Страменопил
| Страменопилес Временной диапазон: поздний мезопротерозой – настоящее время,
| |
|---|---|
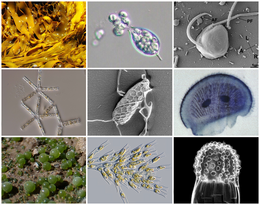 | |
| Разнообразие страменопилей | |
| Научная классификация | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Клэйд : | САР |
| Клэйд : | Страменопилес Паттерсон 1989 г. [ 2 ] купить Аддл и др. 2005 г. [ 3 ] |
| Типы и подтипы [ 4 ] | |
| Разнообразие | |
| >100000 видов [ 1 ] | |
| Синонимы | |
| |
Stramenopiles Heterokonts , также называемые организмов , , представляют собой кладу отличающихся наличием жестких трехсторонних внешних волосков. У большинства видов волоски прикреплены к жгутикам , у некоторых они прикреплены к другим участкам клеточной поверхности, а у некоторых они были вторично утеряны (в этом случае родство с предками страменопиля очевидно из других общих цитологических особенностей или генетических особенностей). сходство). Stramenopiles представляют собой одну из трех основных клад супергруппы SAR , наряду с Alveolata и Rhizaria .
Страменопилы — эукариоты ; большинство из них одноклеточные, но некоторые из них многоклеточные, включая некоторые крупные морские водоросли, бурые водоросли . В группу входят разнообразные водорослевые протисты , гетеротрофные жгутиконосцы, опалины и близкородственные жгутиконосцы протеромонады (все эндобионты у других организмов); актинофриидные солнечники и оомицеты . Трехраздельные волоски, характерные для группы, утеряны у некоторых включенных таксонов, например у большинства диатомовых водорослей .
Многие страменопилы представляют собой одноклеточные жгутиконосцы , а большинство других в какой-то момент своего жизненного цикла производят жгутиковые клетки, например, в виде гамет или зооспор . Большинство жгутиковых гетероконтов имеют два жгутика; передний жгутик имеет один или два ряда жестких волосков или мастигонем , а задний жгутик без таких украшений, гладкий, обычно короче или в некоторых случаях не выступает из клетки.
Термин «гетероконт» используется как прилагательное, обозначающее, что клетка имеет два разных жгутика, и как название таксона. Однако группы, включенные в этот таксон, сильно различались, создавая «проблему гетероконта», которая теперь решена определением страменопилей.
История
[ редактировать ]Термин «страменопил» был введен DJ Patterson в 1989 году, определяя группу, которая пересекается с неоднозначно определяемыми гетероконтами . [ 12 ] [ 13 ] Название «страменопил» обсуждалось Дж. К. Дэвидом. [ 14 ]
Проблема гетероконта
[ редактировать ]Термин «гетероконт» используется как прилагательное, указывающее на то, что клетка имеет два разных жгутика, и как название таксона. Таксон Heterokontae был введен Александром Лютером в 1899 году для водорослей, которые сейчас считаются Xanthophyceae . [ 15 ] Но тот же термин использовался и для других групп водорослей. Например, в 1956 году Коупленд [ 16 ] использовал его для включения ксантофитов (используя название Vaucheriacea), группы, которая включала так называемые хризофиты , силикофлагелляты и гипохитриды . Коупленд также включил неродственные воротниковые жгутиконосцы (как хоанофлагелляты ), в которые он поместил бикозоэциды . Он также включил неблизкородственные гаптофиты . Последствием объединения нескольких понятий с таксоном «гетероконт» является то, что значение слова «гетероконт» можно прояснить, только сославшись на его использование: Heterokontae sensu Luther 1899; Heterokontae sensu Copeland 1956 и т. д. Такое контекстное уточнение встречается редко, поэтому, когда используется название таксона, неясно, как его следует понимать. Термин «Гетероконт» потерял свою полезность в критических дискуссиях об идентичности, природе, характере и родстве группы. [ 17 ] Термин «страменопил» был направлен на идентификацию клады (монофилетической и голофилетической линии), используя подход, разработанный трансформированными кладистами и указывающий на определяющую инновационную характеристику или апоморфию. [ 18 ]
Со временем сфера применения изменилась, особенно когда в 1970-х годах ультраструктурные исследования выявили большее разнообразие водорослей с хромопластами (хлорофиллами а и с), чем считалось ранее. В то же время протистологическая перспектива пришла на смену концепции XIX века, основанной на разделении одноклеточных эукариот на животных и растения. Одним из последствий было то, что множество гетеротрофных организмов, многие из которых ранее не считались «гетероконтами», стали рассматриваться как связанные с «основными гетероконтами» (теми, которые имеют передние жгутики с жесткими волосками). В число недавно признанных родственников вошли паразитические опалины , протеромонады и актинофриидные солнечники . Они присоединились к другим гетеротрофным протистам, таким как бикозоэциды , лабиринтулиды и оомицеты , которые одни включались в гетероконты и исключались другими. Вместо того, чтобы продолжать использовать имя, значение которого со временем изменилось и, следовательно, было неоднозначным, название «страменопил» было введено для обозначения клады протистов, имевших трехсторонние жесткие (обычно жгутиковые) волосы, и всех их потомков. Молекулярные исследования подтверждают, что гены, кодирующие белки этих волос, присущи исключительно страменопилам. [ 19 ]
Характеристики
[ редактировать ]

Хорошо охарактеризована предполагаемая апоморфия трехраздельных жгутиковых волосков страменопилей. Базальная часть волос гибкая и прикрепляется к клеточной мембране; во второй части преобладает длинная жесткая трубка («соломинка» или «страмен»); и, наконец, трубка покрыта множеством тонких волосков, называемых мастигонемами . [ 20 ] Белки, кодирующие мастигонемы, по-видимому, являются эксклюзивными для клады страменопилей и присутствуют даже у таксонов (таких как диатомовые водоросли), у которых больше нет таких волосков. [ 21 ]
У большинства страменопилей около вершины есть два жгутика. [ 22 ] Обычно они поддерживаются четырьмя корнями микротрубочек , имеющими характерный рисунок. Внутри жгутика имеется переходная спираль, где бьющаяся аксонема с характерным геометрическим рисунком из девяти периферических куплетов вокруг двух центральных микротрубочек меняется на девятитройную структуру базального тельца. [ 23 ]
Пластиды
[ редактировать ]Многие страменопилы имеют пластиды , которые позволяют им фотосинтезировать , используя свет для производства собственной пищи . Эти пластиды имеют зеленоватый, оранжевый, золотистый или коричневый цвет из-за присутствия хлорофилла а , хлорофилла с и фукоксантина . Такая форма пластиды называется страменохромом или хромопластом . [ а ] Наиболее значимыми автотрофными страменопилями являются бурые водоросли (раковины и многие другие водоросли) и диатомовые водоросли . Последние являются одними из наиболее важных первичных производителей в морских и пресноводных экосистемах. [ 24 ] Большинство молекулярных анализов показывают, что большинство базальных страменопилей лишены пластид и, соответственно, являются бесцветными гетеротрофами , питающимися другими организмами. Это означает, что страменопилы возникли как гетеротрофы, диверсифицировались, а затем некоторые из них обзавелись хромопластами. Некоторые линии (например, линия аксодина , в которую входили хромофитные пединеллиды , бесцветные цилиофрииды и бесцветные актинофриидные гелиозоа) вторично вернулись к гетеротрофии. [ 25 ] [ 26 ]
Экология
[ редактировать ]
Некоторые страменопилы важны как автотрофы и гетеротрофы в природных экосистемах; другие являются паразитическими. Blastocystis — желудочно-кишечный паразит человека; [ 27 ] опалины и протеромонады обитают в кишечнике хладнокровных позвоночных и описаны как паразитические; [ 28 ] Оомицеты включают некоторые важные патогены растений, такие как возбудитель фитофтороза картофеля Phytophthora infestans . [ 29 ] Диатомеи вносят основной вклад в глобальные углеродные циклы, поскольку они являются наиболее важными автотрофами в большинстве морских сред обитания. [ 30 ] Бурые водоросли, в том числе знакомые морские водоросли, такие как ракушки и бурые водоросли, являются основными автотрофами приливных и сублиторальных морских местообитаний. [ 31 ] Некоторые из бактериоядных страменопилов, такие как Cafeteria , являются обычными и широко распространенными потребителями бактерий и, таким образом, играют важную роль в переработке углерода и питательных веществ в микробных пищевых сетях . [ 32 ] [ 33 ]
Эволюция
[ редактировать ]Внешний
[ редактировать ]Страменопилы наиболее тесно связаны с альвеолятами и ризариями, все из которых имеют трубчатые митохондриальные кристы и вместе образуют супергруппу SAR , название которой образовано из их инициалов. [ 34 ] [ 26 ] [ 35 ] Предок супергруппы SAR, по-видимому, захватил одноклеточную фотосинтезирующую красную водоросль , и у многих Stramenopiles, а также у членов других групп SAR, таких как Rhizaria, все еще есть пластиды, которые сохраняют двойную мембрану красной водоросли и двойную мембрану, окружающую это всего четыре мембраны. [ 36 ] Кроме того, виды Telonemia , родственной группы SAR, демонстрируют гетероконтные жгутики с трехсторонними мастигонемами, что указывает на более древнее происхождение характеристик страменопиля. [ 37 ]

Внутренний
[ редактировать ]Следующая кладограмма суммирует эволюционные отношения между Stramenopiles. Филогенетические монофилетический отношения Bigyra сильно различаются от одного анализа к другому: он был обнаружен либо как монофилетический, либо как . [ 38 ] [ 39 ] или парафилетический . В парафилетическом состоянии порядок ветвления групп бигирана также варьируется: в некоторых исследованиях Sagenista является наиболее базальноветвящейся кладой, [ 38 ] [ 40 ] [ 41 ] в то время как в других Opalozoa является наиболее базальным. [ 42 ] Тем не менее, Platysulcea последовательно считается сестринской кладой всех других страменопилей. [ 39 ] [ 40 ] Кроме того, вид жгутиконосцев, открытый в 2023 году, Kaonashia insperata , остается в неопределенном филогенетическом положении, но более тесно связан с Gyrista, чем с другими кладами. [ 41 ]
| Страменопилес | ||

Классификация
[ редактировать ]
Классификация Stramenopiles по Adl et al. (2019), с дополнениями из новых исследований: [ 43 ] [ 4 ]
- Платисульча Кавальер-Смит 2017 [ 39 ]
- Бигира Кавальер-Смит, 1998 г., исправлено. 2006 г.
- Opalozoa Cavalier Smith 1991, исправлено. 2006 г.
- Наномонадея Кавальер-Смит 2012 г.
- Опалината Веньон 1926 г., исправлено. Кавалер-Смит, 1997 г. [= Слопалинида Паттерсон, 1985 г. ]
- Bicosoecida Grasse 1926, исправлено. Карпов 1998 г.
- Сагениста Кавальер-Смит 1995 г.
- Лабиринтуломицеты Дик 2001
- Pseudophyllomitidae Shiratori et al. 2016 год [ 40 ]
- Opalozoa Cavalier Smith 1991, исправлено. 2006 г.
- Гириста Кавальер-Смит, 1998 г.
- Бигиромонада Кавальер-Смит 1998 г.
- Developea Карпов и Алеошин 2016
- Пирсониалес Кавальер-Смит, 1998 г., исправлено. 2006 г.
- Псевдогрибы Кавальер-Смит 1986 г.
- Гифохитриалес Воробей 1960
- Peronosporomycetes Dick 2001 [=Oomycetes Winter 1897, исправлено. Дик 1976 ]
- Actinophryidae Claus 1874, исправлено. Хартманн 1926 г.
- Ochrophyta Cavalier-Smith 1986, исправлено. Кавалер-Смит и Чао, 1996 г.
- Крисиста Кавальер-Смит, 1986 г.
- Chrysoparadoxophyceae Wetherbee 2019 [ 44 ]
- Хризофицеи Пашер 1914 г.
- Chloromophyceae ( сомнительное название ) [ 45 ]
- Eustigmaphyceae Hibberd & Leedale, 1971 г.
- Olisthodiscophyceae Barcytė, Эйкрем и М. Элиаш, 2021 г. [ 46 ] [ 47 ]
- Phaeophyceae Хансгирг 1886 г.
- Phaeosacciophyceae Р.А.Андерсен, Л.Граф и HSYoon 2020 [ 48 ]
- Phaeothamniophyceae Andersen & Bailey в Bailey et al. 1998 год
- Raphidophyceae Chadefaud 1950, исправить. Лес 1980
- Schizocladiophyceae Генри, Окуда и Каваи, 2003 г.
- Synchromophyceae S.Horn & C.Wilhelm 2007 [=Picophagea Cavalier-Smith 2006, исправить. 2017 ]
- Xanthophyceae Allorge 1930, исправлено. Фрич 1935 [Heterokontae Luther 1899 ; Гетеромонадея Лидейл 1983 ; Ксантофита Гибберд 1990 ]
- Специалист по диатомии Дерелл и др. 2016, ред. Кавалер-Смит 2017
- Bolidophyceae Guillou et al. 1999 год
- Diatomeae Dumortier 1821 [=Bacillariophyta Haeckel 1878 ]
- Диктиохофицеа Сильва 1980
- Pelagophyceae Андерсен и Сондерс, 1993 г.
- Pinguiophyceae Kawachi et al. 2003 г.
- Крисиста Кавальер-Смит, 1986 г.
- Бигиромонада Кавальер-Смит 1998 г.
Примечания
[ редактировать ]- ^ Их не называют хлоропластами, это наиболее распространенная форма фотосинтетических пластид. В узком смысле хлоропласт представляет собой пластиду, содержащую хлорофилл B, как в зеленых водорослях , некоторых эвгленидах и наземных растениях .
Ссылки
[ редактировать ]- ^ Перейти обратно: а б ХС Юн; Р.А. Андерсен; СМ Бу; Д. Бхаттачарья (17 февраля 2009 г.). «Страмненопилс» . Энциклопедия микробиологии (третье издание) : 721–731. дои : 10.1016/B978-012373944-5.00253-4 . Проверено 2 марта 2024 г.
- ^ Паттерсон, диджей (1989). «Страменопилес: хромофиты с точки зрения протистана». Ин Грин, Джей Си; Ледбитер, бакалавриат; Дайвер, WL (ред.). Хромофитные водоросли: проблемы и перспективы . Кларендон Пресс. ISBN 978-0198577133 .
- ^ Сина М Адл; Аластер ГБ Симпсон; Марк Фермер; и др. (1 сентября 2005 г.). «Новая классификация эукариот более высокого уровня с упором на таксономию простейших». Журнал эукариотической микробиологии . 52 (5): 399–451. дои : 10.1111/J.1550-7408.2005.00053.X . ISSN 1066-5234 . ПМИД 16248873 . Викиданные Q22065654 .
- ^ Перейти обратно: а б Кавальер-Смит, Томас (январь 2018 г.). «Королевство Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание на перипластидные белки, эволюцию цитоскелета и перипластид, а также древние расхождения» . Протоплазма . 255 (1): 297–357. дои : 10.1007/s00709-017-1147-3 . ПМЦ 5756292 . ПМИД 28875267 .
- ^ Вёрс, Н. (1993). «Морские гетеротрофные амебы, жгутиконосцы и солнечники из Белиза (Центральная Америка) и Тенерифе». Журнал эукариотической микробиологии . 40 (3): 272–287. дои : 10.1111/j.1550-7408.1993.tb04917.x . S2CID 221852241 .
- ^ Дэвид, Джей Си (2002). «Предварительный каталог названий грибов выше порядка» . Констанция . 83 : 1–30.
- ^ Кавальер-Смит, Т. (1999). «Царство Хромиста, происхождение и систематика» . В Раунде, FE; Чепмен, диджей (ред.). Прогресс психологических исследований . Том. 4. Эльзевир. стр. 309–347. ISBN 978-0-948737-00-8 .
- ^ ван ден Хук, К.; Манн, Д.Г.; Янс, Х.М. (1995). Водоросли. Введение в психологию . Издательство Кембриджского университета. ISBN 978-0-521-30419-1 .
- ^ Алексопулос, CJ; Мимс, CW; Блэквелл, М. (1996). Вводная микология (4-е изд.). Уайли. ISBN 978-0471522294 .
- ^ Дик, М.В. (2013). Straminipilous Fungi: Систематика Peronosporomycetes, включая сведения о морских Straminipilous Protists, Plasmodiophorids и подобных организмах . Спрингер. ISBN 978-94-015-9733-3 .
- ^ "Страменипила М.В. Дик (2001)" . МикоБанк . Международная микологическая ассоциация.
- ^ Паттерсон, диджей (1989). «Страменопилес: хромофиты с протистологической точки зрения». Ин Грин, Джей Си; Ледбитер, бакалавриат; Дайвер, WL (ред.). Хромофитные водоросли: проблемы и перспективы . Оксфорд: Кларендон Пресс. стр. 357–379.
- ^ Паттерсон, Дэвид Дж. (1999). «Разнообразие эукариотов». Американский натуралист . 154 (С4): С96–С124. дои : 10.1086/303287 . ПМИД 10527921 . S2CID 4367158 .
- ^ Дэвид, Джей Си (2002). «Предварительный каталог названий грибов выше порядка» . Констанца (83): 1–30.
- ^ Лютер, Александр Ф. (1899). с некоторыми комментариями по систематике родственных водорослей ( О Chlorosaccus, новом роде пресноводных водорослей, на немецком языке). Стокгольм: Норстедт . стр. 1–22.
- ^ Коупленд, Х.Ф. (1956). Классификация низших организмов . Пало-Альто, Калифорния: Pacific Books.
- ^ Блэквелл, Вашингтон (2009). «Возвращение к Хромисте: дилемма перекрытия предполагаемых королевств и попытка применения ботанического номенклатурного кода» (PDF) . Фитология . 91 (2).
- ^ Паттерсон, Колин (1982). «Морфологические признаки и гомологии». В Джойси, Кеннет А.; Пятница, AE (ред.). Проблемы филогенетической реконструкции . Специальный том 21 Ассоциации систематики. Лондон: Academic Press. ISBN 978-0-1239-1250-3 .
- ^ Привет, Вэй И! Блэкман, Лейла М.; Хардхэм, Эдриенн Р. (2019). «Характеристика специфичных для страменопила мастигонемных белков у Phytophthora parasitica». Протоплазма . 256 (2): 521–535. дои : 10.1007/ s00709-018-1314-1 ПМИД 30302550 . S2CID 52947780 .
- ^ Бук, Г. Бенджамин (1 августа 1971 г.). «Строение, происхождение и состав трубчатых мастигонем жгутика Ochromonas » . Журнал клеточной биологии . 50 (2): 362–384. дои : 10.1083/jcb.50.2.362 . ПМК 2108286 . ПМИД 5123323 .
- ^ Блэкман, Лейла М.; Арикава, Микихико; Ямада, Шухэй; Сузаки, Тошинобу; Хардхэм, Эдриенн Р. (2011). «Идентификация белка мастигонемы Phytophthora nicotianae». Протист . 162 (1): 100–114. дои : 10.1016/j.protis.2010.01.005 . ПМИД 20663714 .
- ^ Юн, HS; Андерсен, РА; Бу, СМ; Бхаттачарья, Д. (2009). «Страменопилес». Энциклопедия микробиологии . Эльзевир. стр. 721–731. дои : 10.1016/b978-012373944-5.00253-4 . ISBN 9780123739445 .
- ^ Фу, Банда; Нагасато, Чикаго; Ока, Сейко; Кок, Дж. Марк; Мотомура, Тайдзо (2014). «Протеомный анализ гетерогенных жгутиков бурых водорослей (Stramenopiles)» (PDF) . Протист 165 (5): 662–675. дои : 10.1016/j.protis.2014.07.007 . ПМИД 25150613 . S2CID 7936118 .
- ^ Лейпе, Д.Д.; Уэйнрайт, ПО; Гандерсон, Дж. Х.; и др. (1994). «Страменопилы с молекулярной точки зрения: 16S-подобные последовательности рРНК из Labyrinthuloides minuta и Cafeteria roenbergensis». Психология . 33 (5): 369–377. дои : 10.2216/i0031-8884-33-5-369.1 .
- ^ Лейланд, Бен; Леу, Стефан; Буссиба, Сэмми (2017). «Являются ли траустохитриды водорослями?». Грибковая биология . 121 (10): 835–840. дои : 10.1016/j.funbio.2017.07.006 . ПМИД 28889907 .
- ^ Перейти обратно: а б Дерелл, Роман; Лопес-Гарсия, «Очищение»; Тимпано, Хелен; Морейра, Дэвид (10 августа 2016 г.). «Филогеномная основа для изучения разнообразия и эволюции страменопилов (= гетероконтов) » Молекулярная биология и эволюция . 33 (11): 2890–2898. дои : 10.1093/molbev/msw168 . ПМЦ 5482393 . ПМИД 27512113 .
- ^ Робертс, Тамали; Старк, Дэмиен; Харкнесс, Джон; Эллис, Джон (2014). «Обновленная информация о патогенном потенциале и вариантах лечения Blastocystis sp» . Кишечные патогены . 6 (1): 17. дои : 10.1186/1757-4749-6-17 . ПМК 4039988 . ПМИД 24883113 .
- ^ Олсен, О. Уилфорд (1986). Паразиты животных: их жизненные циклы и экология . Нью-Йорк: Дувр. стр. 56, 74–75. ISBN 0-486-65126-6 . OCLC 13123309 .
- ^ Новицкий, Марцин; и др. (17 августа 2011 г.). «Фитофтороз картофеля и томатов, вызываемый Phytophthora infestans : обзор патологии и селекции устойчивости» . Болезни растений . 96 (1). Американское фитопатологическое общество : 4–17. дои : 10.1094/PDIS-05-11-0458 . ПМИД 30731850 .
- ^ Юл, Эндрю; Тиррелл, Тоби (2003). «Роль диатомей в регулировании круговорота кремния в океане». Глобальные биогеохимические циклы . 17 (4): н/д. Бибкод : 2003GBioC..17.1103Y . CiteSeerX 10.1.1.394.3912 . дои : 10.1029/2002GB002018 . S2CID 16849373 .
- ^ Кок, Дж. Марк; Питерс, Акира Ф.; Коэльо, Сусана М. (9 августа 2011 г.). «Бурые водоросли» . Современная биология . 21 (15): R573–R575. дои : 10.1016/j.cub.2011.05.006 . ПМИД 21820616 .
- ^ Гири, Венди (7 апреля 2011 г.). «Кафетерий Т. Фенчел и DJ Паттерсон 1988» . База водорослей . Проверено 17 марта 2023 г.
- ^ Фенчел, Т.; Паттерсон, диджей (1988). « Cateria roenbergensis nov. gen., nov. sp., гетеротрофная микрофлагеллята из морского планктона» . Морские микробные пищевые сети . 3 : 9–19.
- ^ Krylov, M. V.; Dobrovolskii, A. A.; Issi, I. V.; Michaelevich, B. I.; Podlipaev, S. A.; Reshetnyak, V. V.; Seravin, L. N.; et al. 1980. New concepts for the system of unicellular organisms. Trudy Zoologischkei Institut Akademiya Nayuk, SSSR 94:122–132.
- ^ Бурки, Ф.; Шалчиан-Тебризи; Павловский, Дж. (август 2008 г.). «Филогеномика открывает новую «мегагруппу», включающую большинство фотосинтезирующих эукариот» . Письма по биологии . 4 (4): 366–369. дои : 10.1098/rsbl.2008.0224 . ПМК 2610160 . ПМИД 18522922 .
- ^ Оборник, Мирослав; Лукеш, Юлиус (2013). «Клеточная биология хромридов». Международное обозрение клеточной и молекулярной биологии . Том. 306. Эльзевир . стр. 333–369. дои : 10.1016/b978-0-12-407694-5.00008-0 . ISBN 9780124076945 . ПМИД 24016529 .
- ^ Tikhonenkov, Denis V.; Jamy, Mahwash; Borodina, Anastasia S.; Belyaev, Artem O.; Zagumyonnyi, Dmitry G.; Prokina, Kristina I.; Mylnikov, Alexander P.; Burki, Fabien; Karpov, Sergey A. (2022). "On the origin of TSAR: morphology, diversity and phylogeny of Telonemia" . Open Biology . 12 (3). The Royal Society. doi : 10.1098/rsob.210325 . ISSN 2046-2441 . PMC 8924772 . PMID 35291881 .
- ^ Перейти обратно: а б Кавалер-Смит, Томас; Скобл, Жозефина Маргарет (2013). «Филогения Heterokonta: Incisomonas marina, одноресничный скользящий опалозой, родственный Solenicola (Nanomonadea), и свидетельство того, что Actinophryida произошла от рафидофитов». Европейский журнал протистологии . 49 (3): 328–353. дои : 10.1016/j.ejop.2012.09.002 . ПМИД 23219323 .
- ^ Перейти обратно: а б с Тхакур, Рабиндра; Ширатори, Такаши; Исида, Кен-итиро (2019). «Мультигенный филогенетический анализ, богатый таксонами, разрешает филогенетические отношения между страменопилами с глубоким разветвлением» . Протист . 170 (5): 125682. doi : 10.1016/j.protis.2019.125682 . ISSN 1434-4610 . PMID 31568885 . S2CID 202865459 .
- ^ Перейти обратно: а б с Ширатори, Такаши; Тхакур, Рабиндра; Исида, Кен-итиро (2017). «Pseudophyllomitus vesiculosus (Ларсен и Паттерсон, 1990) Ли, 2002, плохо изученная фаготрофная двужгутиковая пластинка является первым охарактеризованным членом страменопильной экологической группы MAST-6» . Протист . 168 (4): 439–451. дои : 10.1016/j.protis.2017.06.004 . ISSN 1434-4610 . ПМИД 28822908 .
- ^ Перейти обратно: а б Элизабет Дж. Уэстон; Черный Эглит; Аластер ГБ Симпсон (6 октября 2023 г.). «Kaonashia insperata gen. et sp. nov., эукариотрофный жгутиконосец, представляет собой новую основную линию гетеротрофных страменопилей». Журнал эукариотической микробиологии . дои : 10.1111/ЧЕТВЕРГ.13003 . ISSN 1066-5234 . Викиданные, первый квартал
- ^ Анна Чо; Денис Владимирович Тихоненков; Гордон Лакс; Кристина Ивановна Прокина; Патрик Дж. Килинг (10 ноября 2023 г.). «Филогеномное положение генетически разнообразных фаготрофных жгутиконосцев страменопилей в связанной с отложениями линии MAST-6 и потенциально галотолерантных плацидид». Молекулярная филогенетика и эволюция . 190 : 107964. doi : 10.1016/J.YMPEV.2023.107964 . ISSN 1055-7903 . ПМИД 37951557 . Викиданные Q124719861 .
- ^ Адл С.М., Басс Д., Лейн CE, Лукеш Дж., Шох К.Л., Смирнов А., Агата С., Берни С., Браун М.В., Бурки Ф., Карденас П., Чепичка И., Чистякова Л., дель Кампо Дж., Данторн М., Эдвардсен Б., Эглит. Ю, Гийу Л., Хампл В., Хейсс А.А., Хоппенрат М., Джеймс Тай, Карнковска А., Карпов С., Ким Е., Колиско М., Кудрявцев А., Лар DJG, Лара Е., Ле Галл Л., Линн Д.Х., Манн Д.Г., Массана Р. , Митчелл И.А.Д., Морроу С., Парк Дж.С., Павловски Дж.В., Пауэлл М.Дж., Рихтер Д.Д., Рюкерт С., Шедвик Л., Шимано С., Шпигель Ф.В., Торруэлла Г., Юсеф Н., Златогурский В., Чжан К. (2019). «Пересмотр классификации, номенклатуры и разнообразия эукариот» . Журнал эукариотической микробиологии . 66 (1): 4–119. дои : 10.1111/jeu.12691 . ПМК 6492006 . ПМИД 30257078 .
- ^ Уэтерби Р., Джексон С.Дж., Репетти С.И., Клементсон Л.А., Коста Дж.Ф., ван де Мин А., Кроуфорд С., Вербрюгген Х. (апрель 2019 г.). «Золотой парадокс - новая линия гетероконтов с хлоропластами, окруженными двумя мембранами». Дж. Фикол . 55 (2): 257–278. дои : 10.1111/jpy.12822 . hdl : 11343/233613 . ПМИД 30536815 . S2CID 54477112 .
- ^ Медлин Л.К., Десдевайс Ю (2018). «Анализ последовательности подтверждает новый класс водорослей» . Vie et Milieu/Жизнь и окружающая среда .
- ^ Граф, Луи; Юн, Хван Су (21 июля 2021 г.). «Olisthodiscophyceae, 17-й класс гетероконтных водорослей». Журнал психологии . 57 (4): 1091–1093. дои : 10.1111/jpy.13184 . ПМИД 34289104 . S2CID 236175098 .
- ^ Барците, Довиле; Эйкрем, Венче; Энгесмо, Анетт; Сеоан, Серхио; Вольманн, Йенс; Горак, Алесь; Юрченко Татьяна; Элиаш, Марек (2 марта 2021 г.). « Олистодиск представляет собой новый класс Ochrophyta» . Журнал психологии . 57 (4): 1094–1118. дои : 10.1111/jpy.13155 . hdl : 10852/86515 . ПМИД 33655496 . S2CID 232101186 .
- ^ Граф Л., Ян Э.К., Хан К.Ю., Кюппер ФК, Бенеш К.М., Оядомари Дж.К., Герберт Р.Дж.Х., Вербрюгген Х., Уэтерби Р., Андерсен Р.А., Юн Х.С. (декабрь 2020 г.). «Мультигенная филогения, морфологические наблюдения и пересмотр литературы привели к описанию Phaeosacciophyceae Classis Nova и четырех новых видов Heterokontophyta SI Clade» . Протист . 171 (6): 125781. doi : 10.1016/j.protis.2020.125781 . ПМИД 33278705 . S2CID 227315556 .