охранная камера

Замыкающие клетки — это специализированные растительные клетки в эпидермисе листьев, стеблей и других органов, которые используются для контроля газообмена. Они образуются парами с промежутком между ними, образующим устьичную пору . Устьичные поры являются самыми большими, когда вода находится в свободном доступе и замыкающие клетки становятся набухшими , и закрываются, когда доступность воды критически низкая и замыкающие клетки становятся вялыми. Фотосинтез зависит от диффузии ( углекислого газа СО 2 ) из воздуха через устьица в ткани мезофилла . Кислород (O 2 ), образующийся как побочный продукт фотосинтеза, выходит из растения через устьица. Когда устьица открыты, вода теряется за счет испарения и должна быть заменена транспирационным потоком водой, поглощаемой корнями. Растения должны сбалансировать количество поглощенного из воздуха CO 2 с потерей воды через устьичные поры, что достигается как активным, так и пассивным контролем тургорного давления замыкающих клеток и размера устьичных пор. [ 1 ] [ 2 ] [ 3 ] [ 4 ]
Функция защитных клеток
[ редактировать ]Замыкающие клетки – это клетки, окружающие каждую стому . Они помогают регулировать скорость транспирации, открывая и закрывая устьица. Свет является основным триггером открытия или закрытия. Каждая замыкающая клетка имеет относительно толстую и более тонкую кутикулу со стороны поры и тонкую — напротив нее. Когда вода попадает в клетку, тонкая сторона выпячивается наружу, как воздушный шарик, и тянет за собой толстую сторону, образуя полумесяц; комбинированные полумесяцы образуют отверстие поры.
Замыкающие клетки содержат фототропиновые белки, которые представляют собой сериновые и треониновые киназы с фоторецепторной активностью синего света. Фототрофины содержат два домена датчиков света, кислорода и напряжения (LOV) и являются частью суперсемейства доменов PAS . [ 5 ] Фототропины вызывают множество реакций, таких как фототропизм, движение хлоропластов и расширение листьев, а также открытие устьиц. [ 5 ] Примерно до 1998 года было мало что известно о том, как работают эти фоторецепторы. Механизм действия фототропинов был выяснен в ходе экспериментов с бобами ( Vicia faba ). Иммунодетекция и далеко-вестерн-блоттинг показали, что синий свет возбуждает фототропин 1 и фототропин 2, заставляя протеинфосфатазу 1 запускать каскад фосфорилирования, который активирует H. + -АТФаза, насос, ответственный за перекачку H + ионы выходят из клетки. [ 3 ] Фосфорилированный H + -АТФаза позволяет связывать белок 14-3-3 с аутоингибирующим доменом H. + -АТФаза на С-конце. [ 6 ] Затем серин и треонин фосфорилируются внутри белка, что индуцирует H. + -АТФазная активность. [ 5 ] Тот же эксперимент также показал, что при фосфорилировании белок 14-3-3 связывался с фототропинами до того, как H + -АТФаза фосфорилирована. [ 5 ] В аналогичном эксперименте они пришли к выводу, что связывание белка 14-3-3 с сайтом фосфорилирования необходимо для активации плазматической мембраны H. + -АТФазная активность. [ 6 ] Это было сделано путем добавления фосфопептидов, таких как P-950, который ингибирует связывание белка 14-3-3, к фосфорилированному H. + -АТФаза и соблюдение аминокислотной последовательности. По мере откачки протонов на плазматической мембране образовывался отрицательный электрический потенциал. Эта гиперполяризация мембраны способствовала накоплению заряженного калия (K + ) ионы и хлорид (Cl − ) ионы, что, в свою очередь, увеличивает концентрацию растворенного вещества, вызывая снижение водного потенциала. Отрицательный водный потенциал позволяет происходить осмосу в замыкающей клетке, в результате чего вода попадает внутрь, позволяя клетке становиться набухшей. [ нужна ссылка ]
Открытие и закрытие устьичной поры опосредуется изменениями тургорного давления двух замыкающих клеток. Тургорное давление замыкающих клеток контролируется перемещением больших количеств ионов и сахаров в замыкающие клетки и из них. Замыкающие клетки имеют клеточную стенку различной толщины (ее внутренняя область, прилегающая к устьичной поре, более толстая и сильно кутинизированная). [ 7 ] ) и по-разному ориентированные целлюлозные микроволокна, заставляющие их изгибаться наружу, когда они становятся набухшими, что, в свою очередь, приводит к открытию устьиц. Устьица закрываются, когда происходит осмотическая потеря воды, происходящая из-за потери К. + соседним клеткам, главным образом калию (K + ) ионы. [ 8 ] [ 9 ] [ 10 ]
Потери воды и эффективность использования воды
[ редактировать ]Водный стресс (засуха и солевой стресс) является одной из основных экологических проблем, вызывающих серьезные потери в сельском хозяйстве и природе. Устойчивость растений к засухе опосредована несколькими механизмами, которые работают вместе, включая стабилизацию и защиту растения от повреждений, вызванных высыханием, а также контроль того, сколько воды растения теряют через устьичные поры во время засухи. Растительный гормон абсцизовая кислота (АБК) вырабатывается в ответ на засуху. Идентифицирован основной тип рецептора АБК. [ 11 ] [ 12 ] Растительный гормон АБК вызывает закрытие устьичных пор в ответ на засуху, что снижает потерю воды растениями за счет транспирации в атмосферу и позволяет растениям избегать или замедлять потерю воды во время засухи. Использование засухоустойчивых сельскохозяйственных культур приведет к снижению потерь урожая во время засухи. [ нужна ссылка ] Поскольку замыкающие клетки контролируют потерю воды растениями, исследование того, как регулируется открытие и закрытие устьиц, может привести к развитию растений, которые будут лучше избегать или замедлять высыхание и повышать эффективность использования воды . [ 1 ]
АБК является триггером закрытия устьичного отверстия. Чтобы вызвать это, он активирует высвобождение анионов и ионов калия. Этот приток анионов вызывает деполяризацию плазматической мембраны. Эта деполяризация приводит к тому, что ионы калия плюс в клетке покидают клетку из-за дисбаланса мембранного потенциала. Это внезапное изменение концентрации ионов приводит к сокращению замыкающей клетки, что приводит к закрытию устьиц, что, в свою очередь, уменьшает количество потерянной воды. Согласно его исследованиям, все это — цепная реакция. Увеличение АБК приводит к увеличению концентрации ионов кальция. Хотя сначала они думали, что это совпадение, позже они обнаружили, что увеличение количества кальция важно. Они обнаружили, что ионы Ca2+ участвуют в активации анионных каналов, что позволяет анионам проникать в замыкающую клетку. Они также участвуют в предотвращении коррекции протонной АТФазы и предотвращении деполяризации мембраны. Чтобы подтвердить свою гипотезу о том, что кальций ответственен за все эти изменения в клетке, они провели эксперимент, в котором использовали белки, ингибирующие выработку ионов кальция. Если бы они предположили, что кальций важен в этих процессах, они бы увидели, что с ингибиторами они бы увидели меньше следующих вещей. Их предположение оказалось верным, и когда были использованы ингибиторы, они увидели, что протонная АТФаза лучше работает, чтобы сбалансировать деполяризацию. Они также обнаружили, что поток анионов в замыкающие клетки был не таким сильным. Это важно для того, чтобы ионы попадали в замыкающую клетку. Эти две вещи имеют решающее значение для закрытия устьичного отверстия и предотвращения потери воды растением. [ 13 ]
Поглощение и высвобождение ионов
[ редактировать ]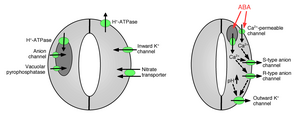
Поглощение ионов замыкающими клетками вызывает открытие устьиц: открытие газообменных пор требует поглощения ионов калия замыкающими клетками. Были идентифицированы калиевые каналы и насосы, которые участвуют в поглощении ионов и открытии устьичных отверстий. [ 1 ] [ 14 ] [ 15 ] [ 16 ] [ 17 ] [ 18 ] [ 19 ] [ 20 ] Высвобождение ионов из замыкающих клеток вызывает закрытие устьичных пор: были идентифицированы другие ионные каналы, которые опосредуют высвобождение ионов из замыкающих клеток, что приводит к осмотическому оттоку воды из замыкающих клеток вследствие осмоса , сокращению замыкающих клеток и закрытию устьичных пор ( рисунки 1 и 2). Специализированные каналы оттока калия участвуют в высвобождении калия из замыкающих клеток. [ 16 ] [ 21 ] [ 22 ] [ 23 ] [ 24 ] Анионные каналы были идентифицированы как важные контроллеры закрытия устьиц. [ 25 ] [ 26 ] [ 27 ] [ 28 ] [ 29 ] [ 30 ] [ 31 ] Анионные каналы выполняют несколько основных функций в контроле закрытия устьиц: [ 26 ] (а) Они позволяют высвобождать анионы, такие как хлорид и малат, из замыкающих клеток, что необходимо для закрытия устьиц. (б) Анионные каналы активируются сигналами, которые вызывают закрытие устьиц, например, внутриклеточным кальцием и АБК. [ 26 ] [ 29 ] [ 32 ] В результате высвобождение отрицательно заряженных анионов из замыкающих клеток приводит к электрическому сдвигу мембраны в сторону более положительных напряжений ( деполяризация ) на внутриклеточной поверхности плазматической мембраны замыкающих клеток . Эта электрическая деполяризация замыкающих клеток приводит к активации внешних калиевых каналов и высвобождению калия через эти каналы. В плазматической мембране охарактеризованы по крайней мере два основных типа анионных каналов: анионные каналы S-типа и анионные каналы R-типа. [ 25 ] [ 26 ] [ 28 ] [ 33 ]
Вакуольный транспорт ионов
[ редактировать ]Вакуоли — это крупные внутриклеточные запасающие органеллы в клетках растений. Помимо ионных каналов в плазматической мембране, вакуолярные ионные каналы выполняют важные функции в регуляции открытия и закрытия устьиц, поскольку вакуоли могут занимать до 90% объема замыкающих клеток. Следовательно, большая часть ионов высвобождается из вакуолей, когда устьица закрыты. [ 34 ] Сосудистый К + (VK) каналы и быстрые вакуолярные каналы могут опосредовать K + освобождение из вакуолей. [ 35 ] [ 36 ] [ 37 ] Вакуолярный К + (VK)-каналы активируются при повышении внутриклеточной концентрации кальция. [ 35 ] Другим типом активируемых кальцием каналов является медленный вакуолярный (SV) канал. [ 38 ] Было показано, что каналы SV функционируют как катионные каналы, проницаемые для Ca. 2+ ионы, [ 35 ] но их точные функции у растений еще не известны. [ 39 ]
Защитные клетки контролируют газообмен и ионный обмен посредством открытия и закрытия. K+ — это один ион, который течет как в клетку, так и из нее, вызывая образование положительного заряда. Малат является одним из основных анионов, используемых для противодействия этому положительному заряду, и он перемещается через ионный канал AtALMT6. [ 40 ] AtALMT6 представляет собой активируемый алюминием переносчик малата, который обнаруживается в замыкающих клетках, особенно в вакуолях. Было обнаружено, что этот транспортный канал вызывает либо приток, либо отток малата в зависимости от концентрации кальция. [ 40 ] В исследовании Meyer et al. эксперименты «пэтч-кламп» были проведены на мезофильных вакуолях арабидопсиса rdr6-11 (WT) и арабидопсиса, которые сверхэкспрессировали AtALMT6-GFP. [ 40 ] В результате этих экспериментов было обнаружено, что в WT наблюдались лишь небольшие токи при введении ионов кальция, тогда как у мутанта AtALMT6-GFP наблюдался огромный направленный внутрь выпрямляющий ток. [ 40 ] Когда транспортер выбивается из вакуолей замыкающих клеток, ток потока малата значительно снижается. Ток меняется от огромного внутреннего тока до почти не отличающегося от тока WT, и Мейер и др. предположили, что это происходит из-за остаточных концентраций малата в вакуоли. [ 40 ] У нокаутных мутантов также наблюдается аналогичная реакция на засуху, как и у WT. Не наблюдалось фенотипических различий между нокаутными мутантами, диким типом или мутантами AtALMT6-GFP, и точная причина этого полностью не известна. [ 40 ]
Преобразование сигнала
[ редактировать ]Замыкающие клетки воспринимают и обрабатывают внешние и эндогенные стимулы, такие как свет, влажность, концентрация CO 2 , температура, засуха и растительные гормоны, чтобы вызвать клеточные реакции, приводящие к открытию или закрытию устьиц. Эти пути передачи сигналов определяют, например, насколько быстро растение будет терять воду в период засухи. Защитные клетки стали моделью передачи сигналов одной клетки. С помощью Arabidopsis thaliana исследование обработки сигналов в одиночных замыкающих клетках стало доступным для генетики . [ 29 ] Были идентифицированы цитозольные и ядерные белки и химические мессенджеры, которые участвуют в движениях устьиц и опосредуют передачу сигналов окружающей среды, тем самым контролируя поступление CO 2 в растения и потерю воды растениями. [ 1 ] [ 2 ] [ 3 ] [ 4 ] Исследования механизмов передачи сигнала замыкающих клеток позволяют понять, как растения могут улучшить свою реакцию на стресс засухи за счет уменьшения потери воды растениями. [ 1 ] [ 41 ] [ 42 ] Замыкающие клетки также представляют собой отличную модель для фундаментальных исследований того, как клетка интегрирует многочисленные виды входных сигналов для получения ответа (открытие или закрытие устьиц). Эти ответы требуют координации многочисленных клеточных биологических процессов в замыкающих клетках, включая прием сигналов, регуляцию ионных каналов и насосов, мембранный транспорт , транскрипцию , цитоскелетные перестройки и многое другое. Задача будущих исследований состоит в том, чтобы приписать функции некоторых из идентифицированных белков этим разнообразным клеточным биологическим процессам. [ нужна ссылка ]
Разработка
[ редактировать ]Во время развития листьев растений специализированные замыкающие клетки дифференцируются из «замыкающих материнских клеток». [ 43 ] [ 44 ] Плотность устьичных пор в листьях регулируется сигналами окружающей среды, в том числе увеличением концентрации CO 2 в атмосфере , что снижает плотность устьичных пор на поверхности листьев у многих видов растений по неизвестным в настоящее время механизмам. Генетику развития устьиц можно непосредственно изучить путем визуализации эпидермиса листа с помощью микроскопа. Идентифицировано несколько основных контролирующих белков, которые функционируют в пути, опосредующем развитие замыкающих клеток и устьичных пор. [ 35 ] [ 44 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Шредер Дж.И., Квак Дж.М. и Аллен Г.Дж. (2001)Передача сигналов абсцизовой кислоты замыкающих клеток и разработка засухоустойчивости растений. Природа 410:327-330.
- ^ Перейти обратно: а б Хетерингтон А.М. и Вудворд Ф.И. (2003)Роль устьиц в восприятии и стимулировании изменений окружающей среды. Природа 424:901-908.
- ^ Перейти обратно: а б с Шимазаки К., Дои М., Ассманн С.М. и Киношита Т. (2007)Световая регуляция движения устьиц. Annu Rev Plant Biol 58:219-247.
- ^ Перейти обратно: а б Квак Дж.М., Мэзер П. и Шредер Дж.И. (2008)Защитная клетка, на которую можно нажать, версия II: Интерактивная модель механизмов и пути передачи сигнала замыкающей клетки. Книга арабидопсиса, ред. Ласт Р., Чанг С., Грэм И., Лейзер О. , МакКлунг Р. и Вейниг С. (Американское общество биологов растений, Роквилл), стр. 1–17.
- ^ Перейти обратно: а б с д Киносита, Тосинори; Эми, Такаши; Томинага, Мисуми; Сакамото, Кодзи; Сигенага, Аяко; Дои, Мичио; Симадзаки, Кен-итиро (1 декабря 2003 г.). «Зависимое от синего света и фосфорилирования связывание белка 14-3-3 с фототропинами в устьичных замыкающих клетках фасоли» . Физиология растений . 133 (4): 1453–1463. дои : 10.1104/стр.103.029629 . ISSN 0032-0889 . ПМК 300702 . ПМИД 14605223 .
- ^ Перейти обратно: а б Киносита, Тосинори; Симадзаки, Кен-итиро (15 ноября 2002 г.). «Биохимические доказательства необходимости связывания белка 14-3-3 для активации H +-АТФазы плазматической мембраны защитных клеток синим светом». Физиология растений и клеток . 43 (11): 1359–1365. дои : 10.1093/pcp/pcf167 . ISSN 1471-9053 . ПМИД 12461136 .
- ^ Цифровая библиотека. «Строение растений и грибов» . regi.tankonyvtar.hu (на венгерском языке) . Проверено 2 апреля 2021 г.
- ^ Имамура С. (1943) Исследования механизма колебаний тургора замыкающих клеток отверстия щели. Ага. Дж Бот 12: 251-346.
- ^ Humble GD & Raschke K (1971)Открытие устьиц количественно связано с транспортом калия. Данные электронного зондового анализа. Физиол растений. 48:447-453.
- ^ Шредер Дж.И., Хедрих Р. и Фернандес Дж.М. (1984)Калия-селективные одиночные каналы в протопластах замыкающих клеток Vicia faba. Природа 312:361-362.
- ^ Ма Ю, Шосткевич И, Корте А, Моес Д, Ян Й, Кристманн А и Гриль Е (2009)Регуляторы активности фосфатазы PP2C действуют как сенсоры абсцизовой кислоты. Наука 324:1064-1068.
- ^ Парк С.Ю., Фунг П., Нишимура Н., Дженсен Д.Р., Фуджи Х., Чжао Ю., Лумба С., Сантьяго Дж., Родригес А., Чоу Т.Ф., Альфред С.Е., Бонетта Д., Финкельштейн Р., Проварт Нью-Джерси, Дево Д., Родригес П.Л., МакКорт П., Чжу Дж.К., Шредер Дж.И., Фолькман Б.Ф. и Катлер С.Р. (2009)Абсцизовая кислота ингибирует протеинфосфатазы типа 2C через семейство белков START PYR/PYL. Наука 324:1068-1071.
- ^ Меймун, Патрис; Видаль, Гийом; Борер, Анн-Софи; Ленер, Арно; Тран, Дэниел; Бриан, Жоэль; Буто, Франсуа; Рона, Жан-Пьер (сентябрь 2009 г.). «Внутриклеточные запасы Са2+ могут участвовать в деполяризации, вызванной абсцизовой кислотой, и закрытии устьиц у Arabidopsis thaliana» . Сигнализация и поведение растений . 4 (9): 830–835. Бибкод : 2009ПлСиБ...4..830М . дои : 10.4161/psb.4.9.9396 . ISSN 1559-2316 . ПМК 2802785 . ПМИД 19847112 .
- ^ Ассманн С.М., Симончини Л. и Шредер Дж.И. (1985) Синий свет активирует электрогенную ионную перекачку в протопластах замыкающих клеток Vicia faba. Природа 318:285-287.
- ^ Шимазаки К., Иино М. и Зейгер Э. (1986)Зависимая от синего света экструзия протонов протопластами замыкающих клеток Vicia faba. Природа 319:324-326.
- ^ Перейти обратно: а б Шредер Дж.И., Рашке К. и Неер Э. (1987) Зависимость K от напряжения + каналы в протопластах замыкающих клеток. Учеб. Натл. акад. наук. США 84:4108-4112.
- ^ Блатт М.Р., Тиль Г. и Трентам Д.Р. (1990)Обратимая инактивация K + каналы замыкающих клеток устьиц Vicia после фотолиза клеточного 1,4,5-трифосфата. Природа 346:766-769.
- ^ Тиль Г., МакРобби EAC и Блатт М.Р. (1992)Мембранный транспорт в замыкающих клетках устьиц: важность контроля напряжения. Дж. Мемб. Биол. 126:1-18.
- ^ Квак Дж.М., Мурата Ю., Байсабал-Агирре В.М., Меррилл Дж., Ван М., Кемпер А., Хоук С.Д., Таллман Г. и Шредер Дж.И. (2001) Доминантная отрицательная замыкающая клетка K + мутанты каналов уменьшают выпрямляющий внутрь K + токи и светоиндуцированное открытие устьиц Arabidopsis. Физиол растений. 127:473-485.
- ^ Лебоди А., Вавассёр А., Хоси Э., Дрейер И., Леонхардт Н., Тибо Дж.Б., Вери А.А., Симонно Т. и Сентенак Х. (2008). Адаптация растений к изменяющейся среде и производство биомассы сильно зависят от калиевых каналов замыкающих клеток. Учеб. Натл. акад. наук. США 105:5271-5276.
- ^ Шредер Дж.И. (1988) К. + транспортные свойства К + каналы в плазматической мембране замыкающих клеток Vicia faba. Дж. Генерал Физиол. 92:667-683.
- ^ Блатт М.Р. и Армстронг Ф. (1993) К. + каналы замыкающих клеток устьиц: вызванный абсцизовой кислотой контроль внешнего выпрямителя, опосредованный pH цитоплазмы. Планта 191:330-341.
- ^ Ache P, Becker D, Ivashikina N, Dietrich P, Roelfsema MR и Hedrich R (2000) GORK, выпрямитель с задержкой наружу, экспрессируемый в замыкающих клетках Arabidopsis thaliana, представляет собой K + -селективный, К + -чувствительный ионный канал. Письмо ФЕВС 486: 93-98.
- ^ Хози Э., Вавассер А., Мулин К., Дрейер И., Гаймар Ф., Пори Ф., Бушерез Дж., Лебоди А., Буше Д., Вери А.А., Симонно Т., Тибо Ж.Б. и Сентенак Х. (2003) The Arabidopsis outward K + канал ГОРК участвует в регуляции движений устьиц и транспирации растений. Proc Natl Acad Sci USA 100:5549-5554.
- ^ Перейти обратно: а б Келлер Б.У., Хедрих Р. и Рашке К. (1989)Зависимые от напряжения анионные каналы в плазматической мембране замыкающих клеток. Природа 341:450-453.
- ^ Перейти обратно: а б с д Шредер Дж. И. Хагивара С. (1989) Цитозольный кальций регулирует ионные каналы в плазматической мембране замыкающих клеток Vicia faba. Природа 338:427-430.
- ^ Хедрих Р., Буш Х. и Рашке К. (1990) Приблизительно 2+ и нуклеотид-зависимая регуляция потенциал-зависимых анионных каналов в плазматической мембране замыкающих клеток. ЭМБО Дж. 9:3889-3892.
- ^ Перейти обратно: а б Шредер Дж.И. и Келлер Б.Ю. (1992)Два типа токов анионных каналов в замыкающих клетках с четкой регулировкой напряжения. Учеб. Натл. акад. наук. США 89:5025-5029.
- ^ Перейти обратно: а б с Пей З.М., Кучицу К., Уорд Дж.М., Шварц М. и Шредер Дж.И. (1997)Дифференциальная регуляция абсцизовой кислотой медленных анионных каналов замыкающих клеток у арабдиопсиса дикого типа и мутантов abi1 и abi2. Растительная клетка 9:409-423.
- ^ Неги Дж., Мацуда О., Нагасава Т., Оба Ю., Такахаши Х., Каваи-Ямада М., Учимия Х., Хасимото М. и Иба К. (2008) Регулятор CO 2 SLAC1 и его гомологи необходимы для анионного гомеостаза в растительных клетках. Природа 452:483-486.
- ^ Триин Вахисалу, Коллист Х., Ван Ю.Ф., Нишимура Н., Чан В.И., Валерио Г., Ламминмаки А., Броше М., Молдау Х., Десикан Р., Шредер Дж.И. и Кангасярви Дж. (2008) SLAC1 необходим для защитных клеток растений S-типа Функция анионных каналов в устьичной передаче сигналов. Природа 452: 487–491.
- ^ Грабов А., Люнг Дж., Жиродат Дж. и Блатт М.Р. (1997)Изменение кинетики анионных каналов в диких типах и трансгенных abi1-1 замыкающих клетках Nicotiana benthamiana под действием абсцизовой кислоты. Плант Дж. 12:203-213.
- ^ Линдер Б. и Рашке К. (1992)Медленный анионный канал в замыкающих клетках, активация при большой гиперполяризации, может быть основным фактором закрытия устьиц. ФЭБС Летт. 131:27-30.
- ^ MacRobbie EAC (1998)Передача сигнала и ионные каналы в замыкающих клетках. Фил. Пер. Рой. Соц. Лондон 1374: 1475–1488.
- ^ Перейти обратно: а б с д Уорд Дж.М. и Шредер Дж.И. (1994) Активируемый кальцием K ++ каналов и кальций-индуцированное высвобождение кальция медленными вакуолярными ионными каналами в вакуолях замыкающих клеток, участвующих в контроле закрытия устьиц. Растительная клетка 6:669-683.
- ^ Аллен Г.Дж. и Сандерс Д. (1996)Контроль ионных токов, защищающих вакуоли клеток, с помощью цитозольного и просветного кальция. Плант Дж. 10:1055-1069.
- ^ Гоберт А., Исаенков С., Фолькер С., Чемпински К. и Маатуис Ф.Дж. (2007) Ген двухпорового канала TPK1 кодирует вакуолярный K + проводимость и играет роль в K + гомеостаз. Proc Natl Acad Sci USA 104:10726-10731.
- ^ Hedrich R & Neher E (1987) Цитоплазматический кальций регулирует зависимые от напряжения ионные каналы в вакуолях растений. Природа 329:833-836.
- ^ Пейтер Э., Маатуис Ф.Дж., Миллс Л.Н., Найт Х., Пеллу Дж., Хетерингтон А.М. и Сандерс Д. (2005) Вакуолярный Ca 2+ -активированный канал TPC1 регулирует прорастание и движение устьиц. Природа 434(7031):404-408.
- ^ Перейти обратно: а б с д и ж Мейер, Стефан; Шольц-Старке, Иоахим; Анджели, Алексис Де; Коверманн, Питер; Бурла, Бо; Гамбале, Франко; Мартинойя, Энрико (2011). «Транспорт малата по вакуольному каналу AtALMT6 в замыкающих клетках подвергается множественной регуляции» (PDF) . Заводской журнал . 67 (2): 247–257. дои : 10.1111/j.1365-313X.2011.04587.x . ISSN 1365-313X . ПМИД 21443686 .
- ^ Пей З.М., Гассемиан М., Квак С.М., МакКорт П. и Шредер Дж.И. (1998)Роль фарнезилтрансферазы в регуляции АБК анионных каналов замыкающих клеток и потере воды растениями. Наука 282:287-290.
- ^ Ван Ю, Ин Дж, Кузьма М, Шалифу М, Образец А, МакАртур С, Учач Т, Сарвас С, Ван Дж, Деннис Д.Т., МакКорт П. и Хуанг Ю (2005). Молекулярная адаптация фарнезилирования для устойчивости растений к засухе и урожайности. защита. Растение J 43:413-424.
- ^ Bergmann DC и Sack FD (2007)Развитие устьиц. Annu Rev Plant Biol 58:163-181.
- ^ Перейти обратно: а б Pillitteri LJ и Torii KU (2007)Нарушая молчание: три белка bHLH управляют решениями о судьбе клеток во время развития устьиц. Биоэссе 29: 861-870.
| Животные | |
|---|---|
| Растения |
|