Фракционирование изотопов углерода при оксигенном фотосинтезе
Фотосинтез преобразует углекислый газ в углеводы посредством нескольких метаболических путей которые обеспечивают организм энергией и преимущественно реагируют с определенными стабильными изотопами углерода , . [ 1 ] Избирательное обогащение одного стабильного изотопа над другим создает различные изотопные фракции , которые можно измерить и соотнести между оксигенными фототрофами . На степень фракционирования изотопов углерода влияют несколько факторов, включая метаболизм, анатомию, скорость роста и условия окружающей среды организма. Понимание этих различий в фракционировании углерода между видами полезно для биогеохимических исследований, включая реконструкцию палеоэкологии , эволюции растений и характеристики пищевых цепей . [ 2 ] [ 3 ]

Кислородный фотосинтез — это метаболический путь , которому способствуют автотрофы , включая растения, водоросли и цианобактерии. Этот путь преобразует неорганический углекислый газ из атмосферы или водной среды в углеводы , используя воду и энергию света, а затем выделяет в качестве продукта молекулярный кислород. Органический углерод содержит меньше стабильного изотопа углерода-13 , или 13 C относительно исходного неорганического углерода из атмосферы или воды, поскольку фотосинтетическая фиксация углерода включает несколько реакций фракционирования с кинетическими изотопными эффектами . [ 4 ] Эти реакции подвергаются кинетическому изотопному эффекту, поскольку они ограничиваются преодолением энергетического барьера активации . Более легкий изотоп имеет более высокое энергетическое состояние в квантовой яме химической связи, что позволяет ему преимущественно образовывать продукты. Разные организмы фиксируют углерод с помощью разных механизмов, что отражается в различном изотопном составе в разных путях фотосинтеза (см. таблицу ниже и объяснение обозначений в разделе «Измерение изотопов углерода»). В следующих разделах будут описаны различные пути оксигенного фотосинтеза и то, что влияет на связанные с ними значения дельты.

| Путь | д 13 С (‰) |
|---|---|
| С3 | от -20 до -37 [ 2 ] |
| С4 | от -12 до -16 [ 5 ] |
| САМ | от -10 до -20 [ 6 ] |
| Фитопланктон | от -18 до -25 [ 4 ] [ 7 ] |
Измерение изотопов углерода
[ редактировать ]Углерод на Земле естественным образом встречается в виде двух стабильных изотопов, причем 98,9% находится в форме 12 С и 1,1% в 13 С. [ 1 ] [ 8 ] Соотношение между этими изотопами варьируется в биологических организмах из-за метаболических процессов, в которых один изотоп углерода избирательно используется вместо другого или «фракционируется» углерод посредством кинетических или термодинамических эффектов. [ 1 ] Кислородный фотосинтез происходит у растений и микроорганизмов разными химическими путями, поэтому различные формы органического материала отражают разные соотношения 13 С. Изотопы Понимание этих различий в фракционировании углерода между видами применяется в изотопной геохимии и экологических изотопных исследованиях для понимания биохимических процессов, установления пищевых цепей или моделирования углеродного цикла в геологическом времени. [ 5 ]
Фракционирование изотопов углерода выражается в виде дельта-обозначения δ. 13 C («дельта тринадцать C»), который выражается в тысячных частях ( промилле , ‰). [ 9 ] д 13 C определяется по отношению к белемниту Венского Пи Ди (VPDB, 13 С/ 12 C = 0,01118) в качестве установленного эталонного стандарта . [ 8 ] [ 10 ] Это называется «значением дельты» и может быть рассчитано по формуле ниже:

Реакции фотосинтеза
[ редактировать ]Химический путь оксигенного фотосинтеза связывает углерод в два этапа: светозависимые реакции и светонезависимые реакции.
Светозависимые реакции улавливают энергию света для переноса электронов из воды и преобразования НАДФ. + , АДФ и неорганический фосфат в запасающие энергию молекулы НАДФН и АТФ . Общее уравнение светозависимых реакций обычно выглядит так: [ 11 ]
2 Н 2 О + 2 НАДФ + + 3 АДФ + 3 П i + свет → 2 НАДФН + 2 Н + + 3 АТФ + О 2
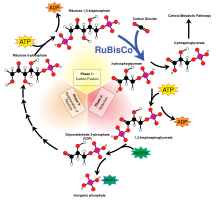
Светонезависимые цикл Кальвина реакции проходят -Бенсона , в котором энергия НАДФН и АТФ используется для преобразования углекислого газа и воды в органические соединения с помощью фермента RuBisCO . Общее уравнение светонезависимых реакций выглядит следующим образом: [ 11 ]
3 СО 2 + 9 АТФ + 6 НАДФН + 6 Н + → C 3 H 6 O 3 -фосфат + 9 АДФ + 8 Р i + 6 НАДФ + + 3 Н 2 О
3-углеродные продукты (C 3 H 6 O 3 -фосфат) цикла Кальвина позже превращаются в глюкозу или другие углеводы, такие как крахмал , сахароза и целлюлоза .
Фракционирование через RuBisCO
[ редактировать ]
Большая фракция 13 С при фотосинтезе происходит за счет реакции карбоксилирования, которую осуществляет фермент рибулозо-1,5-бисфосфаткарбоксилаза оксигеназа, или RuBisCO . [ 5 ] RuBisCO катализирует реакцию между пятиуглеродной молекулой рибулозо-1,5-бисфосфата (сокращенно RuBP) и CO 2 с образованием двух молекул 3-фосфоглицериновой кислоты (сокращенно PGA). PGA реагирует с НАДФН с образованием 3-фосфоглицеральдегида . [ 4 ]
По прогнозам, фракционирование изотопов за счет одного лишь карбоксилирования Рубиско (форма I) приведет к обеднению в среднем на 28 ‰. [ 12 ] [ 5 ] Однако значения фракционирования различаются между организмами: от истощения на 11 ‰, наблюдаемого у кокколитофоридных водорослей, до истощения на 29 ‰, наблюдаемого у шпината . [ 13 ] [ 14 ] RuBisCO вызывает кинетический изотопный эффект, потому что 12 СО 2 и 13 CO 2 конкурируют за один и тот же активный центр и 13 C имеет изначально более низкую скорость реакции. [ 15 ]
13 Модель фракционирования C
[ редактировать ]Помимо избирательных эффектов ферментативных реакций, диффузия газообразного CO 2 к месту карбоксилирования внутри растительной клетки также влияет на фракционирование изотопов. [ 16 ] В зависимости от вида растения (см. разделы ниже) внешний CO 2 должен транспортироваться через пограничный слой и устьица во внутреннее газовое пространство растительной клетки, где он растворяется и диффундирует к хлоропластам. [ 5 ] Коэффициент диффузии газа обратно пропорционален квадратному корню из его молекулярной приведенной массы (по отношению к воздуху), что приводит к 13 CO 2 должен быть на 4,4‰ менее диффузионным, чем 12 СО 2 .
Преобладающая модель фракционирования атмосферного CO 2 в растениях объединяет изотопные эффекты реакции карбоксилирования с изотопными эффектами диффузии газа в растение в следующем уравнении: [ 16 ]

Где:
- д 13 C Образец представляет собой дельта-значение организма для 13 Состав С
- д 13 C atm — это дельта-значение атмосферного CO 2 , которое составляет = -7,8‰.
- дискриминация за счет диффузии a = 4,4‰
- дискриминация карбоксилирования b = 30‰
- c a — парциальное давление CO 2 во внешней атмосфере, а
- c i — парциальное давление CO 2 в межклеточном пространстве.
Эта модель, полученная ab initio , обычно описывает фракционирование углерода в большинстве растений, которое облегчает фиксацию углерода C3 . В эту модель были внесены изменения с учетом эмпирических данных. [ 17 ] Однако несколько дополнительных факторов, не включенных в эту общую модель, будут увеличивать или уменьшать 13 Фракционирование углерода по видам. К таким факторам относятся конкурирующая реакция оксигенации RuBisCO, анатомическая и временная адаптация к активности фермента, а также изменения в росте и геометрии клеток. Изотопное фракционирование различных путей фотосинтеза уникально характеризуется этими факторами, как описано ниже.
На растениях C3
[ редактировать ]
Растение C3 использует фиксацию углерода C3 , один из трех метаболических путей фотосинтеза, которые также включают C4 и CAM (описанные ниже). Эти растения получили название «С3» из-за трехуглеродного соединения ( 3-фосфоглицериновой кислоты , или 3-ФГК), вырабатываемого CO 2 в этих растениях в результате механизма фиксации . Этот механизм C3 является первым этапом цикла Кальвина-Бенсона, который превращает CO 2 и RuBP в 3-PGA .
Растения C3 являются наиболее распространенным типом растений и обычно хорошо себя чувствуют при умеренной интенсивности солнечного света и температуре, концентрации CO 2 выше 200 частей на миллион и обильных грунтовых водах. [ 18 ] Растения C3 плохо растут в очень жарких или засушливых регионах, где растения C4 и CAM лучше адаптированы.
Фракционирование изотопов при фиксации углерода C3 возникает в результате комбинированного воздействия диффузии газа CO 2 через устьица растения и карбоксилирования через RuBisCO . [ 1 ] Устьичная проводимость различает более тяжелые 13 С на 4,4‰. [ 1 ] Карбоксилирование RuBisCO способствует большей дискриминации - 27 ‰. [ 1 ]
Фермент RuBisCO катализирует карбоксилирование CO 2 и 5-углеродного сахара RuBP в 3-фосфоглицерат , 3-углеродное соединение, посредством следующей реакции:
![{\displaystyle CO_{2}+H_{2}O+RuBP{\xrightarrow[{RuBisCO}]{}}2(3{\mbox{-}}{\text{фосфоглицерат}})}](https://wikimedia.org/api/rest_v1/media/math/render/svg/122f351ccccb56a3fed9a5e94d25a15f369918d9)
Продукт 3-фосфоглицерат обеднен 13 C из-за кинетического изотопного эффекта указанной выше реакции. Общий 13 Фракционирование C для фотосинтеза C3 колеблется от -20 до -37 ‰. [ 2 ]
Широкий диапазон изменений значений дельта, выраженных у растений C3, модулируется устьичной проводимостью или скоростью поступления CO 2 или выхода водяного пара из небольших пор в эпидермисе листа. [ 1 ] д 13 C растений C3 зависит от взаимосвязи между устьичной проводимостью и скоростью фотосинтеза, что является хорошим показателем эффективности использования воды в листе. [ 19 ] Установки C3 с высокой эффективностью использования воды, как правило, менее фракционированы в 13 C (т.е. δ 13 C относительно менее отрицателен) по сравнению с растениями C3 с низкой эффективностью использования воды. [ 19 ]
В растениях С4
[ редактировать ]
Растения C4 разработали путь фиксации углерода C4 для сохранения потери воды, поэтому они более распространены в жарком, солнечном и сухом климате. [ 20 ] Эти растения отличаются от растений C3 тем, что CO 2 первоначально преобразуется в четырехуглеродную молекулу малата , которая перемещается в клетки оболочки пучка, высвобождается обратно в виде CO 2 и только затем входит в цикл Кальвина. Напротив, растения C3 непосредственно выполняют цикл Кальвина в клетках мезофилла, не используя метод концентрации CO 2 . Малат, четырехуглеродное соединение, является тезкой фотосинтеза «C4». Этот путь позволяет фотосинтезу C4 эффективно переносить CO 2 к ферменту RuBisCO и поддерживать высокие концентрации CO 2 внутри клеток оболочки пучка . Эти клетки являются частью характерной анатомии листа Кранца , которая пространственно разделяет типы фотосинтезирующих клеток в концентрическом расположении для накопления CO 2 рядом с RuBisCO. [ 21 ]
Эти химические и анатомические механизмы улучшают способность RuBisCO связывать углерод, а не осуществлять его бесполезную оксигеназную активность. Активность оксигеназы RuBisCO, называемая фотодыханием , приводит к потере субстрата RuBP в результате оксигенации и при этом потребляется энергия. Адаптации растений С4 дают преимущество перед путем С3, который теряет эффективность из-за фотодыхания. [ 22 ] Соотношение фотодыхания и фотосинтеза у растения варьируется в зависимости от условий окружающей среды, поскольку снижение концентрации CO 2 и повышение концентрации O 2 увеличивают эффективность фотодыхания. [ 20 ] Уровень CO 2 в атмосфере на Земле резко снизился между 32 и 25 миллионами лет назад. Это дало селективное преимущество эволюции пути C4, который может ограничивать скорость фотодыхания, несмотря на снижение содержания CO 2 в окружающей среде . [ 23 ] Сегодня растения C4 составляют примерно 5% растительной биомассы на Земле, но около 23% земной фиксации углерода. [ 24 ] [ 25 ] [ 26 ] Типы растений, которые используют фотосинтез C4, включают травы и экономически важные культуры, такие как кукуруза , сахарный тростник , просо и сорго . [ 22 ] [ 27 ]
Изотопное фракционирование различается между фиксацией углерода C4 и C3 из-за пространственного разделения в растениях C4 захвата CO 2 (в клетках мезофилла) и цикла Кальвина (в клетках оболочки пучка). В растениях C4 углерод превращается в бикарбонат , фиксируется в оксалоацетат с помощью фермента фосфоенолпируват (PEP) карбоксилазы , а затем превращается в малат . [ 4 ] Малат транспортируется из мезофилла в клетки оболочки пучка , непроницаемые для CO 2 . Внутренний CO 2 концентрируется в этих клетках, поскольку малат повторно окисляется, а затем декарбоксилируется обратно в CO 2 и пируват. Это позволяет RuBisCO осуществлять катализ при достаточно высоком уровне внутреннего CO 2 , чтобы избежать конкурирующей реакции фотодыхания. Значение дельта в пути C4 составляет от -12 до -16‰, обедненное 13 C за счет совместного действия PEP-карбоксилазы и RuBisCO.
Изотопная дискриминация в пути C4 варьируется по сравнению с путем C3 из-за дополнительных стадий химического преобразования и активности PEP-карбоксилазы. После диффузии в устьица превращение CO 2 в бикарбонат концентрирует более тяжелые 13 C. Таким образом, последующая фиксация с помощью PEP-карбоксилазы менее истощается. 13 C, чем у Rubisco: содержание PEP-карбоксилазы обеднено примерно на 2 ‰ по сравнению с 29 ‰ в RuBisCO. [ 1 ] [ 5 ] Однако часть изотопно-тяжелого углерода, фиксируемого PEP-карбоксилазой, вытекает из клеток оболочки пучка. Это ограничивает количество углерода, доступного RuBisCO, что, в свою очередь, снижает эффект его фракционирования. [ 4 ] Это объясняет, что общее значение дельты на растениях C4 составляет от -12 до -16 ‰. [ 4 ]
На заводах CAM
[ редактировать ]Растения, которые используют метаболизм крассулацовых кислот , также известный как фотосинтез CAM, временно разделяют свои химические реакции между днем и ночью. Эта стратегия модулирует устьичную проводимость для повышения эффективности использования воды, поэтому она хорошо адаптирована к засушливому климату. [ 28 ] Ночью CAM-растения открывают устьица, чтобы позволить CO 2 проникнуть в клетку и фиксироваться в органических кислотах, которые хранятся в вакуолях. Этот углерод высвобождается в цикл Кальвина в течение дня, когда устьица закрыты, чтобы предотвратить потерю воды, а световые реакции могут стимулировать необходимое производство АТФ и НАДФН. [ 29 ] Этот путь отличается от фотосинтеза C4, поскольку растения CAM выделяют углерод, сохраняя фиксированный CO 2 в везикулах ночью, а затем транспортируя его для использования в течение дня. Таким образом, растения CAM временно концентрируют CO 2 для повышения эффективности RuBisCO, тогда как растения C4 пространственно концентрируют CO 2 в клетках оболочки пучка. Распространение растений, использующих фотосинтез САМ, включает эпифиты (например, орхидеи , бромелии ) и ксерофиты (например, суккуленты , кактусы ). [ 30 ]
В метаболизме крассуловой кислоты изотопное фракционирование сочетает в себе эффекты пути C3 в дневное время и пути C4 в ночное время. Ночью, когда температура и потеря воды ниже, CO 2 диффундирует через устьица и производит малат под действием фосфенолпируваткарбоксилазы. [ 4 ] [ 6 ] В течение следующих суток устьица закрываются, малат декарбоксилируется и CO 2 фиксируется RuBisCO. Сам по себе этот процесс аналогичен процессу у растений C4 и дает характерные значения фракционирования C4 примерно -11 ‰. [ 6 ] Однако во второй половине дня CAM-растения могут открыть устьица и осуществить фотосинтез C3. [ 6 ] Только в дневное время растения САМ имеют фракционирование примерно -28‰, характерное для растений С3. [ 6 ] Эти комбинированные эффекты обеспечивают δ 13 Значения C для растений CAM находятся в диапазоне от -10 до -20‰.
The 13 С до 12 Соотношение C в растениях CAM может указывать на временное разделение фиксации CO 2 , которое представляет собой степень биомассы, полученной в результате ночной фиксации CO 2 по сравнению с дневной фиксацией CO 2 . [ 31 ] Такое различие можно провести потому, что PEP-карбоксилаза, фермент, ответственный за чистое поглощение CO2 в ночное время, различает 13 C меньше, чем RuBisCO, который отвечает за поглощение CO 2 в дневное время . Предполагается, что растения CAM, которые улавливают CO 2 преимущественно ночью, будут показывать δ 13 Значения C более похожи на значения C4, тогда как дневная CO 2 фиксация будет показывать δ. 13 Значения C больше похожи на растения C3.
В фитопланктоне
[ редактировать ]В отличие от наземных растений, у которых диффузия CO 2 в воздухе происходит относительно быстро и обычно не ограничивает, диффузия растворенного CO 2 в воде происходит значительно медленнее и часто может ограничивать фиксацию углерода в фитопланктоне. [ 5 ] Когда газообразный CO 2 (г) растворяется в водном CO 2 (водный раствор) , он фракционируется под действием как кинетических, так и равновесных эффектов , которые зависят от температуры. [ 32 ] По сравнению с растениями источник растворенного CO 2 для фитопланктона может быть обогащен 13 C примерно на 8‰ от атмосферного CO 2 . [ 33 ]
Изотопное фракционирование 13 C фотосинтеза фитопланктона зависит от диффузии внеклеточного водного CO 2 в клетку, RuBisCO-зависимой скорости роста клеток, а также геометрии клеток и площади поверхности. [ 7 ] Использование механизмов концентрации бикарбоната и углерода в фитопланктоне отличает изотопное фракционирование от путей фотосинтеза растений.
Разница между внутриклеточной и внеклеточной концентрацией CO 2 отражает потребность в CO 2 клетки фитопланктона, которая зависит от скорости ее роста. Отношение потребности в углероде к его предложению определяет диффузию CO 2 в клетку и отрицательно коррелирует с величиной фракционирования углерода фитопланктоном. [ 34 ] фракционирование между CO 2 (водн.) и биомассой фитопланктона для оценки скорости роста фитопланктона. В совокупности эти соотношения позволяют использовать [ 35 ]
Однако сама по себе скорость роста не объясняет наблюдаемое фракционирование. Поток CO 2 (водн.) в клетку и из нее примерно пропорционален площади поверхности клетки, а углеродная биомасса клетки варьируется в зависимости от объема клетки. Геометрия фитопланктона, которая максимизирует площадь поверхности по отношению к объему, должна иметь большее фракционирование изотопов в результате фотосинтеза. [ 36 ]
Биохимические характеристики фитопланктона аналогичны растениям С3, тогда как характеристики газообмена больше напоминают стратегию С4. [ 37 ] Точнее, фитопланктон повышает эффективность своего основного фермента, связывающего углерод, RuBisCO, с помощью механизмов концентрации углерода (CCM), точно так же, как растения C4 накапливают CO 2 в клетках оболочки пучка. Различные формы ККМ в фитопланктоне включают активное поглощение бикарбоната и СО 2 через клеточную мембрану, активный транспорт неорганического углерода от клеточной мембраны к хлоропластам и активное однонаправленное превращение СО 2 в бикарбонат. [ 38 ] Параметры, влияющие 13 Фракционирование C в фитопланктоне способствует δ 13 Значения C от -18 до -25‰. [ 4 ] [ 7 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час Г.Д. Фаркуар; Дж. Р. Элерингер; Хабик и КТ (1989). «Дискриминация изотопов углерода и фотосинтез». Ежегодный обзор физиологии растений и молекулярной биологии растений . 40 (1): 503–537. дои : 10.1146/annurev.pp.40.060189.002443 .
- ^ Перейти обратно: а б с Кон, Мэтью Дж. (16 ноября 2010 г.). «Изотопный состав углерода наземных растений C3 как индикаторы (палео)экологии и (палео)климата» . Труды Национальной академии наук . 107 (46): 19691–19695. Бибкод : 2010PNAS..10719691K . дои : 10.1073/pnas.1004933107 . ISSN 0027-8424 . ПМЦ 2993332 . ПМИД 21041671 .
- ^ Фрай, Б.; Шерр, Э.Б. (1989). «Измерения δ13C как индикаторы потока углерода в морских и пресноводных экосистемах». Стабильные изотопы в экологических исследованиях . Экологические исследования. Том. 68. Нью-Йорк, штат Нью-Йорк: Springer New York. стр. 196–229. дои : 10.1007/978-1-4612-3498-2_12 . ISBN 9781461281276 .
- ^ Перейти обратно: а б с д и ж г час Хейс, Джон (1 января 2001 г.). «Фракционирование изотопов углерода и водорода в процессах биосинтеза» . Обзоры по минералогии и геохимии . 43 (1): 225–277. Бибкод : 2001RvMG...43..225H . дои : 10.2138/gsrmg.43.1.225 .
- ^ Перейти обратно: а б с д и ж г О'Лири, Мэрион Х. (май 1988 г.). «Изотопы углерода в фотосинтезе». Бионаука . 38 (5): 328–336. дои : 10.2307/1310735 . ISSN 0006-3568 . JSTOR 1310735 .
- ^ Перейти обратно: а б с д и О'Лири, Мэрион Х. (1988). «Изотопы углерода в фотосинтезе». Бионаука . 38 (5): 328–336. дои : 10.2307/1310735 . JSTOR 1310735 .
- ^ Перейти обратно: а б с Попп, Брайан Н.; Лоус, Эдвард А.; Бидигар, Роберт Р.; Дор, Джон Э.; Хэнсон, Кристи Л.; Уэйкхэм, Стюарт Г. (январь 1998 г.). «Влияние геометрии клеток фитопланктона на изотопное фракционирование углерода». Geochimica et Cosmochimica Acta . 62 (1): 69–77. Бибкод : 1998GeCoA..62...69P . дои : 10.1016/S0016-7037(97)00333-5 . ISSN 0016-7037 .
- ^ Перейти обратно: а б Гонфиантини, Роберто (апрель 1984 г.). «Совещание консультативной группы МАГАТЭ по эталонным образцам стабильных изотопов для геохимических и гидрологических исследований». Химическая геология . 46 (1): 85. Бибкод : 1984ЧГео..46...85Г . дои : 10.1016/0009-2541(84)90167-0 . ISSN 0009-2541 .
- ^ Маккоун, Гарольд Т. (сентябрь 1992 г.). «Введение в морскую биогеохимию (Либес, Сьюзен М.)» . Журнал химического образования . 69 (9): А251. Бибкод : 1992ЖЧЭд..69..251М . дои : 10.1021/ed069pa251.2 . ISSN 0021-9584 .
- ^ МакКлинток, Барбара М. (март 1977 г.). «Биологическая океанография». Американский учитель биологии . 39 (3): 186. дои : 10.2307/4445858 . hdl : 2027/umn.31951d01800724l . ISSN 0002-7685 . JSTOR 4445858 .
- ^ Перейти обратно: а б Х., Рэйвен, Питер (2005). Биология растений . Эверт, Рэй Франклин, Эйххорн, Сьюзен Э. (7-е изд.). WH Freeman and Co. Нью-Йорк: ISBN 978-0716710073 . OCLC 56051064 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Табита, Франция; Сатагопан, С.; Хэнсон, TE; Крил, штат Невада; Скотт, СС (19 июня 2007 г.). «Отличные формы белков Рубиско I, II, III и IV из трех царств жизни дают ключ к разгадке эволюции Рубиско и взаимоотношений структуры и функции» . Журнал экспериментальной ботаники . 59 (7): 1515–1524. дои : 10.1093/jxb/erm361 . ISSN 0022-0957 . ПМИД 18281717 .
- ^ Скотт, Кэтлин М.; Шведок, Джули; Шраг, Дэниел П.; Кавано, Коллин М. (декабрь 2004 г.). «Влияние формы IA RubisCO и растворенного в окружающей среде неорганического углерода на delta13C симбиоза моллюска и хемоавтотрофа Solemya velum». Экологическая микробиология . 6 (12): 1210–1219. дои : 10.1111/j.1462-2920.2004.00642.x . ISSN 1462-2912 . ПМИД 15560819 .
- ^ Гай, доктор медицинских наук; Фогель, МЛ; Берри, Дж. А. (1 января 1993 г.). «Фотосинтетическое фракционирование стабильных изотопов кислорода и углерода» . Физиология растений . 101 (1): 37–47. дои : 10.1104/стр.101.1.37 . ISSN 0032-0889 . ПМК 158645 . ПМИД 12231663 .
- ^ Макневин, Деннис Б.; Бэджер, Мюррей Р.; Уитни, Спенсер М.; Кеммерер, Сюзанна фон; Черкез, Гийом ГБ; Фаркуар, Грэм Д. (7 декабря 2007 г.). «Различия в дискриминации изотопов углерода трех вариантов D-рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы отражают различия в их каталитических механизмах» . Журнал биологической химии . 282 (49): 36068–36076. дои : 10.1074/jbc.M706274200 . ISSN 0021-9258 . ПМИД 17925403 .
- ^ Перейти обратно: а б Фаркуар, Грэм; О'Лири, Миннесота; Берри, Джозеф (1 января 1982 г.). «О связи между дискриминацией изотопов углерода и межклеточной концентрацией углекислого газа в листьях» . Австралийский журнал физиологии растений . 13 (2): 281–292. дои : 10.1071/PP9820121 .
- ^ Шуберт, Брайан А.; Джарен, А. Хоуп (ноябрь 2012 г.). «Влияние концентрации CO2 в атмосфере на фракционирование изотопов углерода в наземных растениях C3» . Geochimica et Cosmochimica Acta . 96 : 29–43. Бибкод : 2012GeCoA..96...29S . дои : 10.1016/j.gca.2012.08.003 . ISSN 0016-7037 .
- ^ Уайтхед, Марк (6 марта 2017 г.). «Окружающая среда и государство». Международная энциклопедия географии . Оксфорд, Великобритания: John Wiley & Sons, Ltd., стр. 1–11. дои : 10.1002/9781118786352.wbieg0920 . ISBN 9780470659632 .
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Перейти обратно: а б Морено-Гутьеррес, Кристина; Доусон, Тодд Э.; Николас, Эмили; Керехета, Хосе Игнатиус (23 августа 2012 г.). «Изотопы выявляют контрастирующие стратегии использования воды среди сосуществующих видов растений в экосистеме Средиземноморья». Новый фитолог . 196 (2): 489–496. дои : 10.1111/j.1469–8137.2012.04276.x . ISSN 0028-646X . ПМИД 22913668 .
- ^ Перейти обратно: а б Элерингер, Джеймс Р.; Сейдж, Роуэн Ф.; Фланаган, Лоуренс Б.; Пирси, Роберт В. (1 марта 1991 г.). «Изменение климата и эволюция фотосинтеза C4». Тенденции в экологии и эволюции . 6 (3): 95–99. дои : 10.1016/0169-5347(91)90183-X . ISSN 0169-5347 . ПМИД 21232434 .
- ^ Кеннеди, Роберт А. (23 апреля 1976 г.). «Фотодыхание в культурах тканей растений C3 и C4» . Физиология растений . 58 (4): 573–575. дои : 10.1104/стр.58.4.573 . ПМЦ 543284 . ПМИД 16659720 .
- ^ Перейти обратно: а б C4 биология растений . Сейдж, Роуэн Фредерик, Монсон, Р.К. (Рассел К.), 1954-. Сан-Диего: Академическая пресса. 1999. ISBN 9780080528397 . OCLC 176630229 .
{{cite book}}: CS1 maint: другие ( ссылка ) - ^ Сейдж, Роуэн Ф.; Сейдж, Тэмми Л.; Кокачинар, Ферит (2012). «Фотодыхание и эволюция фотосинтеза C4 | Ежегодный обзор биологии растений». Ежегодный обзор биологии растений . 63 (1): 19–47. doi : 10.1146/annurev-arplant-042811-105511 . ПМИД 22404472 . S2CID 24199852 .
- ^ Бонд, Вашингтон; Вудворд, Финляндия; Миджли, GF (12 ноября 2004 г.). «Глобальное распространение экосистем в мире без огня». Новый фитолог . 165 (2): 525–538. дои : 10.1111/j.1469-8137.2004.01252.x . ISSN 0028-646X . ПМИД 15720663 . S2CID 4954178 .
- ^ Осборн, CP; Бирлинг, диджей (29 января 2006 г.). «Зеленая революция природы: замечательный эволюционный рост растений C4» . Философские труды Королевского общества B: Биологические науки . 361 (1465): 173–194. дои : 10.1098/rstb.2005.1737 . ISSN 0962-8436 . ПМК 1626541 . ПМИД 16553316 .
- ^ Келлог, Элизабет А. (июль 2013 г.). «Фотосинтез С4» . Современная биология . 23 (14): 594–599 рэндов. дои : 10.1016/j.cub.2013.04.066 . ISSN 0960-9822 . ПМИД 23885869 .
- ^ Чжу, Синь-Гуан; Лонг, Стивен П; Орт, Дональд Р. (апрель 2008 г.). «Какова максимальная эффективность, с которой фотосинтез может преобразовать солнечную энергию в биомассу?». Современное мнение в области биотехнологии . 19 (2): 153–159. дои : 10.1016/j.copbio.2008.02.004 . ISSN 0958-1669 . ПМИД 18374559 .
- ^ Тинг, ИП (июнь 1985 г.). «Метаболизм крассуловой кислоты». Ежегодный обзор физиологии растений . 36 (1): 595–622. дои : 10.1146/annurev.pp.36.060185.003115 . HDL : 10150/552219 . ISSN 0066-4294 .
- ^ Тинг, И. (1 января 1985 г.). «Метаболизм крассуловой кислоты». Ежегодный обзор физиологии растений и молекулярной биологии растений . 36 (1): 595–622. дои : 10.1146/annurev.pp.36.060185.003115 . HDL : 10150/552219 . ISSN 1040-2519 .
- ^ Смит, JAC; Винтер, К. (1996). «Таксономическое распределение метаболизма крассуловых кислот». Метаболизм крассуловой кислоты . Экологические исследования. Том. 114. Берлин, Гейдельберг: Springer Berlin Heidelberg. стр. 427–436. дои : 10.1007/978-3-642-79060-7_27 . ISBN 9783642790621 .
- ^ Зима, Клаус; Холтум, Джозеф А.М. (1 августа 2002 г.). «Насколько точно значения δ13C растений с кислотным метаболизмом толстянковых отражают долю CO2, фиксируемую днем и ночью?» . Физиология растений . 129 (4): 1843–1851. дои : 10.1104/стр.002915 . ISSN 0032-0889 . ПМК 166772 . ПМИД 12177497 .
- ^ Чжан, Дж.; Набережная, Вашингтон; Уилбур, DO (1 января 1995 г.). «Фракционирование изотопов углерода при газоводяном обмене и растворении СО2». Geochimica et Cosmochimica Acta . 59 (1): 107–114. Бибкод : 1995GeCoA..59..107Z . дои : 10.1016/0016-7037(95)91550-D . ISSN 0016-7037 .
- ^ Кёлер, П.; Фишер, Х.; Шмитт, Дж. (март 2010 г.). «Атмосферное δ13CO2 и его соотношение с верхним CO2 и глубоким океаном δ13C в позднем плейстоцене» (PDF) . Палеоокеанография . 25 (1). Бибкод : 2010PalOc..25.1213K . дои : 10.1029/2008pa001703 . ISSN 0883-8305 .
- ^ Лоус, Эдвард А.; Попп, Брайан Н.; Кассар, Николя; Танимото, Джейми (2002). «Модели дискриминации 13C в океаническом фитопланктоне: вероятное влияние механизмов концентрации CO2 и последствия для палеореконструкций». Функциональная биология растений . 29 (3): 323–333. дои : 10.1071/pp01183 . ISSN 1445-4416 . ПМИД 32689479 .
- ^ Лоус, Эдвард А.; Попп, Брайан Н.; Бидигар, Роберт Р.; Кенникатт, Махлон К.; Мако, Стивен А. (1 марта 1995 г.). «Зависимость изотопного состава углерода фитопланктона от скорости роста и [CO2)вод: теоретические соображения и экспериментальные результаты». Geochimica et Cosmochimica Acta . 59 (6): 1131–1138. Бибкод : 1995GeCoA..59.1131L . дои : 10.1016/0016-7037(95)00030-4 . ISSN 0016-7037 .
- ^ Попп, Брайан Н.; Лоус, Эдвард А.; Бидигар, Роберт Р.; Дор, Джон Э.; Хэнсон, Кристи Л.; Уэйкхэм, Стюарт Г. (1 января 1998 г.). «Влияние геометрии клеток фитопланктона на изотопное фракционирование углерода». Geochimica et Cosmochimica Acta . 62 (1): 69–77. Бибкод : 1998GeCoA..62...69P . дои : 10.1016/S0016-7037(97)00333-5 . ISSN 0016-7037 .
- ^ Лоус, Эдвард А.; Бидигар, Роберт Р.; Попп, Брайан Н. (ноябрь 1997 г.). «Влияние скорости роста и концентрации CO2 на изотопное фракционирование углерода морской диатомовой водорослью Phaeodactylum tricornutum» . Лимнология и океанография . 42 (7): 1552–1560. Бибкод : 1997LimOc..42.1552L . дои : 10.4319/lo.1997.42.7.1552 . ISSN 0024-3590 .
- ^ Кассар, Николя; Лоус, Эдвард А.; Попп, Брайан Н. (ноябрь 2006 г.). «Изотопное фракционирование углерода морской диатомовой водорослью Phaeodactylum tricornutum в условиях роста с ограничением питательных веществ и света». Geochimica et Cosmochimica Acta . 70 (21): 5323–5335. Бибкод : 2006GeCoA..70.5323C . дои : 10.1016/j.gca.2006.08.024 . ISSN 0016-7037 .