Развитие завода
Важными структурами развития растений являются почки , побеги , корни , листья и цветки ; растения производят эти ткани и структуры на протяжении всей своей жизни из меристем. [ 1 ] располагаются на кончиках органов или между зрелыми тканями. Таким образом, живое растение всегда имеет эмбриональные ткани. Напротив, эмбрион животного очень рано производит все части тела, которые у него когда-либо будут в жизни. Когда животное рождается (или вылупляется из яйца), у него есть все части тела, и с этого момента оно становится только крупнее и взрослее. Однако и растения, и животные проходят филотипическую стадию , которая развивалась независимо. [ 2 ] и это вызывает ограничения развития, ограничивающие морфологическое разнообразие. [ 3 ] [ 4 ] [ 5 ] [ 6 ]
По мнению физиолога растений А. Карла Леопольда , свойства организации, наблюдаемые в растении, представляют собой возникающие свойства, которые представляют собой нечто большее, чем сумма отдельных частей. «Сборка этих тканей и функций в единый многоклеточный организм дает не только характеристики отдельных частей и процессов, но и совершенно новый набор характеристик, который невозможно было бы предсказать на основе изучения отдельных частей». [ 7 ]
Рост
[ редактировать ]начинается Сосудистое растение из одноклеточной зиготы , образующейся в результате оплодотворения яйцеклетки сперматозоидом. С этого момента оно начинает делиться, образуя зародыш растения в процессе эмбриогенеза . При этом образующиеся клетки организуются так, что один конец становится первым корнем, а другой конец образует верхушку побега. У семенных растений у зародыша разовьется один или несколько «семенных листьев» ( семядолей ). К концу эмбриогенеза молодое растение будет иметь все части, необходимые для начала его жизни. [ нужна ссылка ]
Как только эмбрион прорастает из семени или родительского растения, он начинает производить дополнительные органы (листья, стебли и корни) в процессе органогенеза . Новые корни вырастают из корневых меристем, расположенных на кончике корня, а новые стебли и листья вырастают из меристем побега , расположенных на кончике побега. [ 8 ] Ветвление происходит, когда небольшие скопления клеток, оставшиеся от меристемы и еще не подвергшиеся клеточной дифференцировке с образованием специализированной ткани, начинают расти как верхушка нового корня или побега. Рост любой такой меристемы на кончике корня или побега называется первичным ростом и приводит к удлинению этого корня или побега. Вторичный рост приводит к расширению корня или побега за счет деления клеток камбия . [ 9 ]
Помимо роста за счет деления клеток , растение может расти за счет удлинения клеток . Это происходит, когда отдельные клетки или группы клеток становятся длиннее. Не все растительные клетки вырастают до одинаковой длины. Когда клетки на одной стороне стебля растут длиннее и быстрее, чем клетки на другой стороне, в результате стебель изгибается в сторону более медленно растущих клеток. Этот направленный рост может происходить через реакцию растения на определенный раздражитель, такой как свет ( фототропизм ), гравитация ( гравитропизм ), вода ( гидротропизм ) и физический контакт ( тигмотропизм ). [ нужна ссылка ]
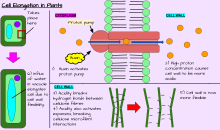

Рост и развитие растений опосредованы специфическими фитогормонами и регуляторами роста растений (РГР) (Росс и др., 1983). [ 10 ] На уровень эндогенных гормонов влияют возраст растения, холодостойкость, период покоя и другие метаболические условия; фотопериод, засуха, температура и другие внешние условия среды; и экзогенные источники ГРР, например, применяемые извне и ризосферного происхождения.
Морфологические изменения во время роста
[ редактировать ]Растения демонстрируют естественные различия в своей форме и строении. Хотя все организмы варьируются от человека к человеку, растения демонстрируют дополнительный тип вариаций. Внутри одной особи повторяются части, которые по форме и строению могут отличаться от других подобных частей. Эту вариацию легче всего увидеть на листьях растения, хотя и в других органах, таких как стебли и цветы, могут наблюдаться аналогичные вариации. Есть три основные причины этого изменения: позиционные эффекты, воздействие окружающей среды и молодость. [ нужна ссылка ]

Части взрослого растения различаются в зависимости от относительного положения, в котором образуется орган. Например, на новой ветке листья могут меняться по одинаковому рисунку вдоль ветки. Форма листьев, образующихся у основания ветки, отличается от листьев, образующихся на кончике растения, и это различие сохраняется от ветки к ветке на данном растении и у данного вида.
На то, как созревают новые структуры по мере их образования, может влиять момент в жизни растений, когда они начинают развиваться, а также окружающая среда, воздействию которой подвергаются структуры. Температура оказывает множественное воздействие на растения в зависимости от множества факторов, включая размер и состояние растения, а также температуру и продолжительность воздействия. Чем меньше и сочнее растение, тем выше вероятность повреждения или гибели от слишком высоких или слишком низких температур. Температура влияет на скорость биохимических и физиологических процессов, причем скорость обычно (в определенных пределах) увеличивается с температурой.
Ювенильность или гетобластия — это когда органы и ткани, вырабатываемые молодым растением, например, рассадой , часто отличаются от тех, которые производятся тем же растением в более старшем возрасте. Например, молодые деревья будут давать более длинные и тонкие ветви, которые будут расти вверх больше, чем ветви, которые они будут производить как взрослое дерево. Кроме того, листья, образующиеся на ранних стадиях роста, обычно крупнее, тоньше и неровнее, чем листья взрослого растения. Образцы молодых растений могут настолько сильно отличаться от взрослых растений того же вида, что насекомые, откладывающие яйца, не воспринимают растение как пищу для своих детенышей. Переход от ранних форм роста к поздним иногда называют вегетативной фазовой сменой . [ 11 ]
Адвентивные структуры
[ редактировать ]Структуры растений, в том числе корни, почки и побеги, развивающиеся в необычных местах, называются адвентивными . Такие структуры часто встречаются у сосудистых растений. [ нужна ссылка ]
Придаточные корни и почки обычно развиваются вблизи существующих сосудистых тканей, чтобы они могли соединяться с ксилемой и флоэмой . Однако точное местоположение сильно различается. У молодых стеблей из паренхимы между сосудистыми пучками часто образуются придаточные корни . У стеблей со вторичным ростом придаточные корни часто берут начало в паренхиме флоэмы вблизи сосудистого камбия . У стеблевых черенков придаточные корни иногда зарождаются и в каллусных клетках, образующихся на поверхности среза. Листовые черенки толстянки образуют в эпидермисе придаточные корни. [ 12 ]
Бутоны и побеги
[ редактировать ]Придаточные почки развиваются из других мест, кроме апикальной меристемы побега , которая возникает на кончике стебля или на узле побега , в пазухе листа, причем почка остается там во время первичного роста. Они могут развиваться на корнях или листьях или на побегах в виде новых побегов. Апикальные меристемы побега дают одну или несколько пазушных или боковых почек в каждом узле. Когда стебли дают значительный вторичный прирост , пазушные почки могут быть разрушены. Затем на стеблях со вторичным ростом могут развиться придаточные почки. [ нужна ссылка ]
стебля Придаточные почки часто образуются после повреждения или обрезки . Придаточные почки помогают заменить потерянные ветки. Придаточные почки и побеги также могут развиваться на зрелых стволах деревьев, когда затененный ствол подвергается воздействию яркого солнечного света из-за вырубки окружающих деревьев. На деревьях секвойи вечнозеленой ( Sequoia sempervirens ) на нижних стволах часто образуется множество придаточных почек. Если главный ствол отмирает, из одной из придаточных почек часто прорастает новый. Небольшие кусочки ствола красного дерева продаются в качестве сувениров, называемых капами красного дерева. Их помещают в поддон с водой, и придаточные почки прорастают, образуя побеги. [ нужна ссылка ]
У некоторых растений на корнях обычно образуются придаточные почки, которые могут простираться на довольно большое расстояние от растения. Побеги, развивающиеся из придаточных почек на корнях, называются присосками . Они представляют собой тип естественного вегетативного размножения многих видов , например, многих трав, осины трясущейся и чертополоха канадского . Осина Пандо дрожащая выросла с одного ствола до 47 000 стволов за счет образования придаточных почек на одной корневой системе. [ нужна ссылка ]
На некоторых листьях образуются придаточные почки, которые затем образуют придаточные корни в рамках вегетативного размножения ; например, комбинированное растение ( Tolmiea menziesii ) и мать тысяч ( Kalanchoe daigremontiana ). Придаточные ростки затем опадают с материнского растения и развиваются как отдельные клоны родительского растения. [ нужна ссылка ]
Коппинг — это практика срезания стволов деревьев до земли, чтобы способствовать быстрому росту дополнительных побегов. Традиционно его используют для производства столбов, материала для заборов или дров. Это также практикуется для культур биомассы , выращиваемых для получения топлива, таких как тополь или ива.
Корни
[ редактировать ]
Для некоторых видов адвентивное укоренение может быть акклиматизацией, позволяющей избежать стресса, вызванной такими факторами, как гипоксия. [ 13 ] или дефицит питательных веществ. Другой экологически важной функцией придаточного укоренения является вегетативное размножение таких древесных пород, как Salix и Sequoia, в прибрежных районах. [ 14 ]
Способность стеблей растений образовывать придаточные корни используется при коммерческом размножении черенками . Понимание физиологических механизмов адвентивного укоренения позволило добиться определенного прогресса в улучшении укоренения черенков за счет применения синтетических ауксинов в качестве укореняющих порошков и использования селективного базального ранения. [ 15 ] Дальнейший прогресс может быть достигнут в будущие годы путем применения исследований других механизмов регулирования коммерческого размножения и путем сравнительного анализа молекулярного и экофизиологического контроля случайного укоренения у «трудно укореняющихся» и «легко укореняющихся» видов. [ нужна ссылка ]
Придаточные корни и почки очень важны при размножении растений черенками, отводками , тканевой культурой . Растительные гормоны , называемые ауксинами , часто наносятся на черенки стеблей, побегов или листьев, чтобы способствовать образованию придаточных корней, например, листья африканской фиалки и очитка, а также побеги пуансеттии и колеуса . Размножение корневыми черенками требует образования придаточных почек, как, например, у хрена и яблони . При отводках на надземных стеблях образуются придаточные корни до того, как часть стебля удаляется и образуется новое растение. Крупные комнатные растения часто размножают воздушными отводками . При размножении растений тканевой культурой должны развиваться придаточные корни и почки.
Модифицированные формы
[ редактировать ]- Клубневые корни лишены определенной формы; пример: сладкий картофель .
- Фасцикулированный корень (клубневидный корень) встречается гроздьями у основания стебля; примеры: спаржа , георгин .
- Узловатые корни возле кончиков опухают; пример: куркума .
- Корни ходулей возникают из первых нескольких узлов стебля. Они проникают в почву под углом и оказывают поддержку растению; примеры: кукуруза , сахарный тростник .
- Опорные корни оказывают механическую поддержку воздушным ветвям. Боковые ветви растут вертикально вниз в почву и действуют как опоры; пример: баньян .
- Лазящие корни, возникающие из узлов, прикрепляются к какой-либо опоре и перелезают через нее; пример: Эпипремнум золотистый .
- Монилиформные или бисерные корни, мясистые корни придают вид бусинок, например: горькая тыква , портулак .
Развитие листьев
[ редактировать ]Генетика, лежащая в основе развития формы листьев Arabidopsis thaliana, разбита на три стадии: зарождение зачатка листа , установление дорсивентральной структуры и развитие маргинальной меристемы . Зачаток листа инициируется подавлением генов и белков семейства KNOX класса I (таких как SHOOT APICAL MERISTEMLESS ). Эти белки KNOX класса I непосредственно подавляют биосинтез гиббереллина в примодии листа. Было обнаружено, что многие генетические факторы участвуют в подавлении этих генов в зачатках листьев (таких как ASYMMETRIC LEAVES1, BLADE-ON-PETIOLE1 , SAWTOOTH1 и т. д.). Таким образом, при таком подавлении уровни гиббереллина повышаются, и приморий листьев инициирует рост. [ нужна ссылка ]
Развитие цветов
[ редактировать ]
Развитие цветка — это процесс, посредством которого покрытосеменные растения создают образец экспрессии генов в меристемах , который приводит к появлению органа, ориентированного на половое размножение , — цветка. три физиологических Для того чтобы это произошло, должны произойти развития: во-первых, растение должно перейти от половой незрелости к половозрелому состоянию (т. е. перейти к цветению); во-вторых, трансформация функции апикальной меристемы из вегетативной меристемы во цветочную меристему или соцветие ; и, наконец, рост отдельных органов цветка. Последняя фаза была смоделирована с использованием модели ABC , которая описывает биологическую основу процесса с точки зрения молекулярной генетики и генетики развития . [ нужна ссылка ]

необходим внешний стимул Чтобы вызвать дифференцировку меристемы в меристему цветка, . Этот стимул активирует митотическое деление клеток в меристеме, особенно на ее сторонах, где новые зачатки образуются . Этот же стимул также заставит меристему следовать модели развития , которая приведет к росту цветочных меристем в отличие от вегетативных меристем. Основным отличием этих двух типов меристемы, помимо очевидного различия объективного органа, является мутовчатый (или мутовчатый) филлотаксис , т. е. отсутствие стеблевого удлинения среди последовательных мутовок или . мутовок зачатка Эти мутовки развиваются акропетально, давая начало чашелистикам , лепесткам , тычинкам и плодолистикам . Еще одно отличие от вегетативных пазушных меристем состоит в том, что цветочная меристема является «детерминированной», что означает, что после дифференцировки ее клетки больше не будут делиться . [ 16 ]
Идентичность органов, присутствующих в четырех цветочных мутовках, является следствием взаимодействия по крайней мере трех типов генных продуктов , каждый из которых имеет различные функции. Согласно модели ABC, функции A и C необходимы для определения идентичности мутовок околоцветника и репродуктивных мутовок соответственно. Эти функции эксклюзивны, и отсутствие одной из них означает, что другая будет определять идентичность всех цветочных мутовок. Функция B позволяет отличить лепестки от чашелистиков на вторичной мутовке, а также дифференцировать тычинку от плодолистика на третичной мутовке.
Цветочный аромат
[ редактировать ]Растения используют цветочную форму, цветы и аромат, чтобы привлечь различных насекомых для опыления . Определенные соединения в издаваемом аромате привлекают определенных опылителей . В петунии гибридной выделяются летучие бензоиды, придающие цветочный запах. Хотя компоненты пути биосинтеза бензоидов известны, ферменты этого пути и последующая регуляция этих ферментов еще предстоит открыть. [ 17 ]
Чтобы определить регуляцию пути, цветы P. Hybrida , специфичном для лепестков, Mitchell были использованы в микрочипе чтобы сравнить цветы, которые вот-вот начнут издавать аромат, с цветками P. Hybrida сорта W138, которые производят мало летучих бензоидов. кДНК генов обоих растений секвенировали. Результаты показали, что в цветках Mitchell присутствует повышенный уровень транскрипционного фактора, но нет в цветках W138, лишенных цветочного аромата. Этот ген получил название ODORANT1 (ODO1). Для определения экспрессии ODO1 в течение дня гель-блоттинг РНК проводили . Гель показал, что уровни транскрипта ODO1 начали увеличиваться между 13:00 и 16:00 часами, достигли максимума в 22:00 и были самыми низкими в 10:00. Эти уровни транскрипта ODO1 напрямую соответствуют временной шкале эмиссии летучих бензоидов. Кроме того, гель подтвердил предыдущий вывод о том, что неароматные цветки W138 имеют лишь одну десятую уровень транскрипта ODO1 цветков Mitchell. Таким образом, количество образующегося ODO1 соответствует количеству выделяемых летучих бензоидов, что указывает на то, что ODO1 регулирует биосинтез бензооидов. [ 17 ]
Дополнительными генами, участвующими в биосинтезе основных ароматических соединений, являются OOMT1 и OOMT2. OOMT1 и OOMT2 помогают синтезировать орцинол-О-метилтрансферазы (OOMT), которые катализируют последние два этапа пути ДМТ, создавая 3,5-диметокситолуол (ДМТ). ДМТ — это ароматическое соединение, вырабатываемое многими различными розами, однако некоторые сорта роз, такие как Rosa Gallica и Дамасская роза Rosa damascene , не выделяют ДМТ. Было высказано предположение, что эти сорта не производят ДМТ, потому что у них нет генов OOMT. Однако после эксперимента по иммунолокализации OOMT был обнаружен в эпидермисе лепестков. Для дальнейшего изучения этого явления лепестки роз подвергли ультрацентрифугированию . Супернатанты и осадки проверяли вестерн-блоттингом . Обнаружение белка OOMT при 150 000 г в супернатанте и осадке позволило исследователям заключить, что белок OOMT тесно связан с мембранами эпидермиса лепестков. Такие эксперименты показали, что гены OOMT действительно существуют у Rosa Gallica и дамасской розы Rosa damascene. сортов, но гены OOMT не экспрессируются в тканях цветка, где вырабатывается ДМТ. [ 18 ]
Ссылки
[ редактировать ]- ^ Бёрле, я; Ло, Т (2003). «Апикальные меристемы: источник молодости растения». Биоэссе . 25 (10): 961–70. дои : 10.1002/bies.10341 . ПМИД 14505363 . Обзор.
- ^ Дрост, Хайк-Георг; Яница, Филипп; Гросс, Иво; Квинт, Марсель (2017). «Сравнение песочных часов развития между королевствами» . Текущее мнение в области генетики и развития . 45 : 69–75. дои : 10.1016/j.где.2017.03.003 . ПМИД 28347942 .
- ^ Ирие, Наоки; Куратани, Сигэру (22 марта 2011 г.). «Сравнительный анализ транскриптома выявляет филотипический период позвоночных в ходе органогенеза» . Природные коммуникации . 2 : 248. Бибкод : 2011NatCo...2..248I . дои : 10.1038/ncomms1248 . ISSN 2041-1723 . ПМК 3109953 . ПМИД 21427719 .
- ^ Домазет-Лошо, Томислав; Таутц, Дитхард (9 декабря 2010 г.). «Филогенетически основанный индекс возраста транскриптома отражает закономерности онтогенетических расхождений». Природа . 468 (7325): 815–818. Бибкод : 2010Natur.468..815D . дои : 10.1038/nature09632 . ISSN 0028-0836 . ПМИД 21150997 . S2CID 1417664 .
- ^ Квинт, Марсель; Дрост, Хайк-Георг; Габель, Александр; Ульрих, Кристиан Карстен; Бонн, Маркус; Гроссе, Иво (4 октября 2012 г.). «Транскриптомные песочные часы в эмбриогенезе растений». Природа . 490 (7418): 98-101. Стартовый код : 2012Nature.490...98Q . дои : 10.1038/nature11394 . ISSN 0028-0836 . ПМИД 22951968 . S2CID 4404460 .
- ^ Дрост, Хайк-Георг; Габель, Александр; Гросс, Иво; Квинт, Марсель (01 мая 2015 г.). «Доказательства активного поддержания филотранскриптомных паттернов песочных часов в эмбриогенезе животных и растений» . Молекулярная биология и эволюция . 32 (5): 1221–1231. дои : 10.1093/molbev/msv012 . ISSN 0737-4038 . ПМК 4408408 . ПМИД 25631928 .
- ^ Леопольд, А. Карл (1964). животное и там молодой . МакГроу-Хилл . п. 183.
- ^ Брэнд, У; Хобе, М; Саймон, Р. (2001). «Функциональные домены в меристемах побегов растений». Биоэссе . 23 (2): 134–41. doi : 10.1002/1521-1878(200102)23:2<134::AID-BIES1020>3.0.CO;2-3 . ПМИД 11169586 . S2CID 5833219 . Обзор.
- ^ Барлоу, П. (2005). «Определение узорчатых клеток в растительной ткани: вторичная флоэма деревьев». Биоэссе . 27 (5): 533–41. дои : 10.1002/bies.20214 . ПМИД 15832381 .
- ^ Росс, SD; Фарис, РП; Биндер, В.Д. 1983. Регуляторы роста и хвойные деревья: их физиология и потенциальное использование в лесном хозяйстве. п. 35–78 в Никелле, Л.Г. (ред.), Химические вещества, регулирующие рост растений. Том. 2, CRC Press, Бока-Ратон, Флорида.
- ^ Джонс, Синтия С. (1 ноября 1999 г.). «Очерк молодости, фазового перехода и гетеробластии семенных растений». Международный журнал наук о растениях . 160 (С6): 105–С111. дои : 10.1086/314215 . ISSN 1058-5893 . ПМИД 10572025 . S2CID 21757481 .
- ^ Маквей, И. 1938. Регенерация Crassula multicava . Американский журнал ботаники 25: 7-11. [1]
- ^ Дрю, MC; Джексон, МБ; Гиффард, С. (1979). «Адвентивное укоренение, способствующее этилену, и развитие кортикальных воздушных пространств (аэренхимы) в корнях могут быть адаптивной реакцией на затопление у Zea mays L» . Планта . 147 (1): 83–88. дои : 10.1007/BF00384595 . ПМИД 24310899 . S2CID 7232582 .
- ^ Найман, Роберт Дж.; Декан, Анри (1997). «Экология интерфейсов: прибрежные зоны» . Ежегодный обзор экологии и систематики . 28 : 621–658. doi : 10.1146/annurev.ecolsys.28.1.621 . JSTOR 2952507 . S2CID 86570563 .
- ^ Клерк Герт-Ян; Ван дер Крикен, Вим; Де Йонг, Шутка К. (1999). «Обзор формирования адвентивных корней: новые концепции, новые возможности» . Клеточная биология и биология развития in vitro — растения . 35 (3): 189–199. дои : 10.1007/s11627-999-0076-z . S2CID 44027145 .
- ^ Аскон-Бьето; и др. (2000). Основы физиологии растений . МакГроу-Хилл/Межамериканская Испания, ЮАР. ISBN 84-486-0258-7 . [ нужна страница ]
- ^ Перейти обратно: а б Шууринк, Роберт С.; Харинг, Мишель А.; Кларк, Дэвид Г. (2006). «Регуляция биосинтеза летучих бензоидов в цветках петунии». Тенденции в науке о растениях . 11 (1): 20–25. doi : 10.1016/j.tplants.2005.09.009 .
- ^ Скаллиет, Габриэль; и др. (01.01.2006). «Роль лепесткоспецифичных орцинол-О-метилтрансфераз в эволюции аромата розы» . Физиология растений . 140 (1): 18–29. дои : 10.1104/стр.105.070961 . ISSN 1532-2548 . ПМК 1326028 . ПМИД 16361520 .