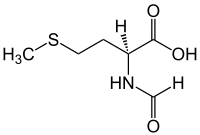
N- формилметионин
 | |
| Имена | |
|---|---|
| Название ИЮПАК N- формилметионин | |
| Систематическое название ИЮПАК ( S )-2-формиламино-4-метилсульфанилбутановая кислота | |
| Другие имена 2-формиламино-4-метилсульфанилмасляная кислота; формилметионин; N -формил(метил)гомоцистеин | |
| Идентификаторы | |
3D model ( JSmol ) | |
| Сокращения | fMet |
| ЧЭБИ | |
| ХимическийПаук | |
| Номер ЕС |
|
ПабХим CID | |
| НЕКОТОРЫЙ | |
| Характеристики | |
| С 6 Н 11 Н О 3 С | |
| Молярная масса | 177.22 g/mol |
| Опасности | |
| СГС Маркировка : [1] | |
 | |
| Предупреждение | |
| H319 | |
| П264+П265 , П280 , П305+П351+П338 , П337+П317 | |
| Страница дополнительных данных | |
| N-формилметионин (страница данных) | |
Если не указано иное, данные приведены для материалов в стандартном состоянии (при 25 °C [77 °F], 100 кПа). | |
N -формилметионин (fMet, [2] ОХО-Мет, [3] Для-Met [3] ) представляет собой производное аминокислоты метионина , в котором формильная добавлена к аминогруппе группа . Он специально используется для инициации синтеза белка из бактериальных и органеллярных генов и может быть удален посттрансляционно .
fMet играет решающую роль в синтезе белков бактерий, митохондрий и хлоропластов . Он не используется в цитозольного синтезе белка эукариот , где гены транслируются ядерные эукариот . Он также не используется археями . В организме человека fMet распознается иммунной системой как чужеродный материал или как сигнал тревоги, испускаемый поврежденными клетками, и стимулирует организм на борьбу с потенциальной инфекцией.
Функция в синтезе белка
[ редактировать ]Перевод
[ редактировать ]fMet необходим для эффективной инициации синтеза белка у большинства групп бактерий. Комплекс 30S рибосома-мРНК специфически рекрутирует тРНК с формилированной аминокислотой – тРНК. fMet прикреплен к fMet в естественном случае. [4]
Поскольку fMet управляет инициацией, белки бактерий начинаются ( N -конец ) с остатка fMet вместо метионина. Дальнейшее появление кодона «AUG» приведет к образованию нормального метионина, поскольку нормальная «удлиняющаяся» тРНК Из используется. [4]
Присоединение формильной группы к метионину катализируется ферментом метионил -тРНК формилтрансферазой . Эта модификация осуществляется после загрузки метионина на тРНК. fMet с помощью аминоацил-тРНК-синтетазы . Сам метионин может быть загружен либо на тРНК, либо на тРНК. fMet или тРНК Из . Однако формилтрансфераза будет катализировать присоединение формильной группы к метионину только в том случае, если метионин загружен на тРНК. fMet , а не на тРНК Из . Это связано с тем, что формилтрансфераза распознает специфические особенности тРНК. fMet . [4]
Митохондрии клеток, в том эукариотических . числе клеток человека, и клеток также хлоропласты растительных инициируют синтез белка с помощью fMet Учитывая, что митохондрии и хлоропласты имеют общий с бактериями первоначальный синтез белка с fMet, это было названо доказательством эндосимбиотической теории . [5]
Неожиданно формилтрансфераза также может действовать на тРНК эукариотического инициатора в живых дрожжевых клетках. Даже в нормальных условиях формилтрансфераза, кодируемая ядром, не полностью импортируется в митохондрии; еще больше остается в цитозоле при стрессе. Эти цитозольные формилтрансферазы продуцируют fMet-тРНК i , которая может использоваться цитозольными рибосомами для производства белков с N-концевым fMet. Эти белки подвергаются деградации в результате специфических процессов в клетке. [6]
Дальнейшая обработка
[ редактировать ]N - концевой fMet удаляется из большинства белков, как хозяина, так и рекомбинантных, посредством последовательности двух ферментативных реакций. Сначала пептиддеформилаза (PDF) деформирует его, превращая остаток обратно в нормальный метионин. Затем метионинаминопептидаза (MetAP) удаляет остаток из цепи. [7] MetAP действует только на белки с остатками во втором положении, которые менее объемны, чем валин. [8]
N - концевой fMet, если он не удаляется PDF, по-видимому, действует как дегрон , сигнал к деградации белка. [8]
Вариация
[ редактировать ]Формильная группа не является строго необходимой для инициирования. Бактерии с нокаутом формилтрансферазы, что предотвращает появление Met-тРНК. fMet (т.е. метионин загружен на тРНК fMet ) от превращения в fMet-тРНК fMet , могут иметь различную степень остаточной способности начинать синтез белка. E. coli , S. pneumoniae и B. subtilis почти не демонстрируют сохраняющейся способности к трансляции, в то время как P. aeruginosa , S. aureus , H. influenzae и, возможно, S. faecalis все еще производят большое количество белка. У P. aeruginosa этой способности способствует бактериальный фактор инициации 2 , который может нести как Met-тРНК, так и fMet и fMet-тРНК fMet к рибосоме. [9]
Актуальность для иммунологии
[ редактировать ]Поскольку fMet присутствует в белках, вырабатываемых бактериями, но не в белках, вырабатываемых эукариотами (за исключением органелл бактериального происхождения), иммунная система может использовать его, чтобы отличить свое от чужого. Полиморфно-ядерные клетки могут связывать белки, начиная с fMet, и использовать их для инициирования привлечения лейкоцитов циркулирующей крови , а затем стимулировать микробицидную активность, такую как фагоцитоз . [10] [11] [12]
Поскольку fMet присутствует в белках, вырабатываемых митохондриями и хлоропластами, более поздние теории не рассматривают его как молекулу, которую иммунная система может использовать, чтобы отличать свое от чужого. [13] Вместо этого fMet-содержащие олигопептиды и белки, по-видимому, высвобождаются митохондриями поврежденных тканей, а также поврежденными бактериями и, таким образом, могут квалифицироваться как «сигнал тревоги», как обсуждается в « Опасность модели иммунитета ». Прототипическим fMet-содержащим олигопептидом является N -формилметионин-лейцил-фенилаланин этих клеток, (FMLP), который активирует лейкоциты и другие типы клеток путем связывания с рецепторами формилпептида 1 (FPR1) и рецептором формилпептида 2 (FPR2) связанными с G-белком ( см. также рецептор формилпептида 3 ). Действуя через эти рецепторы, fMet-содержащие олигопептиды и белки являются частью врожденной иммунной системы ; они функционируют, инициируя острые воспалительные реакции, но в других условиях действуют, чтобы ингибировать и разрешать эти реакции. fMet-содержащие олигопептиды и белки также участвуют в других физиологических и патологических реакциях.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «N-формил-DL-метионин» . pubchem.ncbi.nlm.nih.gov .
- ^ ПабХим. «N-формил-DL-метионин» . pubchem.ncbi.nlm.nih.gov . Проверено 24 октября 2020 г.
- ^ Jump up to: а б Номенклатура и символика аминокислот и пептидов 3AA-18 и 3AA-19.
- ^ Jump up to: а б с Шетти, С; Шах, РА; Чембажи, УФ; Сах, С; Варшней, У (28 февраля 2017 г.). «Две высококонсервативные особенности бактериальных инициаторных тРНК позволяют им проходить через отдельные контрольные точки при инициации трансляции» . Исследования нуклеиновых кислот . 45 (4): 2040–2050. дои : 10.1093/nar/gkw854 . ПМК 5389676 . ПМИД 28204695 .
- ^ Альбертс, Брюс (18 ноября 2014 г.). Молекулярная биология клетки (Шестое изд.). Нью-Йорк, штат Нью-Йорк. п. 800. ISBN 978-0-8153-4432-2 . OCLC 887605755 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Варшавский Александр (8 января 2019 г.). «Пути N-дегрона и C-дегрона деградации белка» . Труды Национальной академии наук . 116 (2): 358–366. Бибкод : 2019PNAS..116..358V . дои : 10.1073/pnas.1816596116 . ПМК 6329975 . ПМИД 30622213 .
- ^ Шерман Ф., Стюарт Дж.В., Цунасава С. (июль 1985 г.). «Метионин или не метионин в начале белка». Биоэссе . 3 (1): 27–31. дои : 10.1002/bies.950030108 . ПМИД 3024631 . S2CID 33735710 .
- ^ Jump up to: а б Пятков, К.И.; Ву, ТТ; Хван, CS; Варшавский, А (2015). «Формилметионин как сигнал деградации на N-концах бактериальных белков» . Microbial Cell (Грац, Австрия) . 2 (10): 376–393. дои : 10.15698/mic2015.10.231 . ПМЦ 4745127 . ПМИД 26866044 .
- ^ Пятков, К.И.; Ву, ТТ; Хван, CS; Варшавский, А (2015). «Формилметионин как сигнал деградации на N-концах бактериальных белков» . Microbial Cell (Грац, Австрия) . 2 (10): 376–393. дои : 10.15698/mic2015.10.231 . ПМЦ 4745127 . ПМИД 26866044 .
- ^ Иммунология на MCG 1/фагшаг
- ^ «Врожденная иммунная система: рецепторы распознавания образов, антиген-неспецифические антимикробные молекулы тела и цитокины» . Архивировано из оригинала 27 июля 2010 г.
- ^ Детмерс П.А., Райт С.Д., Олсен Э., Кимбалл Б., Кон З.А. (сентябрь 1987 г.). «Агрегация рецепторов комплемента на нейтрофилах человека в отсутствие лиганда» . Журнал клеточной биологии . 105 (3): 1137–45. дои : 10.1083/jcb.105.3.1137 . ПМК 2114803 . ПМИД 2958480 .
- ^ Чжан Ц, Рауф М, Чен Ю, Суми Ю, Сурсал Т, Юнгер В, Брохи К, Итагаки К, Хаузер СиДжей (4 марта 2010 г.). «Циркулирующие митохондриальные DAMP вызывают воспалительные реакции на повреждение» . Природа . 464 (7285): 104–107. Бибкод : 2010Natur.464..104Z . дои : 10.1038/nature08780 . ПМЦ 2843437 . ПМИД 20203610 .
Внешние ссылки
[ редактировать ]- N-формилметионин в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)