Опыление орхидей
В этой статье есть несколько проблем. Пожалуйста, помогите улучшить его или обсудите эти проблемы на странице обсуждения . ( Узнайте, как и когда удалять эти шаблонные сообщения )
|



Опыление орхидей — сложная глава в биологии этого семейства растений, отличающихся сложностью цветков и сложными экологическими взаимодействиями с агентами- опылителями . Со временем оно привлекло внимание множества ученых, в том числе Чарльза Дарвина , отца теории эволюции путем естественного отбора . Дарвин опубликовал в 1862 году первые наблюдения о фундаментальной роли насекомых в опылении орхидей в своей книге «Оплодотворение орхидей» . [ 1 ] [ примечание 1 ] Дарвин заявил, что разнообразные уловки, которые орхидеи используют для привлечения опылителей, превосходят воображение любого человека. [ 2 ]
Адаптации орхидей к опылению животными
[ редактировать ]97% видов орхидей оплодотворения нуждаются в опылителе для переноса пыльцевых зерен с одного растения на пестики другого растения и, таким образом, для и образования . семян [ 3 ] Пыльца орхидей сгруппирована в компактные массы, называемые поллиниями (единственное число: «поллиниум»), так что сама по себе или под действием ветра пыльца не может распространяться с одного цветка на другой, поэтому опылители необходимы для обеспечения их полового размножения . [ 3 ] Эти опылители очень разнообразны и, в зависимости от рассматриваемого вида, могут быть мухами , комарами , пчелами , осами , бабочками , жесткокрылыми и птицами (особенно колибри ). [ 4 ] [ 5 ] [ 3 ]
Зоофилия , характеризующая орхидеи , предполагает, что животные-опылители регулярно посещают цветы и останавливаются на них достаточно долго; что они с некоторой частотой чистят или касаются пыльников и рыльцев и что пыльца остается прикрепленной к посетителям настолько прочно, что может безопасно достигать рыльцев других цветов. Результат зоофилии существенно зависит от способности животных узнавать цветы с определенного расстояния и от их влечения к цветам одного и того же вида. Таким образом, зоофильные цветы должны обладать «привлекательными продуктами» (приманками, такими как пыльца и нектар ), «средствами привлечения» (такими как ароматы и цвета) и, кроме того, вязкой или прилипшей пыльцой. [ 4 ]
В ходе эволюции покрытосеменных произошла весьма интенсивная дифференциация способов привлечения и притязания, а также формы цветка; благодаря этому все большее число животных может участвовать в опылении. Случайное посещение цветков различными животными в ходе эволюции постепенно привело к установлению тесных взаимоотношений между животными-опылителями и зоофильными цветками с очевидными преимуществами для обеих групп. Для растений это означало повышение точности привлечения только определенных посетителей и перенос пыльцы на рыльца других растений, что приводило к постепенной экономии при производстве пыльцы. растений составляет порядка миллиона Действительно, соотношение между количеством пыльцевых зерен и числом семязачатков, образующихся в цветке, у анемофильных , тогда как у орхидей оно равно единице. Для специализированных животных-опылителей конкуренция с другими антофильными животными уменьшилась, и целевое или специализированное опыление в пределах одного вида стало для них в конечном итоге выгодным. [ 3 ] [ 4 ]
Эволюционное развитие зоофильных покрытосеменных и приспособившихся к ним групп животных можно понимать только как коэволюцию, обусловленную реципрокными отношениями. Приспособление орхидей и их опылителей друг к другу иногда заходит так далеко, что они не могут существовать друг без друга. [ 4 ] Механизмы опыления являются плодом такой совместной эволюции. В целом такие механизмы выгодны обеим сторонам: опылитель получает нектар из цветка орхидеи, а орхидеи, в свою очередь, получают выгоду от переноса пыльцы с одного цветка на другой. Однако во многих случаях привлечение опылителей к орхидеям происходит обманом и не предполагает вознаграждения. [ 3 ]
Цветы орхидеи
[ редактировать ]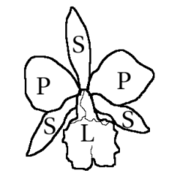


Цветки орхидей обычно гермафродиты , редко однополые и обычно зигоморфные , то есть двусторонне симметричные. У подавляющего большинства родов цветки состоят из трех наружных элементов, называемых чашелистиками , двух боковых и одного спинного, и трех внутренних элементов, называемых лепестками , причем нижний из них видоизменился в губу или губу , больше по размеру и более интенсивно окрашен, чем остальные. . Лабеллум часто трехлопастный или необычной формы, с мясистыми выступами или гребнями или с базальной шпорой и часто имеет совершенно другую окраску, чем два других лепестка. [ 6 ]
Андроцей также обычно имеет одну или две тычинки , иногда три, и прикрепляется к столбику и рыльцу , образуя столбик, называемый гиностемой или гиностегием. Пыльца . сгруппирована в несколько масс различной твердости, называемых поллиниями , которых может быть от одной до двенадцати, но обычно две или четыре Поллинии вместе с липким стеблем, происходящим от пыльника или рыльца, образуют поллиний, транспортную единицу во время опыления . Гинецей . образован тремя сросшимися плодолистиками и бесплоден — развивается ниже чашечки Стиль клеймо и сильно изменены . Стиль одиночный и конечный и является основным компонентом колонны. Рядом с вершиной находится часть рыльца в виде удлиненной лопасти, обычно невосприимчивой, называемая хоботком, расположенная над стигматической областью. Часть хохолка может образовывать липкую платформу, называемую висцидием, прикрепленную к стеблю поллиния. [ 6 ]
Обычно они производят нектар в качестве награды опылителям. Нектарники . различаются по положению и типу Например, они встречаются на губной шпоре, или на вершинах чашелистиков, или на перегородках гинецея. Некоторые виды орхидей являются самоопыляющимися или апомиктиками , т. е. им не нужны опылители для образования семян. [ 7 ]
Привлечение опылителей посредством вознаграждений
[ редактировать ]Многие виды орхидей награждают опылителей пищей, такой как нектар, пищевые волоски или масла, а также другими соединениями, такими как воски, смолы и ароматизаторы. Эти награды, в свою очередь, усиливают поведение опылителей. Однако специализация на одном типе опылителей для обеспечения более эффективного переноса пыльцы обусловила возрастающую морфологическую и структурную специализацию цветков орхидей для обеспечения привлечения одного вида насекомых. [ 3 ] [ 8 ] [ 9 ] [ 10 ]
Этот центральный момент в эволюции межвидовых отношений между орхидеями и их опылителями был уловлен Дарвином при изучении британских и некоторых экзотических видов этого семейства. Например, у мадагаскарского вида Angraecum sesquipedale на цветке имеется шпорец длиной более 30 см, дно которого наполнено нектаром. Дарвин безуспешно пытался удалить поллинии из цветка с помощью иголок. Он смог это сделать только после того, как вставил цилиндр диаметром 2,5 мм в нижнюю часть шпоры и снова вытащил его, так как в этот момент вязкая жидкость осталась прикрепленной к цилиндру. Дарвин полагал, что когда бабочка достигает нижней части шпоры своим хоботом, чтобы выпустить нектар, поллинии останутся прикрепленными к ее голове, как только она вытащит хоботок. Посетив следующий цветок, бабочка опылила его, отложив поллинии на рыльце пестика. [ 11 ] Согласно этому рассуждению, насекомым-опылителем Angraecum sesquipedale должна была быть бабочка со спиритомерой длиной более 30 см — идея, звучавшая для биологов того времени нелепо. Однако предсказание Дарвина могло быть подтверждено в 1903 году, когда на Мадагаскаре была обнаружена бабочка Xanthopan morganii praedicta со спироромбом такого размера. Подвидовой эпитет Predicta относится к тому, что его существование было предсказано Дарвином. [ 3 ]
Бабочку привлекает цветок своим ароматом, особенно ночью. Приближаясь к цветку, он разворачивает хоботок и вставляет его в щель хоботка, ведущую к шпоре. Закончив выделять нектар из основания шпоры, он поднимает голову и отделяет хоботок от шпоры. Когда он выполняет это движение, вязкая жидкость прикрепляется к его голове или другой части тела. Вязкая содержит небольшую ножку (хвостик), на конце которой находятся поллинии. Когда бабочка заканчивает накручивать хоботок и перелетает к другому цветку, хвостовик обезвоживается, а при высыхании его угол по отношению к телу насекомого изменяется, так что, когда насекомое посещает следующий цветок и вставляет в него свой хоботок, поллинии будут обращены друг к другу и впоследствии прикрепятся к рыльцу. После опыления цветы перестают издавать аромат, и вскоре после этого их листочки околоцветника увядают. Однако процесс переноса пыльцы, оплодотворения и образования тысяч новых особей уже обеспечен. [ 3 ]
Многие орхидеи, такие как Angraecum sesquipedale , опыляются ночными бабочками и по этой причине имеют светлые, почти белые цветы и издают аромат в вечернее или ночное время. Примерами этого типа орхидей являются виды Bonatea speciosa , Habenaria epipactidea , род Satyrium , Disa cooperi и D. ophrydea. [ 12 ] С другой стороны, опыление дневными бабочками развилось у нескольких родов орхидей. Цветки этих видов демонстрируют яркие цвета и награждают своих опылителей нектаром. [ 12 ]
Виды, опыляемые пчелами (называемые мелитофильными ), часто источают сильный аромат в течение дня, имеют сильную и яркую окраску. Примерами таких орхидей являются Satyrium erectum и Disa versicolor . Различные виды пчел, опыляющие орхидеи, во многих случаях получают не только нектар, но и масла. Эта награда — довольно редкое явление у растений — известна нескольким десяткам видов орхидей, в том числе около 55 южноафриканским видам, и пчелы используют ее для кормления своих личинок. Роды Disperis , Pterygodium , Corycium , Ceratandra , Evotella , Satyrium и Pachites , среди прочих, включают виды, которые награждают своих опылителей маслами. [ 12 ]
Опыление мухами (миофилия) — второй по распространенности метод среди орхидей, опылители принадлежат к двадцати семействам двукрылых. [ 13 ] Цветы испускают ароматы, напоминающие разлагающиеся органические материалы, экскременты или падаль, субстраты, которые мухи посещают в поисках пищи или для откладывания яиц. [ 8 ] Различные части цветка производят запах гниения или падали и образуют ловушки, удерживающие опылителей, а также обладают различными придатками, привлекающими их, и имеют окраску либо яркую, либо напоминающую плоть, либо какой-то другой оттенок гниющего вещества. [ 14 ] [ 15 ] [ 16 ] [ 17 ] Для цветов дацинифильных видов Bulbophyllum , которые привлекают настоящих плодовых мух, принадлежащих к трибе Dacini (Tephritidae: Diptera), с помощью приятных/пряных аттрактантов (в качестве цветочного синомона) для установления истинного мутуализма, при котором оба организма получают репродуктивные преимущества, между орхидеями и самцы летают - см. ссылку: [ 18 ] Более того, вид B. hortorum эволюционировал вместе с самцами плодовых мух до такой степени, что неповторяющиеся цветки, помимо привлечения мух цветочным синомоном, обладают специальным механизмом опыления, позволяющим выбирать отдельных мух оптимального размера в качестве потенциальных опылителей. [ 19 ]
цветы Stelis hymenantha Например, источают интенсивный аромат сладкого ментола. У основания губки образуется вещество, прочно прилипающее к ее поверхности, наподобие пленки, похожее на нектар. Цветки S. immersa , еще одной орхидеи, опыляемой двукрылыми, также ароматны, но с другим, дынным запахом; и в отличие от первых клейкое вещество расположено как на лепестках, так и у основания губки. Посетителями этих двух видов являются в основном двукрылые нескольких семейств. Большинство мух обычно остаются снаружи цветка, исследуя слизистую жидкость на внешней поверхности лепестков. Опылителями Stelis immersa являются самки небольшой мухи из рода Megaselia ( Phoridae ), единственной мухи точного размера, способной осуществлять перенос пыльцы. Осмотрев нектароподобное вещество, насекомое проникает в цветок сбоку и приземляется на губку, которая находится в положении вниз. Когда это происходит, губка поднимается, прижимая насекомое к вязкому поллинию и захватывая его. Чтобы выйти, муха пятится назад, и вязкая жидкость прилипает к ее грудной клетке. Лабеллум возвращается в исходное положение, освобождая животное. [ 20 ]
Привлечение и вознаграждение с помощью парфюмерии
[ редактировать ]Цветки видов подтриб Stanhopeinae и Catasetinae относятся к самым очаровательным и экстравагантным из всех орхидей. Они не относятся к числу самых красивых, но благодаря своему особому опылению иногда создают уникальные цветы с очень странными механизмами опыления. Виды этих триб опыляются исключительно самцами euglosinae , которые ищут и собирают ароматы в этих цветках; причина такого поведения неизвестна, поскольку духи не обеспечивают питания или защиты, но считается, что они могут быть связаны с брачными ритуалами этих насекомых.
Способы сбора этих ароматов всегда очень похожи. Самец приближается к источнику запаха — осмофору; он обычно садится на губу и начинает длинными густыми волосками передних ног собирать вещества, ответственные за запах. В большинстве случаев они жидкие; у некоторых видов в кристаллической форме. Если они твердые, самец сначала растворяет их выделениями слюнных желез. Когда ароматические компоненты насыщают ножки пчелы, пчела покидает цветок, чтобы с помощью средних ножек передать ароматы в полости задних ножек, где вещества могут сохраняться в течение длительного времени. Разные роды орхидей прикрепляют поллинии к разным частям тела этих опылителей. [ 21 ]
Орхидеи, как и другие растения, избирательно привлекают определенную группу самцов эуглоссина, производя видоспецифичные смеси запахов, которые, по-видимому, действуют как репродуктивной изоляции механизмы . Некоторые орхидеи даже претерпевают морфологические модификации своих цветков таким образом, что они выделяют поллиний только при посещении определенных видов пчел в зависимости от их размера и поведения. По этой причине не все эвглоссины, посещающие тот или иной вид орхидей, являются его эффективными опылителями. [ 22 ]
Некоторые виды Bulbophyllum привлекают исключительно группу самцов плодовых мух Dacini в качестве потенциальных опылителей с помощью специфических аттрактантов самцов, например, метилэвгенола (ME), кетона малины (RK) или зингерона (ZN). Метилэвгенол является предшественником полового феромона для многих видов карантинных вредителей Bactrocera, например, восточной плодовой мушки, B. dorsalis , B. carambolae , B. occipitalis и B. umbrosa . Кетон малины или зигерон включен в качестве компонента полового феромона для самцов, а именно - Zeugodacus caudatus , Z. cucurbitae и Z. tau . [ 23 ] [ 24 ] [ 25 ] [ 26 ] [ 27 ] [ 28 ]
Привлечение опылителей обманом
[ редактировать ]
Многие орхидеи прибегают к тактике соблазнения опылителей, предлагая ароматы, формы, цвета или движения, имитирующие то, что их интересует, не предлагая ничего взамен. Механизмы обмана столь же разнообразны, сколь и удивительны, и перечислены ниже: [ 29 ] [ 30 ]
- Обобщенный обман при кормлении: цветы имитируют форму и окраску видов, которые обычно награждают опылителей.
- подпитка обмана, опосредованного цветочной мимикрией : [ примечание 2 ] В этом случае цветы в точности имитируют тот или иной вид, который награждает опылителей и с которым они соседствуют. [ 31 ]
- мимикрия мест гнездования: цветки имитируют места откладки яиц опылителей.
- имитируя места укрытия: цветы предоставляют опылителям места для укрытия. Эта стратегия может быть не обманчивой, но благоприятной как для насекомого, так и для орхидеи.
- псевдоантагонизм: растение привлекает опылителей за счет активации врожденных защитных механизмов. Таким образом, он имитирует форму, например, другого вида насекомых, которых опылитель хочет прогнать или убить. Опылитель, увидев своего предполагаемого врага, нападает на него снова и снова. В этой тщетной борьбе с цветком насекомое покрывает себя пыльцой, которую оно передаст другим цветам, когда его снова обманут. [ 32 ]
- привлекательность для свиданий : цветы имитируют другие цветы, привлекательные для женщин-опылителей.
- сексуальный обман: в этом случае цветы имитируют брачные сигналы (как зрительные, так и обонятельные) самок-опылителей.
Из описанных механизмов наиболее распространенным среди орхидей является широко распространенный обман при кормлении (сообщается у 38 родов), за которым следует сексуальный обман (18 родов). [ 29 ] [ 30 ]
Привлечение опылителей путем заманивания их в пищу
[ редактировать ]Способность привлекать опылителей, не предлагая им никакого вознаграждения взамен, развилась независимо у нескольких линий покрытосеменных, но обычно только у нескольких видов на семейство. [ 33 ] Напротив, было подсчитано, что около трети видов орхидей используют механизм обмана пищи. [ 34 ] [ 35 ] Этот механизм предполагает привлечение опылителей сигналом о наличии пищи, например нектара или пыльцы, но без какого-либо вознаграждения. Для этого орхидеи используют стратегию сходства с видами, которые вознаграждают своих опылителей и с которыми они сосуществуют. [ 36 ] Чаще всего обман при кормлении заключается в общем сходстве с видами, которые вознаграждают опылителей, т. е. этот тип орхидей имеет большие, ярко окрашенные цветы - стратегия, которая использует врожденные предпочтения опылителей к этому классу цветов. [ 30 ] [ 35 ] [ 37 ]
Имитация других растений
[ редактировать ]В этом случае орхидея «обманывает» опылителей, типичных для других видов растений, заставляя их опылять. Метод их обмана — имитировать цветы других видов. Очень показательный случай этого наблюдается у орхидеи Epidendrum ibaguense . Эта наземная или литофильная орхидея распространена от Мексики до Боливии и Бразилии . Его оранжевые цветы с ярко-желтой губкой имитируют цветы асклепидовых , Asclepias curassavica . Существует бабочка Agraulis vanillae , которая обычно посещает этот вид, чтобы добыть нектар в обмен на транспортировку пыльцы. Однако во многих случаях бабочка, привлеченная цветом и формой цветков эпидендрума , приближается к ним и вставляет ротовой аппарат ( хоботок ) в узкий проток ( гиностемий ), который из-за своего очень маленького диаметра вызывает появление спитромера. оказаться в ловушке на несколько мгновений. Борьба насекомого за освобождение приводит к тому, что поллинии орхидеи прилипают к его головке. После того, как его выпустили и снова обманули другие Эпидендрум будет транспортировать пыльцу и, таким образом, позволит опылять этот вид, но не получив за свои услуги нектара. [ 38 ]
Привлечение опылителей путем сексуального приманивания
[ редактировать ]Есть обманчивые цветки, имитирующие форму, волосатость и запах самок некоторых ос или пчел. Самый известный случай - орхидея Ophrys насекомоядная из южной Европы; его посещают только два вида ос рода Argogorytes . Самцы рождаются весной на несколько недель раньше самок, и при первых полетах их привлекает аромат цветков Офриса , похожий на феромоны, выделяемые самками. Кроме того, губка по форме, цвету и текстуре похожа на самку. Этот процесс называется псевдокопуляцией , потому что самцы ос пытаются спариваться с цветком и при этом вступают в контакт с пыльником, последовательно перенося поллинии с одного цветка на другой. [ 39 ]
Опыление путем псевдокопуляции впервые описано в 1916 и 1917 А. Пуянном и Х. Корревоном , когда они изучали взаимоотношения орхидеи Ophrys speculum и сколиидной осы Campsoscolia ciliata в Алжире . [ 40 ] [ 41 ] Публикации этих французских биологов оставались незамеченными до тех пор, пока Роберт Годфри в 1925 г. не подтвердил их наблюдения, что и определило рост интереса к этой теме. [ примечание 3 ] За статьей 1925 года последовало большое количество публикаций австралийского биолога Эдит Коулман об опылении орхидей рода Cryptostylis самцами осы ихневмоноидеа - Lissopimpla excelsa . [ 42 ] [ 43 ] [ 44 ]
По этому механизму размножаются несколько родов наземных орхидей. Наиболее известными и наиболее документированными родами являются, помимо уже упомянутых Ophrys и Cryptostylis , Drakaea , Caladenia , [ 45 ] Чилоглоттис , [ 46 ] Геобласта , [ 47 ] Артрохил , Калохил , [ 48 ] Лепорелла [ 49 ] и Спикулея . Подавляющее большинство родов наземных орхидей, использующих псевдокопуляцию, встречается в Австралии , а самый крупный род орхидей с псевдокопулятивными видами - Ophrys из Европы. Этот механизм ни в коем случае не ограничивается конкретным континентом, поскольку он также известен для южноамериканского вида ( Geoblasta penicillata ). [ 50 ] и две южноафриканские орхидеи рода Disa . [ 51 ]
Идентичный механизм использует Tolumnia henekenii , цветок которой напоминает самку пчелы Centris insularis , и действует настолько хорошо, что самец этого вида обманывается и пытается совокупиться с имитацией самки, предлагаемой цветком. При этом он опыляет цветок.
У Калеаны , называемой в англоязычных странах утиной орхидеей из-за сходства ее губки с головой утки, а остальной части цветка с телом утки в полете, описан несколько иной механизм псевдокопуляции. Этот вид использует пружинный механизм, чтобы поймать насекомых в сумку, и их единственная возможность выбраться - через поллиний и рыльце пестика. Мужские насекомые-опылители приземляются на губу, что запускает двухшарнирный механизм релаксации (один - губка-ламелла, другой - пластинка-околоцветник), который переворачивает насекомое задней частью грудной клетки в сумку, содержащую рыльце и поллинии. . [ 52 ] [ 53 ]
Псевдокопуляция свойственна не только опылителям отряда перепончатокрылых (пчёлы и осы). ) также было описано Фактически, для двукрылых (комаров рода Bradysia псевдокопулирование видов Lepanthes , одного из крупнейших родов покрытосеменных растений, населяющих неотропические тропические леса. [ 54 ]
Цветки орхидей рода Ophrys не только имитируют форму, размеры и окраску самок своих опылителей, но и источают аромат, включающий в себя несколько соединений, содержащихся в половых феромонах самок, тем самым усиливая половое поведение самцов. . Летучие соединения, выделяемые Ophrys iricolor , и женские феромоны его вида-опылителя Andrena morio были сравнены как химически, так и электрофизиологически. Открыто более 40 соединений, в том числе алканы и алкены с числом атомов углерода от 20 до 29, альдегиды с числом атомов углерода от 9 до 24 и два сложных эфира . Почти все эти соединения были обнаружены в одинаковых пропорциях как в цветочных экстрактах O. iricolor, так и в экстрактах кутикулярной поверхности самок A. morio . [ 55 ]
Биологически активные летучие соединения этого подвоя очень похожи на те, которые используются другими видами Ophrys , которые псевдокопулируются самцами родов Andrena и Colletes . [ 56 ] [ 57 ] [ 58 ] [ 59 ] [ 60 ]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Книга была отредактирована под названием « О различных приспособлениях, с помощью которых британские и зарубежные орхидеи оплодотворяются насекомыми, и о положительных эффектах скрещивания» . Эта книга была первой подробной демонстрацией силы естественного отбора, которая могла бы объяснить, как возникли сложные взаимоотношения между орхидеями и их опылителями. Дарвин описал подробные наблюдения за механизмом опыления различных орхидей, свои эксперименты и препарирование цветов, а также объяснил ряд ранее игнорированных аспектов биологии опыления этого семейства, таких как Catasetum загадка , которая, как считалось, состояла из цветков трех разные виды на одном и том же растении, а также сделал предсказания, которые позже подтвердились в природе.
- ^ Существует два основных типа мимикрии, имеющих отношение к экологии опыления: мюллеровская и бейтсовская мимикрия . В первом случае (который многие авторы воспринимают как конвергенцию, а не мимикрию) несколько видов с одинаковой численностью в природе к взаимной выгоде развивают один и тот же стиль пропаганды , например, в качестве предупреждения хищникам. Напротив, мало представленный в природе организм, который имитирует более многочисленный организм, чтобы обмануть третьего, развивает однонаправленное преимущество, которое называется бейтсовской мимикрией. (Проктор и Йео, стр. 375)
- ↑ Предмет, получивший название «Пуянская мимикрия» в честь своего первооткрывателя.
Ссылки
[ редактировать ]- ^ «Дарвин Онлайн: Оплодотворение орхидей» .
- ^ Дарвин (1862 , стр. 366 )
- ^ Перейти обратно: а б с д и ж г час ван дер Сингель, Н. 2001. Атлас опыления орхидей: Америка, Африка, Азия и Австралия . ЦРК Пресс. ISBN 9789054104865
- ^ Перейти обратно: а б с д Страсбургер, Э. (1994). Трактат по ботанике . 8-е издание. (На испанском языке). Омега, Барселона, 1088 стр.
- ^ Гола, Г., Негри, Г. и Каппелетти, К. 1965. Tratado de Botánica . 2-е издание. (На испанском языке). Редакция Labor SA, Барселона, 1110 стр.
- ^ Перейти обратно: а б Крибб, Филипп (2001). «Морфология орхидных». В: Под ред. А.М. Приджена, П.Дж. Крибба, М.В. Чейза и Ф.Н. Расмуссена, Genera Orchidacearum , vol. 1. Издательство Оксфордского университета , Оксфорд, Великобритания. ISBN 0-19-850513-2 .
- ^ Нейланд, MRM; Уилкок, CC (1998). «Набор фруктов, награда за нектар и редкость среди орхидных». Американский журнал ботаники . 85 (12): 1657–1671. дои : 10.2307/2446499 . JSTOR 2446499 . ПМИД 21680327 . (требуется подписка)
- ^ Перейти обратно: а б Пийл, Л. ван дер и Ч. Додсон. (1969). Цветы орхидеи. Их опыление и эволюция . Университет Майами Пресс. Корал Гейблс. стр. 101-122.
- ^ Дресслер, Роберт Л. (1993). Филогения и классификация семейства орхидных. Издательство Кембриджского университета ISBN 0-521-45058-6 , 9780521450584
- ^ Проктор, Майкл; Йо, Питер и Лэк, Эндрю. (1996). Естественная история опыления. Портленд, Орегон. Лесной пресс. 479 с.
- ^ Дарвин (1862 , стр. 197-203 )
- ^ Перейти обратно: а б с Курцвейл, Х. (2000). Южноафриканские орхидеи: размножение и опыление. Национальный институт биоразнообразия, Южная Африка.
- ^ Кристенсен, Делавэр (1994). «Опыление мухами орхидных». В: Ардитти, Дж. (ред.). Биология орхидей: обзоры и перспективы . VI. Джон Уайли и сыновья. Итака. Нью-Йорк. стр. 415-454.
- ^ Чейз, MW (1985). Опыление Pleurothallis endosthachys . Являюсь. Орх. Соц. Бык . 54:431-434
- ^ Джонсон, С.Д. и Ти.Дж. Эдвардс. (2000). Строение и функции поллинарии орхидейной. Пл. Сист. Эвол. 222: 243-269.
- ^ Борба, Э.Л., Дж.М. Феликс, В.Н. Сольферини и Дж. Семир. (2001б). «Опыляемые мухами виды Pleurothallis (Orchidaceae) обладают высокой фенетической изменчивостью: данные изоферментных маркеров». Являюсь. Дж. Бот. 88: 419-428.
- ^ Борба, Э.Л. и Дж. Семир. (2001). (Orchidaceae) , опыляемых мухами «Специфичность опылителей и конвергенция видов Pleurothallis : подход с использованием нескольких популяций». Энн. Бот. 88: 75-88.
- ^ https://en.wikipedia.org/wiki/Attractant, связанный с синомоном; и ссылки там
- ^ Тан, К.Х., PT Онг и LT Тан. (2023). «Морфология и движение цветков Bulbophyllum hortorum (Orchidaceae) позволяют выбирать самцов плодовых мух Dacini оптимального размера. Arthropod-Plant Interact. 17: 647–660.
- ^ Альборес-Ортис, Октавио и Виктория Соса. (2006). Опыление двух симпатрических видов Stelis (PLEUROTALLIDINAE, ORCHIDACEAE) . Acta Botanica Mexicana 74: 155–168.
- ^ ГЮНТЕР ГЕРЛАХ. (2003). ЛАНКЕСТЕРИАНА 7: 104-106.
- ^ Рамирес, Сантьяго; Роберт Л. Дресслер и Моника Оспина. (2002). Эуглоссиновые пчелы (Hymenoptera: Apidae) Неотропического региона: Список видов с примечаниями по их биологии . (на испанском языке) Biota Colombiana 3:7-118.
- ^ Тан, К.Х. и Нишида, Р. (2000) Взаимные репродуктивные преимущества дикой орхидеи, Bulbophyllum patens и плодовых мух Bactrocera через цветочный синомон. Журнал химической экологии, 26: 533-546.
- ^ Тан, К.Х., Нисида Р. и Тунг, Ю.К. (2002) Цветочный синомон дикой орхидеи Bulbophyllum cheiri привлекает плодовых мух Bactrocera для опыления. Журнал химической экологии, 28:1161-1172.
- ^ Тан, К. Х. и Нисида, Р. (2005) Синомон или Кайромон? - Цветок Bulbophyllum apertum (Orchidaceae) выделяет малиновый кетон, привлекающий плодовых мух Bactrocera. Журнал химической экологии, 31(3): 497-507.
- ^ Тан, К.Х., Тан, Л.Т. и Нисида, Р. (2006) Цветочный фенилпропаноидный коктейль и архитектура орхидеи Bulbophyllum vinaceum в привлечении плодовых мух для опыления. Журнал химической экологии, 32: 2429-2441.
- ^ Тан, К. Х. и Нишида, Р. (2007) Зингерон в цветочном синомоне Bulbophyllum baileyi (Orchidaceae) привлекает плодовых мух Bactrocera во время опыления. Биохимическая систематика и экология, 31: 334-341.
- ^ Катте, Т., Тан, К.Х., Су, Ж., Оно, Х. и Нишида, Р. (2020) Фенилбутаноиды в двух близкородственных орхидеях плодовой мухи, Bulbophyllum hortorum и B. macranthoides подвида tollenoniferum: множество привлекательных компонентов для виды тефритидной плодовой мухи, чувствительные к кетону малины. Прикладная энтомология и зоология, 55 (1): 55-64.
- ^ Перейти обратно: а б Ерсакова, Яна; Стивен Д. Джонсон и Павел Киндлманн. (2006). Механизмы и эволюция обманного опыления у орхидей . Биологические обзоры , 81:2:219-235.
- ^ Перейти обратно: а б с Уотерман, Ричард Дж. и Мартин И. Бидартондо. (2008). Обман вверху, обман внизу: связь опыления и микоризной биологии орхидей . Дж. Эксп. Бот. 59: 1085-1096.
- ^ Дафни, А. (1984). Мимикрия и обман при опылении. Ежегодный обзор экологии и систематики. Том. 15:259-278
- ^ Орлеан, С. «Умное растение». НОВА .
- ^ Реннер, СС (2006). «Бесполезные цветы покрытосеменных растений и роль когнитивных способностей насекомых в их эволюции». В: Васер, Нью-Мексико, Оллертон. Дж., ред. Взаимодействия растений и опылителей: от специализации к генерализации. Чикаго, Иллинойс, США: Издательство Чикагского университета . 123-144.
- ^ Акерман, JD (1986). «Механизмы и эволюция систем опыления орхидей, обманывающих пищу». Линдлеяна. 1:108-113
- ^ Перейти обратно: а б Нильссон, Луизиана (1992) «Биология опыления орхидей». Тенденции экологии и эволюции 7:255-259.
- ^ Джонсон, С.Д. (2000). «Бейтсианская мимикрия у бесполезной орхидеи Disa pulchra и ее последствия для поведения опылителей». Биологический журнал Линнеевского общества. 71:119-132.
- ^ Шистль, Ф.П. (2005). «Об успехе аферы: опыление обманом у орхидей». Naturwissenschaften 92:255-264.
- ^ Эдвард Таборда, Александр. «Естественное опыление орхидей» . Клуб перуанских орхидей (на испанском языке). Архивировано из оригинала 18 февраля.
- ^ Вольф, Т. (1950). «Опыление и оплодотворение мухи ophrys, Ophrys Insectifera L. в Аллинделилле Фредсков, Дания». Ойкос 2:20-59.
- ^ Корревон, Х. и Пуянн, А. (1916). «Загадочный случай миметизма в Офридах». (на французском языке). Дж. Сок. Нат. Садоводство. Франция. 17, 29, 41, 84.
- ^ Пуянн, А. (1917). «Оплодотворение Офриса насекомыми». (на французском языке). Bull Soc Hist Nat Afr Nord 1917;8:6-7
- ^ Коулман, Э. (1927). «Опыление орхидеи Cryptostylis leptochila ». Викторианский натуралист 44: 20-22.
- ^ Коулман, Э. (1929). «Опыление Cryptostylis subulata (Labill.) Reichb». Викторианский натуралист 46: 62-66.
- ^ Коулман, Э. (1930). «Опыление Cryptostylis erecta R.Br». Викторианский натуралист 46: 236–238.
- ^ Пикалл, Р. и Битти, AJ (1996). «Экологические и генетические последствия опыления орхидеи Caladenia tentaculata путем сексуального обмана». Эволюция 50, 2207-2220 гг.
- ^ Мант Дж., Пикалл Р. и Уэстон П.Х. (2005) «Особая привлекательность опылителей и диверсификация вводящих в заблуждение с сексуальной точки зрения Chiloglottis (Orchidaceae)». Систематика и эволюция растений , 253, 185–200.
- ^ Корреа, М.Н. (1968). Реабилитация рода Geoblasta Barb Rodr». Rev. Museo La Plata, № 11, 69–74.
- ^ Фордхэм, Ф. (1946). «Опыление Calochilus Campestris ». Викт. Натуралист . 62, 199–201.
- ^ Пикалл, Р. (1989a). «Уникальное опыление Leporella fimbriata (Orchidaceae) псевдокопулирующими крылатыми муравьями-самцами Myrmecia urens (Formicidae)». Систематика и эволюция растений 167, 137–148.
- ^ Чиотек, Л., П. Гиоргис, С. Бенитес-Виейра и А. А. Кокуччи. (2006). «Первый подтвержденный случай псевдокопуляции у наземных орхидей Южной Америки: опыление Geoblasta pennicillata (Orchidaceae) Campsomeris bistrimacula (Hymenoptera, Scoliidae)». Флора 201:365-69.
- ^ Уайтхед, В.Б. и Штайнер, К.Э. (1991). «Самцы Podalonia canescens (Sphecidae), опыляющие орхидею Disa atricapilla ». Материалы 8-го конгресса Энтомологического общества Южной Африки, Блумфонтейн, 1–4 июля .
- ^ Кэди, Л. (1965). «Заметки об опылении Caleana major ». Орчадский. 2:34.
- ^ Хоппер, Стивен Д.; Эндрю П. Браун. (2006) «Австралийские орхидеи летучей утки, опыляемые осами, пересмотренные (Paracaleana: Orchidaceae)». Австралийская систематическая ботаника 19:3, 211.
- ^ БЕЛЫЙ, Марио А. и Габриэль БАРБОЗА. (2005). «Псевдокопулятивное опыление Lepanthes (Orchidaceae: Pleurothallidinae) грибными комарами». Энн Бот 95: 763–772.
- ^ Стёкль, Йоханнес; Роберт Твеле, Дирк Х. Эрдманн, Виттко Франке, Манфред Аяссе. (2007). «Сравнение цветочного аромата сексуально обманчивой орхидеи Ophrys iricolor и феромона женского пола ее опылителя Andrena morio ». Химиоэкология, том 17 (4): 231-233.
- ^ Мант, Дж. Г., Брендли, К., Верикекен, Нью-Джерси, Шульц, К., Франке, В. и Шистль, Ф. П. (2005). «Кутикулярные углеводороды как источник полового феромона у Colletes cuncularius (Hymenoptera: Colletidae) и ключ к его мимикрии сексуально обманчивой орхидеи Ophrys exaltata ».
- ^ Шистль, Ф.П., М. Аясс, Х.Ф. Паулюс, К. Лёфстедт, Б.С. Ханссон, Ф. Ибарра, В. Франке. (2000). «Имикрия половых феромонов у ранней орхидеи-паука ( Ophrys sphegodes ): структура углеводородов как ключевой механизм опыления путем сексуального обмана». Журнал сравнительной физиологии A: Нейроэтология, сенсорная, нервная и поведенческая физиология 186:567-574.
- ^ Аяссе, М., Ф. П. Шистль, Х. Ф. Паулюс, Д. Эрдманн и В. Франке. (1997). «Химическая связь в репродуктивной биологии Ophrys sphegodes ». Сообщения Немецкого общества общей и прикладной энтомологии 11: 473-476.1
- ^ Аясс, М., Ф. П. Шистль, Х. Ф. Паулюс, Ф. Ибарра и В. Франке. (2003). «Привлечение опылителей к сексуально обманчивой орхидее с помощью нетрадиционных химикатов». Труды Королевского общества B: Биологические науки 270: 517-522.
- ^ Шистл Ф.П., Пикалл Р. и Мант Дж. (2004) «Химическая связь в сексуально обманчивом роде орхидей Cryptostylis ». Ботанический журнал Линнеевского общества 144, 199–205.