Слуховой пузырек
| Слуховой пузырек | |
|---|---|
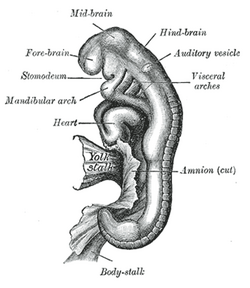 Эмбрион от восемнадцати до двадцати одного дня | |
 Общее формирование слухового пузырька | |
| Подробности | |
| Предшественник | Отическая плакода, слуховая ямка или отическая ямка, отическая чашка, |
| Дает начало | Перепончатый лабиринт внутреннего уха |
| Идентификаторы | |
| латинский | слуховой пузырек |
| ТО | vesicle_by_E5.15.1.0.0.0.4 E5.15.1.0.0.0.4 |
| Анатомическая терминология | |
Слуховой пузырек , или слуховой пузырек , состоит из двух мешкообразных впячиваний, образующихся и впоследствии закрывающихся во время эмбрионального развития. Это часть нервной эктодермы , которая разовьется в перепончатый лабиринт внутреннего уха. Этот лабиринт представляет собой сплошной эпителий , дающий начало вестибулярному аппарату и слуховым компонентам внутреннего уха. [1] На более ранних стадиях эмбриогенеза отическая плакода инвагинирует, образуя слуховую чашу. После этого слуховая чаша закрывается, образуя слуховой пузырек. После формирования слуховой пузырек будет располагаться рядом с нервной трубкой медиально, а на латеральной стороне будет параксиальная мезодерма . Клетки нервного гребня мигрируют рострально и каудально к плакоде.
Общая последовательность формирования слухового пузырька относительно консервативна у позвоночных , хотя существуют большие различия во времени и стадиях. [2] Формирование паттерна во время морфогенеза в характерных структурах внутреннего уха определяется гомеобокса транскрипционными факторами , включая PAX2 , DLX5 и DLX6 , причем первый специфицирует слуховые структуры, происходящие из вентральных ушных пузырьков, а два последних специфицируют дорсальные вестибулярные структуры. [ нужна ссылка ]
Разработка
[ редактировать ]Слуховой пузырек происходит из краниальной плакоды. [3] Ранний ушной пузырек характеризуется широкой компетентностью и может быть подразделен на сенсорный, несенсорный и нейрогенный компоненты. Сенсорные эпителиальные клетки и нейроны происходят из пронейросенсорного домена. Этот домен можно далее подразделить на нейрогенный субдомен и просенсорный субдомен. Просенсорный субдомен в конечном итоге дает начало опорным клеткам и волосковым клеткам, тогда как нейрогенный субдомен дает начало слуховому нейрону и вестибулярному нейрону.
Средняя часть слухового пузырька развивается в проток и эндолимфатический мешок . [4] Передний конец слухового пузырька постепенно удлиняется в виде трубки и закручивается, образуя начало улиткового протока . Вестибулярная конечность впоследствии сужается, образуя canalis reuniens . Центральная часть слухового пузырька представляет собой перепончатое преддверие и подразделяется перетяжкой на меньшую вентральную часть — мешочек и более крупную дорсальную и заднюю часть — утрикулу . Дорсальный компонент внутреннего уха также состоит из полукружных каналов . Маточка и мешочек сообщаются друг с другом посредством Y-образного канала.
Передача сигналов генов
[ редактировать ]Гены FGF , Bmp , Wnt и Pax, вероятно, участвуют в отической индукции. [5] Сигналы FGF и BMP помогают контролировать формирование паттерна в ранних ушных пузырьках. Предполагается, что Fgf3 и Fgf10 играют роль в индукции слуховых пузырьков у мышей, а гены Msx, как предполагается, играют роль в формировании слуховых пузырьков у кур. Pax8 экспрессируется на протяжении всего формирования ушных пузырьков. Другие гены, обнаруженные в слуховых пузырьках у разных видов и которые могут играть роль в формировании паттерна, включают гены Hmx, Fox, Dlx и Gbx.
Другие животные
[ редактировать ]Формирование слухового пузырька широко изучалось на модельных организмах развития, включая курицу , Xenopus , рыбок данио , аксолотля и мышь . [6] Переход от отической плакоды к отическому пузырьку происходит на стадии 19-го сомита у рыбок данио, Xenopus и кур. У птенца инвагинация слуховой плакоды происходит пассивно за счет движений окружающей плакоды. С другой стороны, отическая плакода у рыбок данио возникает в результате кавитации; эктодермальная плакода конденсируется и образует яйцевидный шар непосредственно под поверхностью эмбриона. Формирование слуховых пузырьков происходит позже, на стадии 25-30 сомитов у мышей.
Дополнительные изображения
[ редактировать ]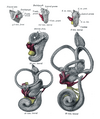 Боковые виды перепончатого лабиринта и акустического комплекса. Х 25 диам.
Боковые виды перепончатого лабиринта и акустического комплекса. Х 25 диам. Срединные виды перепончатого лабиринта и акустического комплекса эмбрионов человека. Х 25 диам.
Срединные виды перепончатого лабиринта и акустического комплекса эмбрионов человека. Х 25 диам.


Ссылки
[ редактировать ]- ^ Фрейер Л., Аггарвал В., Морроу Б.Е. (декабрь 2011 г.). «Двойное эмбриональное происхождение ушного пузырька млекопитающих, образующего внутреннее ухо» . Разработка . 138 (24): 5403–14. дои : 10.1242/dev.069849 . ПМЦ 3222214 . ПМИД 22110056 .
- ^ Парк BY, Сен-Жанне JP (декабрь 2008 г.). «Происходящие из заднего мозга сигналы Wnt и Fgf взаимодействуют, чтобы специфицировать отическую плакоду у Xenopus» . Биология развития . 324 (1): 108–21. дои : 10.1016/j.ydbio.2008.09.009 . ПМК 2605947 . ПМИД 18831968 .
- ^ Эплер Дж. М., Гудрич Л. В. (апрель 2011 г.). «Соединение уха с мозгом: Молекулярные механизмы сборки слухового контура» . Прогресс нейробиологии . 93 (4): 488–508. doi : 10.1016/j.pneurobio.2011.01.004 . ПМК 3078955 . ПМИД 21232575 .
- ^ Бриганда СП, Кирнан А.Е., Гао X, Итен Л.Е., Фекете Д.М. (октябрь 2000 г.). «Молекулярная генетика формирования структур внутреннего уха: играют ли роль границы отсеков?» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (22): 11700–6. Бибкод : 2000PNAS...9711700B . дои : 10.1073/pnas.97.22.11700 . ПМК 34338 . ПМИД 11050198 .
- ^ Чаттерджи С., Краус П., Луфкин Т. (июль 2010 г.). «Симфония генов, контролирующих развитие внутреннего уха» . БМК Генетика . 11 (1): 68. дои : 10.1186/1471-2156-11-68 . ПМК 2915946 . ПМИД 20637105 .
- ^ Норамли С., Грейнджер Р.М. (ноябрь 2002 г.). «Определение эмбрионального внутреннего уха» . Журнал нейробиологии . 53 (2): 100–28. дои : 10.1002/neu.10131 . ПМИД 12382270 .