ЭСКРТ
Эндосомальные сортировочные комплексы, необходимые для механизма транспорта ( ESCRT ), состоят из цитозольных белковых комплексов, известных как ESCRT-0, ESCRT-I, ESCRT-II и ESCRT-III. Вместе с рядом дополнительных белков эти комплексы ESCRT обеспечивают уникальный способ ремоделирования мембран , который приводит к изгибу/отпочкованию мембран от цитоплазмы . [ 1 ] [ 2 ] Эти компоненты ESCRT были выделены и изучены на ряде организмов, включая дрожжи и человека. [ 3 ] Этот эукариот механизм обнаружен у всех и некоторых архей . [ 4 ]
Механизм ESCRT играет жизненно важную роль в ряде клеточных процессов, включая биогенез мультивезикулярных телец (MVB), клеточное отделение и почкование вируса . Биогенез мультивезикулярных телец (MVB) представляет собой процесс, при котором меченные убиквитином белки проникают в органеллы, называемые эндосомами, посредством образования везикул . Этот процесс необходим клеткам для разрушения неправильно свернутых и поврежденных белков. [ 5 ] Без аппарата ESCRT эти белки могут накапливаться и приводить к нейродегенеративным заболеваниям. Например, нарушения в компонентах ESCRT-III могут привести к неврологическим расстройствам, таким как наследственная спастическая параплегия (HSP). [ 6 ] Отторжение клетки, процесс, при котором мембрана, соединяющая две дочерние клетки, расщепляется, также опосредуется механизмом ESCRT. Без комплексов ESCRT дочерние клетки не могли бы разделяться, и аномальные клетки, содержащие вдвое больше ДНК возникали бы . Эти клетки неизбежно будут уничтожены посредством процесса, известного как апоптоз . Наконец, вирусное почкование или процесс выхода определенных типов вирусов из клеток может не происходить в отсутствие механизма ESCRT. Это неизбежно предотвратит распространение вирусов от клетки к клетке.
Комплексы ESCRT и вспомогательные белки
[ редактировать ]
Каждый из комплексов ESCRT и вспомогательные белки имеют уникальную структуру, обеспечивающую различные биохимические функции. Для каждого белкового компонента механизма ESCRT существует ряд синонимов как у дрожжей, так и у многоклеточных животных . Сводная таблица всех этих белков представлена ниже.
У дрожжей существуют следующие комплексы/вспомогательные белки:
ЭСКРТ-0
[ редактировать ]Комплекс ESCRT-0 играет жизненно важную роль в образовании мультивезикулярных телец путем связывания и кластеризации убиквитинированных белков и/или рецепторов на поверхности клетки. Затем комплекс отвечает за связывание с липидом на эндосомальной мембране, который рекрутирует эти меченые белки в эндосому. [ 7 ] После правильной локализации эти белки затем попадают в эндосому через везикулы, образуя мультивезикулярные тельца, и в конечном итоге доставляются в лизосому , где они разрушаются. Этот процесс важен, поскольку он является основным путем деградации поврежденных белков, прошедших через Гольджи . [ 5 ] Компоненты комплекса ЭСКРТ-0 существуют следующие:
Комплекс представляет собой гетеродимер 1:1 Vps27 ( белок 27 , сортирующий вакуолярный белок ) и Hse1 . [ 1 ] [ 6 ] Vps27 и Hse1 димеризуются через антипараллельные спиральные домены GAT (названные так в честь белков GGA и Tom1). [ 1 ] И Vps27, и Hse1 содержат аминоконцевой домен VHS (названный так потому, что он содержится в V ps27, H RS и ST TAM). белках [ 8 ] Эти домены VHS связывают убиквитин с белками, которые клетка стремится разрушить. Убиквитин также может ассоциироваться с мотивами, взаимодействующими с убиквитином, такими как мотив Hse1 или двухсторонний домен, обнаруженный на Vps27. Домен FYVE (названный в честь четырех белков, в которых он был первоначально идентифицирован: Fab1p, YOTB, Vac1 и EEA1) находится между VHS и взаимодействующими с убиквитином мотивными доменами Vps27. [ 6 ] [ 9 ] Фосфатидилинозит-3-фосфат , общий эндосомальный липид, связывается с этим доменом FYVE, что приводит к привлечению ESCRT-0 в эндосому. [ 6 ]
ЭСКРТ-I
[ редактировать ]Роль комплекса ESCRT-I заключается в содействии образованию мультивезикулярных телец путем кластеризации убиквитинированных белков и действия в качестве моста между комплексами ESCRT-0 и ESCRT-II. [ 10 ] Он также играет роль в распознавании и ремоделировании мембран во время отделения мембраны, образуя кольца по обе стороны от среднего тела делящихся клеток. ESCRT-I также отвечает за рекрутирование ESCRT-III, который формирует зону сужения непосредственно перед разделением клеток. [ 11 ] Более того, ESCRT-I играет роль в почковании вируса, взаимодействуя со специфическими вирусными белками, что приводит к привлечению дополнительных механизмов ESCRT к потенциальному месту высвобождения вируса. [ 12 ] Подробности оборудования ESCRT-I описаны ниже.
Комплекс ESCRT-I представляет собой гетеротетрамер (1:1:1:1) Vps23, Vps28 , Vps37 и Mvb12. [ 3 ] Собранный гетеротетрамер выглядит как палочковидный стебель, состоящий из Vps23, Vps37 и Mvb12, с веерообразной шляпкой, состоящей из одиночных спиралей Vps23, Vps28 и Vps37. [ 3 ] [ 6 ] Vps23 содержит один вариантный домен убиквитина E2, который отвечает за связывание убиквитина, комплекса ESCRT-0 и мотива PTAP ( треонин . , ланин Gag , пролин ) вирусных пролин - белков , [ 3 ] [ 6 ] богатый пролином мотив (GPPX 3 Y), который направляет ESCRT-I к срединному телу во время отторжения мембраны. Сразу после этого вариантного домена убиквитина E2 присутствует [ 6 ] Mvb12 также может связывать убиквитин через свой карбокси-конец . Vps28 отвечает за взаимодействие ESCRT-I и ESCRT-II путем ассоциации с доменом GLUE ( GRAM - подобное бикитин U- -связывание в E AP45) Vps36 через его карбокси-концевой четырехспиральный домен пучка. [ 1 ]
ЭСКРТ-II
[ редактировать ]Комплекс ESCRT-II функционирует преимущественно во время биогенеза мультивезикулярных телец и доставки белков, меченных убиквитином, в эндосому. Белки, меченные убиквитином, передаются из ESCRT-0 в ESCRT-I, а затем в ESCRT-II. ESCRT-II соединяется с ESCRT-III, который закрывает груз, содержащий везикулу. [ 6 ] Конкретные аспекты ESCRT-II заключаются в следующем:
ESCRT-II представляет собой гетеротетрамер (2:1:1), состоящий из двух субъединиц Vps25 , одной субъединицы Vps22 и одной субъединицы Vps36 . [ 3 ] Молекулы Vps25 содержат мотивы PPXY, которые связываются с мотивами крылатой спирали (WH) Vps22 и Vps36, образуя Y-образный комплекс с Vps22 и Vps36 в качестве основания и молекулами Vps25 в качестве плеч. [ 3 ] [ 6 ] Молекулы Vps25 также содержат мотивы WH, которые отвечают за взаимодействие ESCRT-II с ESCRT-III. Vps36 содержит домен GLUE, который связывает фосфатидилинозитол-3-фосфат и Vps28 ESCRT-I. [ 3 ] [ 6 ] Два домена «цинковых пальцев» встраиваются в домен GLUE дрожжевого Vps36. Один из этих доменов цинковых пальцев связывает карбокси-концевой домен Vps28, а другой ассоциируется с убиквитином. [ 6 ]
ЭСКРТ-III
[ редактировать ]Комплекс ESCRT-III, вероятно, является наиболее важным из всех механизмов ESCRT, поскольку он играет роль во всех процессах, опосредованных ESCRT. [ 13 ] Во время отрыва мембраны и отпочкования вируса ESCRT-III образует длинные нити, которые обвиваются вокруг места сужения мембраны непосредственно перед ее расщеплением. [ 11 ] [ 14 ] Это опосредование отторжения происходит посредством взаимодействия с центрально-шпиндлиновым комплексом. [ 15 ] Эти нитевидные структуры также присутствуют во время формирования мультивезикулярных телец и функционируют как кольцеобразный забор, который закупоривает отпочковавшуюся везикулу, чтобы предотвратить выход белков-грузов в цитозоль клетки. [ 11 ] ESCRT-III существует и функционирует следующим образом:
Комплекс ESCRT-III отличается от всех других механизмов ESCRT тем, что он существует лишь временно и содержит как существенные, так и несущественные компоненты. [ 1 ] [ 11 ] основные субъединицы должны собраться в правильном порядке (Vps20, Snf7, Vps24 , затем Vps2). Чтобы механизм функционировал, [ 6 ] Несущественные субъединицы включают Vps60, Did2 и Ist1. [ 11 ] Vps20 инициирует сборку ESCRT-III, действуя как зародыш сборки полимера Snf7. Затем Vps24 объединяется с Snf7, чтобы ограничить комплекс и завербовать Vps2. [ 1 ] [ 3 ] Затем Vps2 подключает к комплексу Vps4. [ 16 ] Все «свободные» цитозольные формы каждой субъединицы считаются закрытыми. То есть карбокси-концевая часть каждой субъединицы сворачивается сама в себя автоингибирующим образом, стабилизируя мономерные субъединицы. [ 1 ] [ 3 ] Карбокси-конец большинства субъединиц ESCRT-III, как эссенциальных, так и заменимых, содержат мотивы MIM ( ( взаимодействующий микротрубочек транспортный домен и ) мотивы MTIT ). [ 17 ] Эти мотивы ответственны за связывание Vps4 и AAA-АТФазы спастина . [ 3 ]
Впс4-Вта1
[ редактировать ]Белки Vps4-Vta1 необходимы для удаления других компонентов ESCRT (обычно ESCRT-III) из мембран после завершения определенного процесса. Ведутся споры о том, расщепляет ли Vps4 комплекс ESCRT-III или реконструирует комплекс так, чтобы один компонент удалялся в определенное время. [ 12 ] Считается, что Vta1 действует как активатор Vps4, помогая его сборке и усиливая активность AAA-АТФазы. [ 13 ] [ 18 ] Функционирование этих белков следующее:
Субъединицы Vps4 имеют два функциональных домена: аминоконцевой домен MIT и центральный домен AAA-АТФазы. [ 3 ] Домен MIT отвечает за взаимодействие Vps4 с доменом MIM Vps2. [ 1 ] Домен AAA-ATPase гидролизует АТФ, обеспечивая разборку комплекса ESCRT-III. [ 11 ] Такое «зачистка» ESCRT-III позволяет переработать все связанные субъединицы для дальнейшего использования. [ 11 ] [ 12 ] Vta1 представляет собой димерный белок, содержащий один домен VSL (названный так потому, что он обнаружен в белках V ps4, S BP1 и L IP5), который обеспечивает связывание с Vps4, и домен MIT для связи с субъединицей ESCRT-III Vps60. Было показано, что Vta1, хотя и не является обязательным, помогает в сборке кольца Vps4, ускоряет АТФазную активность Vsp4 и способствует разборке ESCRT-III. [ 6 ]
Бро1
[ редактировать ]Основная функция Bro1 — рекрутирование деубиквитиназ в комплекс ESCRT-III. [ 19 ] Это приводит к удалению убиквитиновых меток из белков, предназначенных для деградации в лизосомах, непосредственно перед образованием мультивезикулярных телец. Также предполагалось, что Bro1 помогает стабилизировать ESCRT-III, в то время как метки убиквитина отщепляются от грузовых белков. [ 19 ]
Bro1 содержит аминоконцевой домен Bro1, который связывается с Snf7 ESCRT-III. [ 20 ] Это связывание приводит Bro1 к месту отрыва мембраны. Bro1 также связывает каталитический домен Doa4, убиквитингидролазы (деубиквитиназы), доставляя его к месту отторжения. Doa4 удаляет убиквитин из грузовых белков, направляемых в лизосому. [ 20 ]
Биогенез мультивезикулярного тела и транспортировка грузов
[ редактировать ]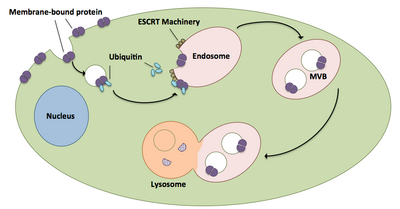
Мультивезикулярные тельца играют большую роль в транспорте убиквитинированных белков и рецепторов к лизосоме. [ 21 ] Комплексы ESCRT транспортируют убиквитинированные грузы в клеточные везикулы, которые отпочковываются непосредственно в эндосомальный компартмент клетки, образуя мультивезикулярные тельца. [ 21 ] Эти мультивезикулярные тельца в конечном итоге сливаются с лизосомой, вызывая деградацию груза. [ 16 ] Более подробное описание процесса, включая связанное с ним оборудование, существует следующим образом:
- Каждый из компонентов ESCRT-0 Vps27 и Hse1 связывается с убиквитинированным грузом. [ 1 ] [ 21 ]
- Vps27 связывается с фосфатидилинозитол-3-фосфатом, эндосомальным липидом, который затем рекрутирует весь комплекс в эндосому. [ 1 ] [ 21 ]
- Vps27 связывает субъединицу Vps23 ESCRT-I, перенося ESCRT-I в эндосому. ESCRT-I также может связывать убиквитинированные белки. [ 1 ] [ 21 ]
- Vps36 связывается с субъединицей Vps28 ESCRT-I, что приводит к рекрутированию комплекса ESCRT-II. [ 1 ]
- Субъединица Vps25 ESCRT-II связывается с Vps20 комплекса ESCRT-III и активирует его. [ 1 ] [ 16 ] [ 21 ]
- Vps20 запускает образование цепей Snf7, которые затем блокируются Vps24. [ 16 ]
- Vps24 набирает Vps2, который привносит в комплекс Vps4. [ 16 ]
- Vps4 образует пору, состоящую из двух гексамерных колец, с которыми связывается Vta1. [ 1 ] Этот комплекс Vps4-Vta1 запускает разборку ESCRT-III и отмечает конец формирования мультивезикулярных телец. [ 2 ]
Отрыв мембраны
[ редактировать ]
Отрыв мембраны во время цитокинеза — это процесс, при котором мембрана, соединяющая две дочерние клетки, расщепляется во время клеточного деления . Поскольку он консервативен у ряда архей , отторжение мембраны считается самой ранней ролью механизма ESCRT. [ 6 ] Процесс начинается, когда центросомный белок Cep55 рекрутируется в среднюю часть делящихся клеток вместе с MKLP1, митотическим кинезин -подобным белком, который связывается с микротрубочками. [ 6 ] [ 23 ] Затем Cep55 рекрутирует субъединицу Vps23 ESCRT-I и вспомогательный белок ALIX, которые образуют кольца по обе стороны от среднего тела. [ 6 ] [ 11 ] [ 12 ] ESCRT-I и ALIX рекрутируют ESCRT-III через его субъединицу Snf7. [ 6 ] Субъединицы ESCRT-III Vps20, Snf7, Vps24, Vps2 и Did2 образуют спиралевидную фибриллу, примыкающую к кольцам, образованным Vps23. [ 1 ] [ 12 ] [ 19 ] Формирование этой спиралевидной структуры деформирует мембрану, и спастин AAA-АТФазы вводится с помощью Did2 и Ist1, чтобы расщепить микротрубочки, образовавшиеся в среднем теле. [ 12 ] [ 19 ] Затем Vps4 катализирует разборку комплекса ESCRT-III, в результате чего образуются две вновь разделенные дочерние клетки. [ 19 ] Процесс отрыва мембраны был описан с использованием белков многоклеточных животных, поскольку этот процесс в большей степени изучен у многоклеточных животных.
Вирусное почкование
[ редактировать ]
Высвобождение вирусных частиц, также известное как вирусное почкование , представляет собой процесс, при котором свободные вирионы высвобождаются изнутри клеток путем захвата механизма ESCRT клетки-хозяина. [ 1 ] [ 14 ] Ретровирусам , таким как ВИЧ-1 и Т-лимфотропный вирус человека , а также ряду вирусов с оболочкой , включая вирус Эбола , требуется механизм ESCRT для выхода из клетки-хозяина. [ 1 ] Процесс инициируется вирусными белками Gag — основными структурными белками ретровирусных оболочек, которые взаимодействуют с TSG101 комплекса ESCRT-I и вспомогательным белком ALIX. [ 12 ] [ 13 ] Субъединицы ESCRT-III (обязательны только CHMP4 и CHMP2) [ 10 ] рекрутируются к месту отпочкования вируса, чтобы сжимать и разрывать шейку зачатка способом, аналогичным тому, который описан для отрыва мембраны во время цитокинеза. [ 1 ] [ 6 ] [ 12 ] Затем Vps4 перерабатывает компоненты ESCRT-III в цитозоль, и вирус высвобождается из клетки. [ 6 ] Описанный здесь механизм использует белки многоклеточных животных, поскольку почкование вирусов более подробно изучалось у многоклеточных животных.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Шмидт О., Тейс Д. (февраль 2012 г.). «Машина ESCRT» . Курс. Биол . 22 (4): Р116–20. дои : 10.1016/j.cub.2012.01.028 . ПМК 3314914 . ПМИД 22361144 .
- ^ Перейти обратно: а б Бабст М. (август 2011 г.). «Формирование пузырьков MVB: ESCRT-зависимый, ESCRT-независимый и все, что между ними» . Курс. Мнение. Клеточная Биол . 23 (4): 452–7. дои : 10.1016/j.ceb.2011.04.008 . ПМК 3148405 . ПМИД 21570275 .
- ^ Перейти обратно: а б с д и ж г час я дж к л Херли Дж. Х., Хэнсон П. И. (август 2010 г.). «Отпочкование и расщепление мембраны с помощью механизма ESCRT: все дело в шее» . Нат. Преподобный мол. Клеточная Биол . 11 (8): 556–66. дои : 10.1038/nrm2937 . ПМК 2922035 . ПМИД 20588296 .
- ^ Самсон, РЮ; Добро, MJ; Дженсен, Дж.Дж.; Белл, SD (2017). «Структура, функции и роли архейного аппарата ESCRT». Прокариотические цитоскелеты . Субклеточная биохимия. Том. 84. С. 357–377. дои : 10.1007/978-3-319-53047-5_12 . ISBN 978-3-319-53045-1 . ПМИД 28500532 .
- ^ Перейти обратно: а б Пайпер Р.К., Кацманн диджей (2007). «Биогенез и функция мультивезикулярных тел» . Анну. Преподобный Cell Dev. Биол . 23 : 519–47. doi : 10.1146/annurev.cellbio.23.090506.123319 . ПМЦ 2911632 . ПМИД 17506697 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т Херли Дж. Х. (декабрь 2010 г.). «Комплексы ЭСКРТ» . Крит. Преподобный Биохим. Мол. Биол . 45 (6): 463–87. дои : 10.3109/10409238.2010.502516 . ПМЦ 2988974 . ПМИД 20653365 .
- ^ Уоллерт Т., Херли Дж. Х. (апрель 2010 г.). «Молекулярный механизм биогенеза мультивезикулярных тел комплексами ESCRT» . Природа . 464 (7290): 864–9. Бибкод : 2010Natur.464..864W . дои : 10.1038/nature08849 . ПМЦ 2851844 . ПМИД 20305637 .
- ^ Рен Икс, Херли Дж. Х. (март 2010 г.). «VHS-домены ESCRT-0 взаимодействуют в высокоавидном связывании с полиубиквитинированным грузом» . ЭМБО Дж . 29 (6): 1045–54. дои : 10.1038/emboj.2010.6 . ПМЦ 2845278 . ПМИД 20150893 .
- ^ Банерджи С., Басу С., Саркар С. (2010). «Сравнительная геномика выявляет избирательное распределение и доменную организацию белков доменов FYVE и PX в эукариотических линиях» . БМК Геномика . 11:83 . дои : 10.1186/1471-2164-11-83 . ПМЦ 2837644 . ПМИД 20122178 .
- ^ Перейти обратно: а б Морита Э., Сандрин В., Маккалоу Дж., Кацуяма А., Бачи Гамильтон I, Сандквист В.И. (март 2011 г.). «Требования к белку ESCRT-III для зачатия ВИЧ-1» . Микроб-хозяин клетки . 9 (3): 235–42. дои : 10.1016/j.chom.2011.02.004 . ПМК 3070458 . ПМИД 21396898 .
- ^ Перейти обратно: а б с д и ж г час Аделл М.А., Тейс Д. (октябрь 2011 г.). «Сборка и разборка мембранного расщепляющегося комплекса ESCRT-III» . ФЭБС Летт . 585 (20): 3191–6. дои : 10.1016/j.febslet.2011.09.001 . ПМК 3192940 . ПМИД 21924267 .
- ^ Перейти обратно: а б с д и ж г час Мюллер М., Аделл М.А., Тейс Д. (август 2012 г.). «Отрыв мембраны: первый взгляд на динамические ESCRT» . Курс. Биол . 22 (15): Р603–5. дои : 10.1016/j.cub.2012.06.063 . ПМЦ 3414845 . ПМИД 22877781 .
- ^ Перейти обратно: а б с Макдональд Б., Мартин-Серрано Дж. (июль 2009 г.). «Без каких-либо условий: механизм ESCRT в почковании вируса и цитокинезе» . Дж. Клеточная наука . 122 (Часть 13): 2167–77. дои : 10.1242/jcs.028308 . ПМЦ 2723143 . ПМИД 19535732 .
- ^ Перейти обратно: а б Жувене Н., Жадина М., Бениас П.Д. , Симон С.М. (апрель 2011 г.). «Динамика рекрутирования белка ESCRT во время сборки ретровируса» . Нат. Клеточная Биол . 13 (4): 394–401. дои : 10.1038/ncb2207 . ПМК 3245320 . ПМИД 21394083 .
- ^ Глотцер, Майкл. «Цитокинез: лунный свет Центрального шпидлина как мембранный якорь» , Current Biology , 18 февраля 2013 г.
- ^ Перейти обратно: а б с д и Тейс Д., Саксена С., Джадсон Б.Л., Эмр С.Д. (март 2010 г.). «ESCRT-II координирует сборку нитей ESCRT-III для сортировки грузов и формирования мультивезикулярных везикул тела» . ЭМБО Дж . 29 (5): 871–83. дои : 10.1038/emboj.2009.408 . ПМЦ 2837172 . ПМИД 20134403 .
- ^ Скотт А., Гаспар Дж., Стучелл-Бреретон, доктор медицинских наук, Алам С.Л., Скалицкий Дж.Дж., Сундквист В.И. (сентябрь 2005 г.). «Структура и взаимодействие белков ESCRT-III домена MIT человеческого VPS4A» . Учеб. Натл. акад. наук. США . 102 (39): 13813–8. Бибкод : 2005PNAS..10213813S . дои : 10.1073/pnas.0502165102 . ПМЦ 1236530 . ПМИД 16174732 .
- ^ Азми И., Дэвис Б., Димаано С., Пейн Дж., Эккерт Д., Бабст М., Кацманн DJ (февраль 2006 г.). «Переработка ESCRT с помощью AAA-АТФазы Vps4 регулируется консервативной областью VSL в Vta1» . Дж. Клеточная Биол . 172 (5): 705–17. дои : 10.1083/jcb.200508166 . ПМК 2063703 . ПМИД 16505166 .
- ^ Перейти обратно: а б с д и Бабст М., Дэвис Б.А., Кацманн DJ (октябрь 2011 г.). «Регуляция Vps4 во время сортировки MVB и цитокинеза» . Трафик . 12 (10): 1298–305. дои : 10.1111/j.1600-0854.2011.01230.x . ПМК 3171586 . ПМИД 21658171 .
- ^ Перейти обратно: а б Веммер М., Азми И., Вест М., Дэвис Б., Кацманн Д., Одорицци Г. (январь 2011 г.). «Связывание Bro1 с Snf7 регулирует активность разрыва мембраны ESCRT-III у дрожжей» . Дж. Клеточная Биол . 192 (2): 295–306. дои : 10.1083/jcb.201007018 . ПМК 3172170 . ПМИД 21263029 .
- ^ Перейти обратно: а б с д и ж Херли Дж. Х., Эмр С.Д. (2006). «Комплексы ESCRT: структура и механизм мембранной транспортной сети» . Annu Rev Biophys Biomol Struct . 35 : 277–98. doi : 10.1146/annurev.biophys.35.040405.102126 . ПМК 1648078 . ПМИД 16689637 .
- ^ Кармена М. (июль 2012 г.). «Контроль контрольно-пропускного пункта отрыва: застрял посередине с Авророй Б» . Открытая Биол . 2 (7): 120095. doi : 10.1098/rsob.120095 . ПМК 3411112 . ПМИД 22870391 .
- ^ Чжу С., Босси-Ветцель Э., Цзян В. (июль 2005 г.). «Привлечение MKLP1 в среднюю зону/среднее тело веретена с помощью INCENP имеет важное значение для формирования среднего тела и завершения цитокинеза в клетках человека» . Биохим. Дж . 389 (Часть 2): 373–81. дои : 10.1042/BJ20050097 . ПМК 1175114 . ПМИД 15796717 .