Биологическая мембрана

, Биологическая мембрана биомембрана или клеточная мембрана — это избирательно проницаемая мембрана , которая отделяет внутреннюю часть клетки от внешней среды или создает внутриклеточные отсеки , служа границей между одной частью клетки и другой. Биологические мембраны в форме мембран эукариотических клеток состоят из фосфолипидного бислоя со встроенными, интегральными и периферическими белками, используемыми для передачи и транспортировки химических веществ и ионов . Основная масса липидов в клеточной мембране обеспечивает жидкую матрицу, позволяющую белкам вращаться и латерально диффундировать для физиологического функционирования. Белки приспособлены к высокой мембранной текучести среды липидного бислоя с наличием кольцевой липидной оболочки , состоящей из липидных молекул, прочно связанных с поверхностью целостных мембранных белков . Клеточные мембраны отличаются от изолирующих тканей, образованных слоями клеток, таких как слизистые оболочки , базальные мембраны и серозные оболочки .
Состав
[ редактировать ]Асимметрия
[ редактировать ]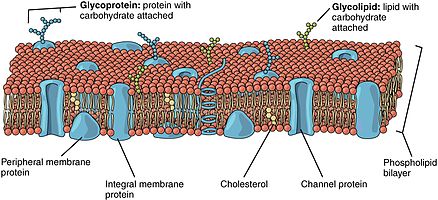
Липидный бислой состоит из двух слоев: внешнего листка и внутреннего листка. [1] Компоненты бислоев распределены между двумя поверхностями неравномерно, что создает асимметрию между внешней и внутренней поверхностями. [2] Эта асимметричная организация важна для клеточных функций, таких как передача сигналов клетки. [3] Асимметрия биологической мембраны отражает разные функции двух створок мембраны. [4] Как видно на модели жидкостной мембраны фосфолипидного бислоя, внешний и внутренний листочки мембраны асимметричны по своему составу. Некоторые белки и липиды располагаются только на одной поверхности мембраны, а не на другой.
• И плазматическая мембрана, и внутренние мембраны имеют цитозольную и экзоплазматическую поверхности.• Эта ориентация сохраняется во время мембранного транспорта – белки, липиды, гликоконъюгаты, обращенные к просвету ЭР и Гольджи, экспрессируются на внеклеточной стороне плазматической мембраны. В эукариотических клетках новые фосфолипиды производятся ферментами, связанными с той частью мембраны эндоплазматического ретикулума, которая обращена к цитозолю. [5] Эти ферменты, которые используют свободные жирные кислоты в качестве субстратов , откладывают все вновь образовавшиеся фосфолипиды в цитозольную половину бислоя. Чтобы мембрана в целом могла расти равномерно, половина новых молекул фосфолипидов должна быть перенесена на противоположный монослой. Этот перенос катализируется ферментами, называемыми флипазами . В плазматической мембране флипазы избирательно переносят специфические фосфолипиды, так что в каждом монослое концентрируются разные типы. [5]
Однако использование селективных флипаз — не единственный способ вызвать асимметрию липидных бислоев. В частности, другой механизм действует для гликолипидов — липидов, которые демонстрируют наиболее яркое и последовательное асимметричное распределение в клетках животных . [5]
Липиды
[ редактировать ]Биологическая мембрана состоит из липидов с гидрофобными хвостами и гидрофильными головками. [6] Гидрофобные хвосты представляют собой углеводородные хвосты, длина и насыщенность которых важны для характеристики клетки. [7] Липидные рафты возникают, когда виды липидов и белки агрегируются в доменах мембраны. Они помогают организовать мембранные компоненты в локализованные области, которые участвуют в определенных процессах, таких как передача сигнала.
Красные кровяные тельца, или эритроциты, имеют уникальный липидный состав. Бислой эритроцитов состоит из холестерина и фосфолипидов в равных весовых пропорциях. [7] Мембрана эритроцитов играет решающую роль в свертывании крови. В бислое эритроцитов находится фосфатидилсерин. [8] Обычно это находится на цитоплазматической стороне мембраны. Однако он переворачивается на внешнюю мембрану и используется при свертывании крови. [8]
Белки
[ редактировать ]Фосфолипидные бислои содержат разные белки. Эти мембранные белки имеют различные функции и характеристики и катализируют различные химические реакции. Интегральные белки охватывают мембраны с разными доменами с каждой стороны. [6] Интегральные белки прочно связаны с липидным бислоем и не могут легко отсоединиться. [9] Они диссоциируют только при химической обработке, разрушающей мембрану. Периферические белки отличаются от интегральных белков тем, что они слабо взаимодействуют с поверхностью бислоя и могут легко диссоциировать от мембраны. [6] Периферические белки расположены только на одной стороне мембраны и создают асимметрию мембраны.
| ФУНКЦИОНАЛЬНЫЙ КЛАСС | ПРИМЕР БЕЛКА | СПЕЦИАЛЬНАЯ ФУНКЦИЯ |
|---|---|---|
| Транспортеры | На+ Насос | активно выкачивает Na+ из клеток и K+ в |
| Якоря | интегрины | связывают внутриклеточные актиновые филаменты с белками внеклеточного матрикса |
| Рецепторы | фактора роста тромбоцитов рецептор | связывает внеклеточный PDGF и, как следствие, генерирует внутриклеточные сигналы, которые заставляют клетку расти и делиться |
| Ферменты | аденилатциклаза | катализирует выработку внутриклеточной сигнальной молекулы циклического АМФ в ответ на внеклеточные сигналы |
Олигосахариды
[ редактировать ]Олигосахариды представляют собой сахаросодержащие полимеры. В мембране они могут быть ковалентно связаны с липидами с образованием гликолипидов или ковалентно связаны с белками с образованием гликопротеинов . Мембраны содержат сахаросодержащие липидные молекулы, известные как гликолипиды. В бислое сахарные группы гликолипидов выходят на поверхность клетки, где они могут образовывать водородные связи. [9] Гликолипиды представляют собой наиболее крайний пример асимметрии липидного бислоя. [10] Гликолипиды выполняют в биологической мембране огромное количество функций, в основном коммуникативных, включая распознавание клеток и межклеточную адгезию. Гликопротеины являются интегральными белками. [2] Они играют важную роль в иммунном ответе и защите. [11]
Формирование
[ редактировать ]Фосфолипидный бислой образуется за счет агрегации мембранных липидов в водных растворах. [4] Агрегация вызвана гидрофобным эффектом , при котором гидрофобные концы вступают в контакт друг с другом и изолируются от воды. [6] Такое расположение максимизирует водородные связи между гидрофильными головками и водой, сводя к минимуму неблагоприятный контакт между гидрофобными хвостами и водой. [10] Увеличение количества доступных водородных связей увеличивает энтропию системы, создавая самопроизвольный процесс.
Функция
[ редактировать ]Биологические молекулы бывают амфифильными или амфипатическими, т.е. одновременно гидрофобными и гидрофильными. [6] Фосфолипидный бислой содержит заряженные гидрофильные головные группы, которые взаимодействуют с полярной водой . Слои также содержат гидрофобные хвосты, которые встречаются с гидрофобными хвостами комплементарного слоя. Гидрофобные хвосты обычно представляют собой жирные кислоты разной длины. [10] Взаимодействия такие липидов, особенно гидрофобных хвостов, определяют физические свойства липидного бислоя, как текучесть.
Мембраны в клетках обычно определяют замкнутые пространства или отсеки, в которых клетки могут поддерживать химическую или биохимическую среду, отличную от внешней. Например, мембрана вокруг пероксисом защищает остальную часть клетки от пероксидов, химических веществ, которые могут быть токсичными для клетки, а клеточная мембрана отделяет клетку от окружающей ее среды. Пероксисомы — это одна из форм вакуолей, обнаруженных в клетке, которые содержат побочные продукты химических реакций внутри клетки. Большинство органелл ограничены такими мембранами и называются мембраносвязанными органеллами .
Селективная проницаемость
[ редактировать ]Вероятно, наиболее важной особенностью биомембраны является то, что она представляет собой структуру с избирательной проницаемостью. Это означает, что размер, заряд и другие химические свойства атомов и молекул, пытающихся пересечь его, будут определять, удастся ли им это сделать. Избирательная проницаемость необходима для эффективного отделения клетки или органеллы от окружающей среды. Биологические мембраны также обладают определенными механическими или эластичными свойствами, которые позволяют им менять форму и двигаться по мере необходимости.
Как правило, небольшие гидрофобные молекулы могут легко пересекать фосфолипидные бислои путем простой диффузии . [12]
Частицы, которые необходимы для клеточной функции, но не могут свободно диффундировать через мембрану, проникают через мембранный транспортный белок или поглощаются посредством эндоцитоза , когда мембрана позволяет вакуоли присоединиться к ней и вытолкнуть ее содержимое в клетку. От внешней среды клетку могут отделять специализированные плазматические мембраны многих типов: апикальные, базолатеральные, пресинаптические и постсинаптические, мембраны жгутиков, ресничек, микроворсинок , филоподий и ламеллиподий , сарколеммы мышечных клеток, а также специализированные миелиновые и дендритные мембраны шипиков. нейроны. Плазматические мембраны также могут образовывать различные типы «надмембранных» структур, таких как кавеолы , постсинаптическая плотность, подосома , инвадоподий , десмосома, гемидесмосома , фокальная адгезия и клеточные соединения. Эти типы мембран различаются по липидному и белковому составу.
Различные типы мембран также образуют внутриклеточные органеллы: эндосомы; гладкая и шероховатая эндоплазматическая сеть; саркоплазматическая сеть; аппарат Гольджи; лизосома; митохондрии (внутренняя и внешняя мембраны); ядро (внутренняя и внешняя мембраны); пероксисома ; вакуоль; цитоплазматические гранулы; клеточные везикулы (фагосомы, аутофагосомы , клатрином везикулы, покрытые , везикулы, покрытые COPI и COPII ) и секреторные везикулы (включая синаптосомы , акросомы , меланосомы и хромаффинные гранулы).Различные типы биологических мембран имеют разнообразный липидный и белковый состав. Состав мембран определяет их физические и биологические свойства. Некоторые компоненты мембран играют ключевую роль в медицине, например, откачивающие насосы, которые выкачивают лекарства из клетки.
Текучесть
[ редактировать ]Гидрофобное ядро фосфолипидного бислоя постоянно находится в движении за счет вращения вокруг связей липидных хвостов. [13] Гидрофобные хвосты бислоя изгибаются и сцепляются между собой. Однако из-за водородных связей с водой гидрофильные головные группы проявляют меньшее движение, поскольку их вращение и подвижность ограничены. [13] Это приводит к увеличению вязкости липидного бислоя ближе к гидрофильным головкам. [6]
Ниже температуры перехода липидный бислой теряет текучесть, когда высокоподвижные липиды проявляют меньше движения и превращаются в гелеобразное твердое вещество. [14] Температура перехода зависит от таких компонентов липидного бислоя, как длина углеводородной цепи и насыщенность его жирными кислотами. Текучесть, зависящая от температуры, является важным физиологическим свойством бактерий и хладнокровных организмов. Эти организмы поддерживают постоянную текучесть, изменяя состав жирных кислот мембранных липидов в соответствии с различными температурами. [6]
В клетках животных текучесть мембран модулируется за счет включения стерина- холестерола . Эта молекула присутствует в особенно больших количествах в плазматической мембране, где она составляет примерно 20% липидов мембраны по массе. Поскольку молекулы холестерина короткие и жесткие, они заполняют пространства между соседними молекулами фосфолипидов, оставленные изломами в их ненасыщенных углеводородных хвостах. Таким образом, холестерин имеет тенденцию придавать жесткость бислою, делая его более жестким и менее проницаемым. [5]
Для всех клеток текучесть мембран важна по многим причинам. Это позволяет мембранным белкам быстро диффундировать в плоскости бислоя и взаимодействовать друг с другом, что имеет решающее значение, например, для передачи сигналов в клетках . Это позволяет мембранным липидам и белкам диффундировать из мест, где они встраиваются в бислой после их синтеза, в другие области клетки. Он позволяет мембранам сливаться друг с другом и смешивать свои молекулы, а также обеспечивает равномерное распределение молекул мембраны между дочерними клетками при делении клетки. Если бы биологические мембраны не были жидкими, трудно представить, как клетки могли бы жить, расти и размножаться. [5]
Свойство текучести находится в центре модели Хелфриха , которая позволяет рассчитать энергетические затраты на упругую деформацию мембраны.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Мурате, Мотохидэ; Кобаяси, Тошихидэ (2016). «Возвращаясь к трансбислойному распределению липидов в плазматической мембране». Химия и физика липидов . 194 : 58–71. doi : 10.1016/j.chemphyslip.2015.08.009 . ПМИД 26319805 .
- ^ Перейти обратно: а б Никельс, Джонатан Д.; Смит, Джереми К.; Ченг, Сяолинь (2015). «Латеральная организация, бислойная асимметрия и межстворчатое соединение биологических мембран» . Химия и физика липидов . 192 : 87–99. doi : 10.1016/j.chemphyslip.2015.07.012 . ПМИД 26232661 .
- ^ Чонг, Чжи-Сун; Ву, Вэй-Фен; Чнг, Шу-Син (01 декабря 2015 г.). «Осмопорин OmpC образует комплекс с MlaA для поддержания асимметрии липидов внешней мембраны Escherichia coli» . Молекулярная микробиология . 98 (6): 1133–1146. дои : 10.1111/mmi.13202 . ПМИД 26314242 .
- ^ Перейти обратно: а б Форрест, Люси Р. (1 января 2015 г.). «Структурная симметрия мембранных белков» . Ежегодный обзор биофизики . 44 (1): 311–337. doi : 10.1146/annurev-biophys-051013-023008 . ПМК 5500171 . ПМИД 26098517 .
- ^ Перейти обратно: а б с д и Альбертс, Брэй, Хопкин, Джонсон, Льюис, Рафф, Робертс, Уолтер, Брюс, Деннис, Карен, Александр, Джулиан, Мартин, Кейт, Питер (2010). «Основная клеточная биология», третье издание . Нью-Йорк: Garland Science, Taylor & Francisco Group, LLC, информационный бизнес. п. 370. ИСБН 978-0815341291 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б с д и ж г Воэт, Дональд (2012). Основы биохимии: Жизнь на молекулярном уровне (4-е изд.) . Уайли. ISBN 978-1118129180 .
- ^ Перейти обратно: а б Догерти, Р.М.; Галли, К.; Ферро-Луцци, А.; Яконо, Дж. М. (1987). «Липидный и фосфолипидно-жирнокислотный состав плазмы, эритроцитов и тромбоцитов и влияние на них пищевых липидов: исследование нормальных субъектов из Италии, Финляндии и США». Американский журнал клинического питания . 45 (2): 443–455. дои : 10.1093/ajcn/45.2.443 . ПМИД 3812343 . S2CID 4436467 .
- ^ Перейти обратно: а б Ленц, Барри Р. (2003). «Воздействие фосфатидилсерина на мембраны тромбоцитов регулирует свертывание крови». Прогресс в исследованиях липидов . 42 (5): 423–438. дои : 10.1016/s0163-7827(03)00025-0 . ПМИД 12814644 .
- ^ Перейти обратно: а б Лейн, Макс; деРонд, Бриттани М.; Сголастра, Федерика; Тью, Грегори Н.; Холден, Мэтью А. (1 ноября 2015 г.). «Транспорт белков через мембраны: сравнение между носителями, богатыми лизином и гуанидинием» . Biochimica et Biophysical Acta (BBA) – Биомембраны . 1848 (11, Часть А): 2980–2984. дои : 10.1016/j.bbamem.2015.09.004 . ПМЦ 4704449 . ПМИД 26342679 .
- ^ Перейти обратно: а б с Альбертс, Брюс; Джонсон, Александр; Льюис, Джулиан; Рафф, Мартин; Робертс, Кейт; Уолтер, Питер (1 января 2002 г.). «Липидный бислой» .
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Добенспек, Джеймс М.; Джордан, Дэвид С.; Симмонс, Уоррен; Ренфроу, Мэтью Б.; Дибвиг, Кевин (23 ноября 2015 г.). «Общее N- и О-связанное гликозилирование липопротеинов в микоплазмах и роль экзогенных олигосахаридов» . ПЛОС ОДИН . 10 (11): e0143362. Бибкод : 2015PLoSO..1043362D . дои : 10.1371/journal.pone.0143362 . ПМК 4657876 . ПМИД 26599081 .
- ^ Браун, Бернард (1996). Биологические мембраны (PDF) . Лондон, Великобритания: Биохимическое общество. п. 21. ISBN 978-0904498325 . Архивировано из оригинала (PDF) 6 ноября 2015 г. Проверено 1 мая 2014 г.
- ^ Перейти обратно: а б Витрак, Хайди; Маклин, Дэвид М.; Джаяраман, Васанти; Богданов Михаил; Доухан, Уильям (10 ноября 2015 г.). «Динамическое топологическое переключение мембранных белков при изменении фосфолипидного окружения» . Труды Национальной академии наук . 112 (45): 13874–13879. Бибкод : 2015PNAS..11213874V . дои : 10.1073/pnas.1512994112 . ПМЦ 4653158 . ПМИД 26512118 .
- ^ Ройко, Нейц; Андерлу, Грегор (07 декабря 2015 г.). «Как липидные мембраны влияют на активность порообразующих токсинов». Отчеты о химических исследованиях . 48 (12): 3073–3079. дои : 10.1021/acs.accounts.5b00403 . ПМИД 26641659 .
Внешние ссылки
[ редактировать ] СМИ, связанные с биологическими мембранами , на Викискладе?
СМИ, связанные с биологическими мембранами , на Викискладе? - Мембраны в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
| Базы данных органов управления : Национальные |
|---|