Литический цикл
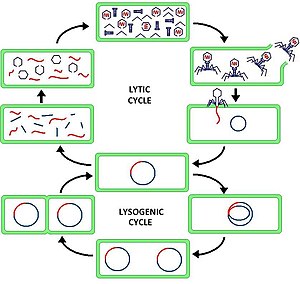
Литический цикл ( / ˈ l ɪ t ɪ k / LIT -ik ) — один из двух циклов вируса размножения (имеется в виду бактериальные вирусы или бактериофаги ), другой — лизогенный цикл . В результате литического цикла происходит разрушение инфицированной клетки и ее мембраны. Бактериофаги, способные проходить только литический цикл, называются вирулентными фагами (в отличие от умеренных фагов).
В литическом цикле вирусная ДНК существует в виде отдельной свободно плавающей молекулы внутри бактериальной клетки и реплицируется отдельно от бактериальной ДНК хозяина, тогда как в лизогенном цикле вирусная ДНК интегрируется в геном хозяина. В этом ключевое различие между литическими и лизогенными циклами. Однако в обоих случаях вирус/фаг реплицируется с использованием механизма ДНК хозяина.
Описание
[ редактировать ]Литический цикл часто разбивают на шесть стадий. Шесть стадий: прикрепление, проникновение, транскрипция, биосинтез, созревание и лизис.
- Прикрепление – фаг прикрепляется к поверхности клетки-хозяина, чтобы ввести в клетку свою ДНК.
- Проникновение – фаг вводит свою ДНК в клетку-хозяина, проникая через клеточную мембрану.
- Транскрипция - ДНК клетки-хозяина разрушается, и метаболизм клетки направлен на инициацию биосинтеза фага.
- Биосинтез – ДНК фага реплицируется внутри клетки, синтезируя новую ДНК фага и белки.
- Созревание – реплицированный материал собирается в полностью сформированные вирусные фаги (каждый из которых состоит из головы, хвоста и хвостовых волокон).
- Лизис – клеточная стенка или мембрана разрываются, разрушая ее и высвобождая вирус.
Прикрепление и проникновение
[ редактировать ]Чтобы заразить клетку-хозяина, вирус должен сначала ввести в клетку собственную нуклеиновую кислоту через плазматическую мембрану и (если она имеется) клеточную стенку. Вирус делает это либо путем прикрепления к рецептору на поверхности клетки, либо с помощью простой механической силы. Связывание происходит за счет электростатических взаимодействий и зависит от pH и присутствия ионов. Затем вирус высвобождает свой генетический материал (одно- или двухцепочечную РНК или ДНК ) в клетку. У некоторых вирусов этот генетический материал имеет кольцевую форму и имитирует бактериальную плазмиду . На этом этапе клетка инфицируется и также может стать мишенью иммунной системы. В основном этому способствуют рецепторы на поверхности клетки. [ нужна ссылка ] Последовательность событий, которые происходят во время инициации инфекции бактериофага , от адсорбции (присоединения) до выброса ДНК из вириона в клетку-хозяина (проникновение), была рассмотрена Молино. [1]
Транскрипция и биосинтез
[ редактировать ]На стадиях транскрипции и биосинтеза вирус захватывает механизмы репликации и трансляции клетки, используя их для создания новых вирусов. Нуклеиновая кислота вируса использует метаболический механизм клетки-хозяина для производства большого количества вирусных компонентов. [2]
В случае ДНК-вирусов ДНК транскрибируется в молекулы информационной РНК (мРНК), которые затем используются для управления рибосомами клетки. Один из первых транслируемых полипептидов разрушает ДНК хозяина. В ретровирусах (которые вводят цепь РНК) уникальный фермент, называемый обратной транскриптазой, транскрибирует вирусную РНК в ДНК, которая затем снова транскрибируется в РНК. Как только вирусная ДНК берет на себя управление, она побуждает механизм клетки-хозяина синтезировать вирусную ДНК, белок и начинает размножаться. [ нужна ссылка ]
Биосинтез (например, Т4 ) регулируется в три фазы продукции мРНК, за которой следует фаза продукции белка. [3]
- Ранняя фаза
- Ферменты модифицируют процесс транскрипции хозяина с помощью РНК-полимеразы . Помимо других модификаций, вирус Т4 изменяет сигма-фактор хозяина, вырабатывая анти-сигма-фактор , так что промоторы хозяина больше не распознаются, но теперь распознают средние белки Т4. Для синтеза белка подпоследовательность Шайна-Дальгарно GAGG доминирует в трансляции ранних генов. [4]
- Средняя фаза
- Нуклеиновая кислота вируса (ДНК или РНК в зависимости от типа вируса). [ нужна ссылка ]
- Поздняя фаза
- Структурные белки, включая белки головы и хвоста. [ нужна ссылка ]
Созревание и лизис
[ редактировать ]Примерно через 25 минут после первичного заражения образуется примерно 200 новых вирионов (вирусных тел). Как только созреет и накопится достаточное количество вирионов, для растворения стенок клеток используются специализированные вирусные белки. Клетка разрывается (то есть подвергается лизису ) из-за высокого внутреннего осмотического давления (давления воды), которое больше не может сдерживаться клеточной стенкой. При этом дочерние вирионы высвобождаются в окружающую среду, где они могут инфицировать другие клетки, и начинается новый литический цикл. Фаг, вызывающий лизис хозяина, называется литическим или вирулентным фагом. [5]
Биохимия регуляции генов
[ редактировать ]В геноме фага есть три класса генов, которые регулируют возникновение литических или лизогенных циклов. Первый класс — это непосредственные ранние гены, второй — отсроченные ранние гены и третий — поздние гены. Нижеследующее относится к хорошо изученному умеренному фагу лямбда E. coli. [ нужна ссылка ]
- Непосредственно ранние гены: эти гены экспрессируются с промоторов, распознаваемых РНК-полимеразой хозяина, и Cro , cII и N. включают CII представляет собой фактор транскрипции, который стимулирует экспрессию основного гена-репрессора лизогенности cI , тогда как Cro является репрессором экспрессии cI . На решение о лизисе-лизогении в основном влияет конкуренция между Cro и CII, в результате которой определяется, достаточно ли создано репрессора CI. Если это так, CI репрессирует ранние промоторы, и инфекция переключается на лизогенный путь. N представляет собой фактор антитерминации, который необходим для транскрипции отсроченных ранних генов.
- Ранние гены с задержкой: к ним относятся гены репликации O и P, а также Q , который кодирует антитерминатор, ответственный за транскрипцию всех поздних генов.
- Поздние гены:
Q-опосредованное включение поздней транскрипции начинается примерно через 6–8 минут после заражения, если выбран литический путь. Более 25 генов экспрессируются с одного позднего промотора, что приводит к четырем параллельным путям биосинтеза. Три пути предназначены для производства трех компонентов вириона: заполненной ДНК головки, хвоста и боковых хвостовых волокон. Вирионы самособираются из этих компонентов, причем первый вирион появляется примерно через 20 минут после заражения. Четвертый путь – лизис. В лямбда-лизисе участвуют 5 белков: холин и антихолин из гена S , эндолизин из гена R и белки спанин из генов Rz и Rz1 . У лямбда дикого типа лизис происходит примерно через 50 минут, в результате чего высвобождается около 100 завершенных вирионов. Время лизиса определяется белками холином и антихолином, причем последний ингибирует первый. Вкратце, белок холин накапливается в цитоплазматической мембране до тех пор, пока внезапно не образует отверстия микронного размера, что запускает лизис. Эндолизин R высвобождается в периплазму, где атакует пептидогликан. Белки спанин Rz и Rz1 накапливаются в цитоплазматической и внешней мембранах соответственно и образуют комплекс, охватывающий периплазму через сеть пептидогликана. Когда эндолизин разрушает пептидогликан, комплексы спанина высвобождаются и вызывают разрушение внешней мембраны. Разрушение пептидогликана эндолизином и разрушение внешней мембраны спаниновым комплексом необходимы для лизиса при лямбда-инфекциях. [ нужна ссылка ]
Ингибирование лизиса: Т4-подобные фаги имеют два гена, rI и rIII , которые ингибируют холин Т4, если инфицированная клетка подвергается суперинфекции другим вирионом Т4 (или близкородственным ему). Повторяющаяся суперинфекция может привести к тому, что инфекция Т4 будет продолжаться без лизиса в течение нескольких часов, что приведет к накоплению вирионов до уровня, в 10 раз превышающего нормальный. [6] [ нужен лучший источник ]
Ссылки
[ редактировать ]- ^ Молино, Ян Дж. (январь 2006 г.). «Пятьдесят три года со времен «Херши и Чейза»; много шума по поводу давления, но какое это давление?». Вирусология . 344 (1): 221–229. дои : 10.1016/j.virol.2005.09.014 .
- ^ «Эбола – литическая или лизогенная» . Эбола-Случаи.com . Проверено 26 января 2023 г.
- ^ Мэдиган, Майкл Т.; Мартинко, Джон М., ред. (2006). Брока Биология микроорганизмов (11-е изд.). Прентис Холл. ISBN 978-0-13-144329-7 .
- ^ Малис, Н. (2012). «Последовательность Шайна-Дальгарно бактериофага Т4: GAGG преобладает в ранних генах». Отчеты по молекулярной биологии . 39 (1): 33–9. дои : 10.1007/s11033-011-0707-4 . ПМИД 21533668 . S2CID 17854788 .
- ^ Гуммалла, Вимати С.; Чжан, Юцзе; Ляо, Йен-Те; Ву, Вивиан Ч. (21 февраля 2023 г.). «Роль умеренных фагов в патогенности бактерий» . Микроорганизмы . 11 (3): 541. doi : 10.3390/microorganisms11030541 . ПМЦ 10052878 .
- ^ «Литический цикл Т-четного бактериофага» . nemetoadreviews.com . Проверено 9 января 2018 г.
