Сульфатные черенки
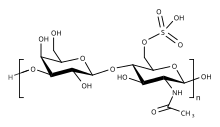
Кератансульфат ( KS ), также называемый кератосульфатом , представляет собой один из нескольких сульфатированных гликозаминогликанов (структурных углеводов), которые обнаружены особенно в роговице , хрящах и костях . Он также синтезируется в центральной нервной системе , где участвует как в развитии [ 1 ] и при формировании глиального рубца после травмы. [ 2 ] Кератансульфаты представляют собой крупные, сильно гидратированные молекулы, которые в суставах могут выступать в качестве подушки, поглощающей механические удары .
Структура
[ редактировать ]Как и другие гликозаминогликаны, кератансульфат представляет собой линейный полимер , состоящий из повторяющихся дисахаридных звеньев. Кератансульфат представляет собой протеогликан (PG), в котором цепи KS прикреплены к белкам клеточной поверхности или внеклеточного матрикса , называемым коровыми белками. Коровые белки KS включают люмикан , кератокан , мимекан , фибромодулин , PRELP , остеоадгерин и аггрекан .
Основной повторяющейся дисахаридной единицей в составе кератансульфата является -3 Gal β1-4 GlcNAc6S β1-. Он может быть сульфатирован по положению углерода 6 (C6) одного или обоих моносахаридов Gal или GlcNAc . Однако детальную первичную структуру конкретных типов КС лучше всего рассматривать как состоящую из трех областей: [ 3 ]
- Область связи, на одном конце которой цепь KS связана с основным белком.
- Область повтора, состоящая из повторяющейся дисахаридной единицы -3 Gal β1-4 GlcNAc β1 и
- Область, закрывающая цепь, расположенная на конце цепи KS, противоположном области связывания белка.
Моносахарид манноза находится в области связи кератансульфата типа I (KSI). Дисахариды в повторяющейся области KSII могут быть фукозилированы , а N-ацетилнейраминовая кислота блокирует концы всех цепей кератансульфата типа II (KSII) и до 70% цепей типа KSI. [ 4 ]
занятия КС
[ редактировать ]Обозначения KSI и KSII первоначально были присвоены в зависимости от типа ткани, из которой был выделен кератансульфат. KSI выделяли из роговицы ткани , а KSII — из скелетной ткани. [ 5 ] [ 6 ] Незначительные различия в составе моносахаридов существуют между KS, экстрагированным из обоих источников, и даже между KS, экстрагированным из одного и того же источника. Однако основные различия происходят в том, как каждый тип KS соединяется со своим основным белком. [ 7 ] Обозначения KSI и KSII теперь основаны на этих различиях в белковых связях. KSI N -связан со специфическими аспарагина аминокислотами через N -ацетилглюкозамин , а KSII O -связан со специфическими серина или треонина аминокислотами через N -ацетилгалактозамин . [ 8 ] Тканевой классификации СК больше не существует, поскольку было показано, что типы СК неспецифичны для тканей. [ 4 ] Третий тип KS (KSIII) также был выделен из ткани головного мозга , которая O -связана со специфическими аминокислотами серином или треонином через маннозу . [ 9 ]
Роговичник КСИ
[ редактировать ]Количество КС в роговице в 10 раз выше, чем в хряще и в 2-4 раза выше, чем в других тканях. [ 10 ] Его производят кератоциты роговицы. [ 11 ] Считается, что он играет роль динамического буфера гидратации роговицы. При редком прогрессирующем заболевании, называемом макулярной дистрофией роговицы (MCDC), синтез кератансульфата либо отсутствует (MCDC типа I), либо нарушен (MCDC типа II). [ 12 ]
Нероговичный КСИ
[ редактировать ]Остеоадгерин , фибромодулин и PRELP — это основные белки костей и хрящей, модифицированные N-связанными KS-цепями. Цепи KS, связанные с остеоадгерином и фибромодулином, короче, чем цепи, обнаруженные в роговице, обычно составляют 8-9 дисахаридных единиц в длину. [ 13 ] Тогда как KSI роговицы состоит из ряда доменов с различной степенью сульфатации, самый длинный из которых может иметь длину 8-32 дисахаридных единиц. Невосстанавливающий конец фибромодулина KS по структуре больше похож на невосстанавливающий конец кератансульфата типа KSII, чем на роговичный KSI. Таким образом, считается, что структура KS определяется тканеспецифичной доступностью гликозилтрансфераз, а не типом связи с коровым белком. [ 4 ]
КСИИ
[ редактировать ]Хрящ KSII почти полностью сульфатирован и состоит из дисульфатированных мономеров , иногда прерываемых одним моносульфатированным мономером лактозамина . [ 8 ] Фукозилирование также часто встречается при наличии альфа-связанной фукозы в положении углерода 3 сульфатированного GlcNAc , за исключением случая трахеального KSII, где этот признак отсутствует.
См. также
[ редактировать ]- N-ацетилглюкозамин
- Галактоза
- Лумикан
- Синдром Моркио характеризуется накоплением КС.
- Протеогликан 4
Ссылки
[ редактировать ]- ^ Миллер Б., Шеппард А.М., Перлман А.Л. (апрель 1997 г.). «Экспрессия кератансульфатподобной иммунореактивности в процессе развития отличает ядра таламуса и корковые домены». Дж. Комп. Нейрол. 380 (4): 533–52. doi : 10.1002/(SICI)1096-9861(19970421)380:4<533::AID-CNE9>3.0.CO;2-2 . ПМИД 9087531 .
- ^ Чжан Х., Учимура К., Кадомацу К. (ноябрь 2006 г.). «Кератансульфат головного мозга и образование глиальных рубцов». Энн. Н-Й акад. наук. 1086 (1): 81–90. Бибкод : 2006NYASA1086...81Z . дои : 10.1196/анналы.1377.014 . ПМИД 17185507 .
- ^ Тай Г.Х., Хакерби Т.Н., Недушинский И.А. (1996). «Множественные концы невосстанавливающих цепей, выделенные из кератансульфатов роговицы крупного рогатого скота» . Ж. Биол. Хим . 271 (38): 23535–23546. дои : 10.1074/jbc.271.38.23535 . ПМИД 8798563 .
- ^ Jump up to: а б с Фундербург Дж.Л. (2000). «Кератансульфат: строение, биосинтез и функции» . Гликобиология . 10 (10): 951–958. дои : 10.1093/гликоб/10.10.951 . ПМИД 11030741 .
- ^ Мейер К., Линкер А. и др. (1 декабря 1953 г.). «Мукополисахариды бычьей роговицы» . Ж. Биол. Хим . 205 (2): 611–616. дои : 10.1016/S0021-9258(18)49203-4 . ПМИД 13129238 .
- ^ Мейер К; Хоффман П.; Линкер А. (1958). «Мукополисахариды реберного хряща». Наука . 128 (3329): 896. Бибкод : 1958Sci...128..896M . дои : 10.1126/science.128.3329.896 . ПМИД 13592269 .
- ^ Сено Н., Мейер К. и др. (1 марта 1965 г.). «Вариации кератосульфатов» . Ж. Биол. Хим . 240 (3): 1005–1019. дои : 10.1016/S0021-9258(18)97528-9 . ПМИД 14284693 .
- ^ Jump up to: а б Недушинский И.А., Хакерби Т.Н. и др. (1990). «Существует два основных типа скелетных кератансульфатов» . Биохим. Дж . 271 (1): 243–245. дои : 10.1042/bj2710243 . ПМЦ 1149539 . ПМИД 2222415 .
- ^ Крузиус Т., Финн Дж. и др. (25 июня 1986 г.). «Идентификация O-гликозидного маннозо-связанного сиалилированного тетрасахарида и кератансульфатолигосахаридов в хондроитинсульфат-протеогликанах мозга» . Ж. Биол. Хим . 261 (18): 8237–8242. дои : 10.1016/S0021-9258(19)83901-7 . ПМИД 2941416 .
- ^ Фундербург Дж.Л.; Катерсон Б.; Конрад ГВ. (1987). «Распределение протеогликанов, антигенно связанных с протеогликанами кератансульфата роговицы» . Ж. Биол. Хим . 262 (24): 11634–11640. дои : 10.1016/S0021-9258(18)60856-7 . ПМИД 2957372 .
- ^ Фундербург Дж.Л., Манн М.М., Фундербург М.Л. (ноябрь 2003 г.). «Фенотип кератоцитов опосредует структуру протеогликана: роль фибробластов в фиброзе роговицы» . Ж. Биол. хим. 278 (46): 45629–37. дои : 10.1074/jbc.M303292200 . ПМЦ 2877919 . ПМИД 12933807 .
- ^ Макулодистрофия роговицы, 1 - OMIM
- ^ Лаудер Р.М., Хакерби Т.Н., Недушинский И.А. (1997). «Структура цепей кератансульфата, прикрепленных к фибромодулину суставного хряща человека». Гликокондж. Дж . 14 (5): 651–660. дои : 10.1023/А:1018552913584 . ПМИД 9298700 .
Внешние ссылки
[ редактировать ]- Хондроитин и кератинсульфат в Университета Дьюка программе ортопедии
- Кератан + сульфат Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)