]
Физиология
[ редактировать ]организма Обычное объяснение тетрахромазии состоит в том, что сетчатка содержит четыре типа световых рецепторов более высокой интенсивности (называемых колбочками у позвоночных, в отличие от палочек , которые представляют собой световые рецепторы более низкой интенсивности) с различной спектральной чувствительностью . Это означает, что организм может видеть длины волн, превосходящие возможности зрения обычного человека, и может различать цвета, которые обычному человеку кажутся идентичными . Виды с тетрахроматическим цветовым зрением могут иметь неизвестное физиологическое преимущество перед конкурирующими видами. [5]
Люди
[ редактировать ]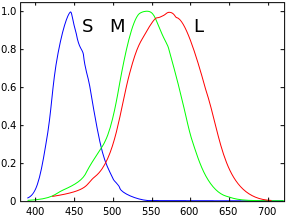
Обезьяны (включая людей ) и обезьяны Старого Света обычно имеют только три типа колбочек и, следовательно, являются трихроматами . Однако предполагается, что тетрахроматия человека существует у небольшого процента населения. Трихроматы имеют три типа колбочек, каждый тип чувствителен к соответствующей части спектра, как показано на схеме. Но по крайней мере одна женщина считается тетрахроматом. [6] Точнее, она имела дополнительный тип колбочек L' , промежуточный между M и L по своей чувствительности, и демонстрировала трехмерную ( компоненты M , L' и L ) цветовую дискриминацию для длин волн 546–670 нм (к которым относится четвертый тип, S , нечувствителен).
Тетрахроматия требует существования четырех независимых классов фоторецепторных клеток с различной спектральной чувствительностью . Однако также должен существовать соответствующий пострецепторный механизм для сравнения сигналов от четырех классов рецепторов. Согласно теории оппозиционного процесса , у людей есть три оппозиционных канала, которые дают трихроматию. Неясно, достаточно ли наличия четвертого противостоящего канала для тетрахромазии. [ нужна ссылка ]
Мыши, которые обычно имеют только два колбочек-пигмента (и, следовательно, два противостоящих канала), были созданы для экспрессии третьего колбочек-пигмента и, по-видимому, демонстрируют повышенную хроматическую дискриминацию. [7] возможно, указывая на трихроматию и предполагая, что они смогли создать или повторно включить третий канал противника. Это поддержало бы теорию о том, что люди должны иметь возможность использовать четвертый канал противника для тетрахроматического зрения. Однако утверждения оригинальной публикации о пластичности зрительного нерва также оспариваются. [8]
Тетрахроматия у носителей ССЗ
[ редактировать ]Было высказано предположение, что женщины, несущие рецессивные опсина аллели , которые могут вызывать дефицит цветового зрения (ССЗ), могут обладать тетрахроматией. Женщины -носители аномальной трихромазии (легкой дальтонизма) обладают гетерозиготными аллелями генов, кодирующих L-опсин или М-опсин . Эти аллели часто имеют разную спектральную чувствительность , поэтому, если носитель экспрессирует обе аллели опсина, они могут проявлять тетрахроматию.
У человека конусных клеток присутствуют два гена пигмента на Х-хромосоме : классический ген опсина 2 типа OPN1MW и OPN1MW2 . Люди с двумя Х-хромосомами могут обладать несколькими пигментами колбочек, возможно, рожденными как полные тетрахроматы, у которых есть четыре одновременно функционирующих типа колбочек, каждый из которых имеет определенный образец реагирования на разные длины волн света в диапазоне видимого спектра. [9] Одно исследование показало, что 15% женщин в мире могут иметь тип четвертой колбочки, пик чувствительности которой находится между стандартными красными и зелеными колбочками, что теоретически дает значительное увеличение цветовой дифференциации. [10] Другое исследование показывает, что до 50% женщин и 8% мужчин могут иметь четыре фотопигмента и соответствующую повышенную хроматическую дискриминацию по сравнению с трихроматами. [11] В 2010 году, после двадцатилетнего исследования женщин с четырьмя типами колбочек (нефункциональные тетрахроматы), нейробиолог Габриэль Джордан выявила женщину (субъект «cDa29» ), которая могла различать большее разнообразие цветов, чем трихроматы, что соответствует функциональному или «истинный» тетрахромат. [6] [12] В частности, было показано, что она является трихроматом в диапазоне 546–670 нм, где люди с нормальным зрением по сути являются дихроматами из-за незначительной реакции S- конусов на эти длины волн. Таким образом, если S- конусы «cDa29» обеспечивают независимое измерение восприятия цвета, как обычно, это подтвердит, что она является тетрахоматом, если рассматривать весь спектр.
Вариации генов пигмента колбочек широко распространены в большинстве популяций человека, но наиболее распространенная и выраженная тетрахроматия происходит от женщин-носителей основных аномалий красного/зеленого пигмента, обычно классифицируемых как формы « дальтонизма » ( протаномалия или дейтераномалия ). Биологической основой этого явления является Х-инактивация гетерозиготных аллелей генов пигмента сетчатки, что является тем же механизмом, который дает большинству самок обезьян Нового Света трехцветное зрение. [13]
У человека предварительная обработка изображений происходит в нейронах сетчатки . Неизвестно, как эти нервы отреагируют на новый цветовой канал: будут ли они обрабатывать его отдельно или просто объединить с одним из существующих каналов. Точно так же визуальная информация покидает глаз через зрительный нерв, и в мозгу происходит различная окончательная обработка изображения; неизвестно, обладают ли зрительный нерв или области мозга способностью эффективно реагировать на стимул в виде нового цветового сигнала .
Тетрахроматия также может улучшить зрение при тусклом освещении или при взгляде на экран. [14] [ не удалось пройти проверку ]
Условная тетрахроматия
[ редактировать ]Несмотря на то, что люди являются трихроматами, люди могут испытывать легкую тетрахроматию при низкой интенсивности света , используя свое мезопическое зрение . При мезопическом зрении как колбочки , так и палочки активны . Хотя палочки обычно не способствуют цветовому зрению, в этих специфических условиях освещения они могут давать небольшую область тетрахроматики в цветовом пространстве. [15] Чувствительность палочек человека наиболее высока при длине волны 500 нм (голубовато-зеленая), которая значительно отличается от пиковой спектральной чувствительности колбочек (обычно 420, 530 и 560 нм).
Блокированная тетрахромация
[ редактировать ]Хотя многие птицы являются тетрахроматами с четвертым цветом ультрафиолета, люди не могут видеть ультрафиолетовый свет напрямую, потому что хрусталик глаза блокирует большую часть света в диапазоне длин волн 300–400 нм; более короткие волны блокируются роговицей . [16] Фоторецепторные клетки сетчатки ), видят ближний ультрафиолетовый свет (вплоть до 300 нм) как беловато-синий, а для некоторых длин волн — беловато - чувствительны к ближнему ультрафиолетовому свету, и люди, у которых нет хрусталика (состояние, известное как афакия фиолетовый, вероятно, потому, что все три типа колбочек примерно одинаково чувствительны к ультрафиолетовому свету (при этом синие колбочки немного более чувствительны). [17]
Хотя расширенный видимый диапазон не означает тетрахроматию, некоторые полагают, что доступны зрительные пигменты с чувствительностью в длинах волн, близких к УФ , что делает возможным тетрахромацию в случае афакии . [18] Однако нет никаких рецензируемых доказательств, подтверждающих это утверждение.
Другие животные
[ редактировать ]
Рыба
[ редактировать ]Рыбы, особенно костистые , обычно являются тетрахроматами. [3] Исключения включают:
- Акулы и скаты - варьируются от монохромности до трихромазии. [3]
- Глубоководные рыбы – часто палочки-монохроматы.
- Цихлида - предположительно пентахроматия или выше. [3]
Птицы
[ редактировать ]Некоторые виды птиц, такие как зебра-зяблик и Columbidae , используют ультрафиолетовую длину волны 300–400 нм, специфичную для тетрахроматического цветового зрения, в качестве инструмента во время выбора партнера и поиска пищи . [19] При выборе партнеров ультрафиолетовое оперение и окраска кожи показывают высокий уровень отбора. [20] Типичный птичий глаз реагирует на длины волн примерно 300–700 нм. По частоте это соответствует полосе в районе 430–1000 ТГц . У большинства птиц сетчатка имеет четыре спектральных типа колбочек, которые, как полагают, обеспечивают тетрахроматическое цветовое зрение. Цветовое зрение птиц дополнительно улучшается за счет фильтрации пигментированных капель масла в фоторецепторах. Капли масла фильтруют падающий свет до того, как он достигнет зрительного пигмента во внешних сегментах фоторецепторов.
Четыре типа колбочек и специализация пигментированных капель масла обеспечивают птицам лучшее цветовое зрение, чем у людей. [21] [22] Однако более поздние исследования показали, что тетрахроматия у птиц обеспечивает птицам только более широкий зрительный спектр, чем у людей (люди не могут видеть ультрафиолетовый свет, 300–400 нм ), в то время как спектральное разрешение («чувствительность» к нюансам) аналогично. . [23]
Многие дневные хищные птицы, такие как орлы, соколы и ястребы, имеют ограниченную способность воспринимать ультрафиолетовый свет. Считается, что ультрафиолетовое зрение выбрано против , потому что хроматические аберрации существенно увеличиваются с увеличением чувствительности к ультрафиолету А , снижая остроту зрения, на которую хищные птицы полагаются при охоте. [24]
Пентахроматия и выше
[ редактировать ]Размерность с цветовым зрением цветового зрения не имеет верхней границы, но позвоночные , превышающим тетрахроматию, встречаются редко. Следующий уровень — пентахроматия , то есть пятимерное цветовое зрение, требующее как минимум 5 различных классов информации через первичную зрительную фоторецепторов, а также 5 независимых каналов цветовой систему.
Женщина, гетерозиготная как по опсинам LWS , так и по MWS (и, следовательно, являющаяся носителем как протаномалии , так и дейтераномалии ), будет экспрессировать пять опсинов различной спектральной чувствительности . Однако для того, чтобы она была настоящим (сильным) пентахроматом, эти опсины должны быть разделены на разные фоторецепторные клетки , и ей нужно было бы иметь соответствующие пострецепторные механизмы для обработки 5 каналов противостоящих процессов , что является спорным. [ нужна ссылка ]
Некоторые птицы (особенно голуби ) имеют пять или более видов цветовых рецепторов в сетчатке и поэтому считаются пентахроматами, хотя психофизические доказательства функциональной пентахроматики отсутствуют. [25] Исследования также показывают, что некоторые миноги , представители Petromyzontiformes , могут быть пентахроматами. [26]
Беспозвоночные могут иметь большое количество различных классов опсинов, в том числе 15 опсинов у бабочек-синиц. [27] или 33 у креветок-богомолов . [28] Однако не показано, что цветовое зрение у этих беспозвоночных имеет размерность, соизмеримую с числом опсинов.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Данные рисунка, соответствует нескорректированной кривой поглощения, взято из Харт, Н.С.; Партридж, Дж. К.; Беннетт, АТД; Катхилл, IC (2000). «Зрительные пигменты, капли конусного масла и глазные среды у четырех видов эстрильдидных вьюрков». Журнал сравнительной физиологии А. 186 (7–8): 681–694. дои : 10.1007/s003590000121 . ПМИД 11016784 . S2CID 19458550 .
- ^ Голдсмит, Тимоти Х. (2006). «Что видят птицы». Scientific American (июль 2006 г.): 69–75.
- ^ Jump up to: а б с д и Боумейкер, Джеймс К. (сентябрь 2008 г.). «Эволюция зрительных пигментов позвоночных» . Исследование зрения . 48 (20): 2022–2041. дои : 10.1016/j.visres.2008.03.025 . ПМИД 18590925 . S2CID 52808112 .
- ^ Джейкобс, GH (2009). «Эволюция цветового зрения у млекопитающих» . Фил. Пер. Р. Сок. Б. 364 (1531): 2957–2967. дои : 10.1098/rstb.2009.0039 . ПМК 2781854 . ПМИД 19720656 .
- ^ Бакхаус В., Клигл Р., Вернер Дж.С. (1998). Цветовое зрение: перспективы разных дисциплин . Де Грютер. стр. 163–182. ISBN 9783110806984 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Jump up to: а б Джордан, Габриэле; Диб, Самир С.; Бостен, Дженни М.; Моллон, доктор юридических наук (июль 2010 г.). «Размерность цветового зрения у носителей аномальной трихромазии» . Журнал видения . 10 (12): 12. дои : 10.1167/10.8.12 . ПМИД 20884587 .
- ^ Джейкобс, Джеральд Х.; Уильямс, Гэри А.; Кэхилл, Хью; Натанс, Джереми (23 марта 2007 г.). «Появление нового цветового зрения у мышей, созданного для экспрессии фотопигмента человеческой колбочки». Наука . 315 (5819): 1723–1725. Бибкод : 2007Sci...315.1723J . дои : 10.1126/science.1138838 . ПМИД 17379811 . S2CID 85273369 .
- ^ Макоус, В. (12 октября 2007 г.). «Комментарий к статье «Появление нового цветового зрения у мышей, созданного для экспрессии фотопигмента колбочек человека»» . Наука . 318 (5848): 196. Бибкод : 2007Sci...318..196M . дои : 10.1126/science.1146084 . ПМИД 17932271 .
- ^ Джеймсон, Калифорния; Хайноут, СМ; Вассерман, LM (2001). «Более богатый цветовой опыт у наблюдателей с несколькими генами опсина фотопигмента» (PDF) . Психономический бюллетень и обзор . 8 (2): 244–261. дои : 10.3758/BF03196159 . ПМИД 11495112 . S2CID 2389566 . Архивировано из оригинала (PDF) 14 февраля 2012 года – через Cal Tech .
- ^ Джордан, Г. (июль 1993 г.). «Исследование женщин, гетерозиготных по дефициту цвета». Исследование зрения . 33 (11): 1495–1508. дои : 10.1016/0042-6989(93)90143-К . ПМИД 8351822 . S2CID 17648762 .
- ^ Бакхаус, Вернер Г.К.; Бакхаус, Вернер; Клигль, Рейнхольд; Вернер, Джон Саймон (1998). Цветовое зрение: перспективы разных дисциплин . Вальтер де Грютер. ISBN 9783110161007 – через книги Google.
- ^ Жительнице Сан-Диего, Кончетте Антико, поставили диагноз «суперзрение» (видео). 22 ноября 2013 г. — через YouTube .
- ^ Фрэнсис, Ричард К. (2011). «Глава 8. Женщины Икс». Эпигенетика: величайшая тайна наследственности . Нью-Йорк, Нью-Йорк и Лондон, Великобритания: WW Norton. стр. 93–104. ISBN 978-0-393-07005-7 .
- ^ Робсон, Дэвид (5 сентября 2014 г.). «Женщины со сверхчеловеческим видением» . Новости Би-би-си . Архивировано из оригинала 13 сентября 2014 года . Проверено 30 декабря 2017 г.
- ^ Аутрум, Хансйохем и Юнг, Ричард (1973). Интегративные функции и сравнительные данные . Том. 7. Шпрингер-Верлаг. п. 226. ИСБН 978-0-387-05769-9 – через книги Google.
- ^ Мейнстер, Массачусетс (2006). «Фиолетовый и синий свет, блокирующий интраокулярные линзы: фотозащита и фоторецепция» . Британский журнал офтальмологии . 90 (6): 784–792. дои : 10.1136/bjo.2005.086553 . ПМК 1860240 . ПМИД 16714268 .
- ^ Хэмблинг, Дэвид (29 мая 2002 г.). «Пусть свет сияет» . Хранитель .
- ^ Фултон, Джеймс Т. (31 июля 2009 г.). «Человек — это заблокированный тетрахромат» . Neuronresearch.net . Проверено 1 июня 2022 г.
- ^ Беннетт, Эндрю Т.Д.; Катхилл, Иннес К.; Партридж, Джулиан К.; Майер, Эрхард Дж. (1996). «Ультрафиолетовое зрение и выбор партнера у зебровых амадин». Природа . 380 (6573): 433–435. Бибкод : 1996Natur.380..433B . дои : 10.1038/380433a0 . S2CID 4347875 .
- ^ Беннетт, Эндрю Т.Д.; Тери, Марк (2007). «Цветовое зрение и окраска птиц: междисциплинарная эволюционная биология» (PDF) . Американский натуралист . 169 (С1): С1–С6. дои : 10.1086/510163 . ISSN 0003-0147 . JSTOR 510163 . S2CID 2484928 .
- ^ Катхилл, Иннес К.; Партридж, Джулиан К.; Беннетт, Эндрю Т.Д.; Черч, Стюарт К.; Харт, Натан С.; Хант, Сара (2000). Слейтер, Питер Дж.Б.; Розенблатт, Джей С.; Сноудон, Чарльз Т.; Ропер, Тимоти Дж. (ред.). Ультрафиолетовое зрение у птиц . Достижения в изучении поведения. Том. 29. Академическая пресса. п. 159. дои : 10.1016/S0065-3454(08)60105-9 . ISBN 978-0-12-004529-7 .
- ^ Воробьев М. (ноябрь 1998 г.). «Тетрахроматия, капли масла и цвета оперения птиц». Журнал сравнительной А. физиологии 183 (5): 621–33. дои : 10.1007/s003590050286 . ПМИД 9839454 . S2CID 372159 .
- ^ Олссон, Питер; Линд, Олле; Кельбер, Альмут (15 января 2015 г.). «Цветовое зрение птиц: поведенческие пороги выявляют рецепторный шум» . Журнал экспериментальной биологии . 218 (2): 184–193. дои : 10.1242/jeb.111187 . ISSN 0022-0949 . ПМИД 25609782 .
- ^ Линд, Олле; Миткус, Миндаугас; Олссон, Питер; Кельбер, Альмут (15 мая 2013 г.). «Чувствительность к ультрафиолету и цветовое зрение при кормлении хищников» . Журнал экспериментальной биологии . 216 (Часть 10): 1819–1826. дои : 10.1242/jeb.082834 . ISSN 0022-0949 . ПМИД 23785106 . S2CID 3138290 . Проверено 13 июня 2023 г.
- ^ Эммертон, Джеки; Делис, Хуан Д. (1980). «Дискриминация длин волн в «видимом» и ультрафиолетовом спектре голубями» . Журнал сравнительной физиологии А. 141 (1): 47–52. дои : 10.1007/BF00611877 . S2CID 12462107 .
- ^ Дэвис, Уэйн Л.; Коуинг, Джилл А.; Карвальо, Ливия С.; Поттер, Ян С.; Трезизе, Энн Э.О.; Хант, Дэвид М.; Коллин, Шон П. (сентябрь 2007 г.). «Функциональная характеристика, настройка и регуляция экспрессии генов зрительного пигмента у анадромной миноги» . Журнал ФАСЭБ . 21 (11): 2713–2724. дои : 10.1096/fj.06-8057com . ПМИД 17463225 . S2CID 5932623 .
- ^ Чен, Пей-Джу; Авата, Хироко; Мацусита, Ацуко; Ян, Энь-Чэн; Арикава, Кентаро (8 марта 2016 г.). «Чрезвычайное спектральное богатство глаз обыкновенной бабочки Bluebottle Graphium sarpedon» . Границы экологии и эволюции . 4 . дои : 10.3389/fevo.2016.00018 .
- ^ Хансен, Сара (17 июля 2020 г.). «Глаза креветок-богомолов становятся еще более дикими: команда UMBC обнаружила в два раза больше ожидаемого количества светочувствительных белков - UMBC: Университет Мэриленда, округ Балтимор» . УМБК . Проверено 7 октября 2022 г.
Внешние ссылки
[ редактировать ]- Голдсмит, Тимоти Х. «Что видят птицы», журнал Scientific American, июль 2006 г. Статья о тетрахроматическом зрении птиц.
- Томпсон, Эван (2000). «Сравнительное цветовое зрение: качественное пространство и визуальная экология». Стивен Дэвис (ред.), «Восприятие цвета: философские, психологические, художественные и вычислительные перспективы» , стр. 163–186. Оксфорд: Издательство Оксфордского университета.
- В поисках мадам Тетрахромат , Гленн Зорпетт. Red Herring , 1 ноября 2000 г. Журнал
- «Исследование четвертого измерения». Архивировано 15 апреля 2016 г. в Wayback Machine . Бристольского университета Школа биологических наук . 20 марта 2009 г.
- Цвета - идеальный желтый от Radiolab, 21 мая 2012 г. (исследуется тетрахроматия у людей)
- Размерность цветового зрения у носителей аномальной трихромазии — Габриэль Джордан и др. — Журнал Vision, 12 августа 2010 г.:
- О тетрахромазии Агнес Хольба и Б. Лукач
Цветовые темы |
|---|