Catoptbate
| Catoptbate Временный диапазон: поздний меховой ,
| |
|---|---|

| |
| Наиболее полный скелет (образец PM120/107), показанный сверху (слева, а) и ниже (справа, б) в соответствии с консервациями, с диаграммами, показывающими отдельные кости | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Млекопитающие |
| Заказ: | † Многотуберкулата |
| Семья: | † djadochtatheriidae |
| Род: | † Catopsbaatar Kielan-Jaworowska , 1994 |
| Разновидность: | † C. Catopsaloides
|
| Биномиальное название | |
| † Catopsbaatar Catopsaloides (Kielan-Jaworowska, 1974)
| |
| Синонимы | |
| |
Catopsbaatar -это род Multituberculate , вымерший порядок мимошних млекопитающих . Он жил в том, что сейчас является Монголией в позднем кампанианском возрасте поздней меловой эпохи , около 72 миллионов лет назад. Первые окаменелости были собраны в начале 1970 -х годов, и животное было названо новым видом рода Djadochtatherium в 1974 году D. catopsaloides . Конкретное название относится к сходству животного с родом Catopsalis . Этот вид был перенесен на род Catopsalis в 1979 году и получил свой собственный род ( Catopsbaatar , греческий и монгольский для «видимого героя») в 1994 году. Известны пять черепов, один моляр и один скелет с черепом; Последний - самый полный образец рода. Catopsbaatar был членом семьи Djadochtatheriidae .
Череп Catopsbaatar был длиной до 70 мм (2,8 дюйма) и, как и в других многотуркуляциях, пропорционально больших. Внешний вид голов этих животных, возможно, был похож на грызуны. Череп был тяжелым и широким, с зигоматическими арками, сильно расширенными по бокам. Глазки были меньше и были помещены назад, чем у его родственников, и морда была более удлиненной. Catopsbaatar имели полукруглые хребты на стороне черепа, к которым были прикреплены мышцы челюсти. Манкюль . была сильной и очень удлиненной У него были очень надежные резцы и зубья щечных с несколькими пульсами (для которых названы многотуркуляты). Тазовые кости отличались от других многотуркуляций в том, что они не были слиты друг с другом. Catopsbaatar имел шпоры на его лодыжках, как и у мужского платок и эхидны , без признаков ядовитого канала (присутствующего в первом).
Шпоры Catopsbaatar и других мезозойских возможно, использовались для защиты от динозавров млекопитающих , и других хищников. Считается, что мультитуберкуляты рожают живое рождение , и тот факт, что у них были волосы, указывает на то, что они были гомеотермическими («теплые»). Многотуберкуляты были бы всеядными ; Catopsbaatar имели мощные мышцы челюсти, и его резцы были хорошо адаптированы для грызения твердых семян, используя обратный жевательный ход. Считается, что многотуркуляты имели обширную позу, и Catopsbaatar , возможно, смог прыгнуть. Catopsbaatar известен из формирования Barun Goyot , которой, как считается, около 72 миллионов лет.
Таксономия
[ редактировать ]
В 1970 и 1971 годах лако-монгольские палеонтологические экспедиции собрали окаменелости млекопитающих из формирования Баруна Гойота Монголии на красных ложах Гермиина Цава (также написанного «Хермин Цав») в пустыне Гоби . Около 100 образцов, извлеченных из четырех мест, размещаются в Польской академии наук в Варшаве. Две трети собранных образцов были многотуркуляциями : вымерший порядок млекопитающих с зубными зубными зубами, названным в честь многочисленных шипков (или бугорков ) на их молярах . В 1974 году польский палеонтолог Зофия Килан-Яворовский назвал новым видом монгольского многотуркуляционного рода Djadochtatherium в качестве D. catopsaloides , с образцом zpal mgm-I/78 из коллекции польской в качестве голотипа . Конкретное название относится к сходству животного с североамериканскими видами Cotopsalis Joyneri , который, как думал Киланн-Джаворовский возможным потомком. Образец, собранный в местности Hermiin Tsav I, является почти полным черепом юношеского с частями поврежденного черепа. Киелан-джаворовская также назначил другие образцы видам: поврежденный череп отсутствует нижние челюсти (ZPAL MGM-I/79, взрослый), череп с частичными нижними челюстями (ZPAL MGM-I/80) и моляр с фрагментом челюсти (ZPAL MGM - I/159 от Хулсана, единственный образец, не из которых не из местностей Hermiin Tsav I и II). [ 1 ] [ 2 ] [ 3 ] [ 4 ]

Роберт Э. Слоан считали род Djadochtatherium младшим синонимом Catopsalis Киелан-Джаворовский и американский палеонтолог и создали новую комбинацию C. catopsaloides в 1979 году. [ 5 ] Американские палеонтологи Нэнси Б. Симмонс и Мяо Десуи провели кладистический анализ 1986 года , который показал, что Catopsalis был парафилетическим таксоном (неестественная группировка видов), а C. catopsaloides требовал собственного общего названия. [ 6 ] Килан-Джаворовский последовал за предложением Симмонса и Мяо, перенесему C. catopsaloides к своему собственному монотипическому роду в 1994 году, Catopsbaatar . Слово Catops получено из греческого катоптоса («видимого» или «очевидного»); Баатар является монгольским для «героя», и название относится к сходству Catopsbaatar с родом Catopsalis (как в случае конкретного имени). [ 7 ] Само название Catopsalis состоит из греческих слов для «видимых» и «режущих ножниц» ( псалис ). [ 8 ] [ 7 ] Слово Baatar используется в качестве суффикса в названиях многих многотуркуляционных родов и намекает на монгольский столицу Улаанбаатара , что само по себе означает «красный герой». [ 9 ]
Позже, в 1994 году, Киелан-Джаворовская и русский палеонтолог Петр П. Гамбарян упомянули каудальные (хвостовые) позвонки, которые могли принадлежать Catopsbaatar ; Эта атрибуция неопределенна, поскольку они могут вместо этого принадлежать к соответствующему Томбаатару (названный в 1997 году). [ 10 ] [ 11 ] Четвертый череп (PIN 4537/4, несовершеннолетний), обнаруженный во время советско-Монгольской экспедиции 1975 года, был упомянут Гамбарян и Киелан-Яворовской в 1995 году. [ 12 ] Канадский палеонтолог Филипп Дж. Керри обнаружил новый образец Catopsbaatar во время динозавров 1999 года экспедиции Гоби, организованного американской компанией по кочевым экспедициям. Расположенный в Монгольской академии наук в Улаанбаатаре в качестве PM120/107, этот наиболее полностью сохранившийся известный образец состоит из полного черепа (который может быть слегка сплющен) и частичного скелета взрослого человека. Образец имеет довольно полные передние и задние конечности, которые были неизвестны для рода до тех пор и которые обычно редко сохраняются в многотуркуляциях. Его тазовая ильена была украдена и разрушена школьником в туре в Музее естественной истории в Осло , где он был подготовлен в 2000 году. Образец был зарегистрирован в 2002 году Киелан-Джаворовской, норвежским палеонтологом Йорн Хурум , Керри и монгольским палеонтологом Ринчен Барсболд. , которые также упомянули еще один череп (PIN 4537/5, ювенильный), обнаруженный во время экспедиции 1975 года. [ 11 ] [ 13 ] [ 3 ] Catopsalis Joyneri , основа и названия C. catopsaloides , была перенесена в новый род Valenopsalis в 2015 году. [ 8 ] [ 1 ]
Эволюция
[ редактировать ]Catopsbaatar принадлежал Order Multituberculata, группе в Allotheria (инфракла млекопитающих за пределами Терии , группы, которая содержит современные плаценты и сумчатые ). Многотуркуляты характеризуются наличием премоляров и моляров с несколькими низкими пульсами, расположенными в продольных рядах. Они являются самой известной группой млекопитающих из мезозойской эры , когда динозавры доминировали ; Хотя самые ранние многотуркулятные останки находятся в юрском периоде , группа известна как недавно как эпоха эоцена (тем самым выживая на мели -палеогеновом вымирании ). Группа, возможно, вымерла из -за конкуренции с эврианскими млекопитающими, такими как грызуны. Многотуркуляты были в основном известны из зубов и челюстей до 1920 -х годов, когда были обнаружены более полные образцы - впервые в Азии, а затем в другом месте. [ 4 ] Посткраниальные кости (остальная часть скелета, кроме черепа) остаются редкими. [ 13 ]
Kielan-Jaworowska первоначально классифицировал Catopsbaatar как член MultiTuberculate Family Taeniolabididae в 1974 году. [ 1 ] В 1994 году она предположила, что Джадохтатериум был близок к Catopsbaatar происхождению . [ 7 ] многотуркулятов Djadochtatheriidae они помещали в новую подготовку Djadochtatheria ) и ( новую которую семью . Она Хурум назвали Семейство отличается от других многотуркулятов (и других млекопитающих) тем, что передние края ее морды были сливались с зигоматическими арками (скулы), придавая морду трапеции формы, если увидеть сверху. В целом, у других млекопитающих есть морды, где боковые края изогнуты внутрь перед зигоматическими арками. [ 14 ] Киелан-Джаворовская и Хурум пересмотрели более высокие ряды в мультитуберкулате в 2001 году, заменив подряд Джадохтатерии суперсемейством Джадохтатериоидеа (помещенного в подзарядку Cimolodonta ). [ 15 ]
Следующая кладограмма показывает размещение Catopsbaatar среди других многотуркуляций, согласно Kielan-Jaworowska and Hurum, 1997: [ 14 ]
Описание
[ редактировать ]Череп
[ редактировать ]
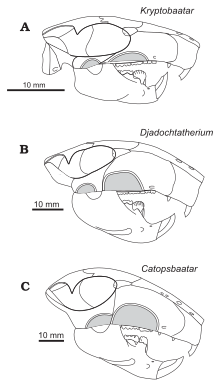
Самый полный череп для взрослых Catopsbaatar (образец PM 120/107) составляет 63 мм (2,5 дюйма) длиной и шириной 55 мм (2,2 дюйма), с нижней челюстью длиной 41 мм (1,6 дюйма). Для сравнения, череп ювенильного голотипа (ZPAL MGM-I/78) составляет около 53 мм (2,1 дюйма) длиной и шириной 56 мм (2,2 дюйма), с нижней челюстью длиной 35 мм (1,4 дюйма). Самый большой череп для взрослых (ZPAL MGM - I/79) составляет длину 70 мм (2,8 дюйма), но, поскольку он неполный, другие его измерения неизвестны. Catopsbaatar был больше, чем его родственники, Kryptobaatar и Djadochtatherium . [ 3 ] Многотуберкуляты имели относительно большие черепа и короткие шеи; Их черепа были пропорционально дольше и шире, чем у грызунов и сумчатых аналогичного размера. [ 4 ] Внешний внешний вид их голов мог быть похож на кадры грызунов. [ 12 ]
Череп Catopsbaatar был тяжелым, с широким отрывом на передней части. Это было короче вдоль средней линии, чем по бокам, потому что герб нуч в задней части головы изогнулся внутрь в середине, создавая недостаток на заднем крае черепа, если смотреть сверху. Зигоматические арки были сильно расширены до бок, с шириной черепа (через арки) около 85 процентов от длины черепа. Передняя часть орбит (глазных глаз) была дальше, чем у других джадохтатеридов, что привело к более удлиненной морде (65 процентов длины черепа) и небольших орбитах. Пенсисилла (передняя кость верхней челюсти) простиралась менее чем на две трети длины морды перед глазами, короче, чем у Криптобатара . Пенсисилла состояла из частей лица и неба; Джадохтатерииды имели предчелюстный гребень на границе между ними (видимый при просмотре черепа снизу). Пятническая часть предчезгиллы была вогнутой, со многими случайно расположенными питательными веществами Форамина (отверстия). , Носовая кость которая сформировала верхнюю часть морды, была относительно широкой (становилась все шире в сторону спины), а ее фронт был покрыт нерегулярно распределенным сосудистым отверстием. Швар был менее заостренным вперед в середине , между носовыми и фронтальными костями чем у родственников Catopsbaatar . [ 3 ]

( Верхняя челюсть основная часть верхней челюсти) была обширной и сформировала большую часть стороны морды. Он содержал все верхние зубы, кроме резцов . Инфраорбитальный отверстия (отверстия в нижней части верхней челюсти) были похожи на щель в некоторых образцах и округлены в других и варьировались по количеству от одного до трех. Одной из наиболее характерных особенностей лица Catopsbaatar был очень большой передний зигоматический хребет по бокам верхней челюсти (место для прикрепления челюсти мышцы). Это было намного выше, чем у других джадохтатеридов, за исключением Джадохтатериума , от которого он отличался тем, что хребет был полукруглым, а не примерно трапециевидным (другие роды имеют эллиптические хребты). Передний край переднего зигоматического хребта был утолщен, что дало выпуклость на боковом краю морды при просмотре со стороны и выше. Нижняя часть шва между верхней челюстью и сквамозальной костью простиралась вдоль задних границ переднего зигоматического хребта. Пятнические процессы верхней челюсти сформировали большую часть неба. А У основного неглубокого отверстия было неглубокое канавки, простирающиеся вперед. Постпалатиновый торус (костная выступление во вкусе) Catopsbaatar была менее заметной, чем был то Томбаатар . Фронтальная кость была большой и образовала большую часть черепной крыши. Швар между лобными и теменными костями образовал U-образную часть в середине, которая указывала назад-мимолетно с состоянием в криптобатаре , но менее глубоком, с меньшими U-образными структурами по бокам. Это отличалось от Djadochtatherium , у которого был более узкий V-образный швар между лобными и теменными костями. [ 3 ]

Промежуточный зигоматический гребень на сквомольной кости (также для прикрепления челюсти) был намного меньше и ниже, чем передний зигоматический хребет перед ним. Catopsbaatar отличался от других джадохтатеридов тем, что промежуточный хребет связался с передним хребтом с его передним краем. Задний зигоматический гребень в нижней части спины квамосовой кости был самым слабым из трех хребтов и только отмечен депрессией. Посторбитальные процессы , стоящие за орбитой на теменной кости, были очень длинными, и теменные хребты простирались от заднего края посторбитальных процессов друг к другу (но не достигая). Нюшальный гребень был очень заметным и распространялся на стороны, чтобы сформировать «крылья». пластина Затылочная была почти вертикальной и слегка вогнутой, и была скрыта герб Нучала, когда череп был просмотрен сверху. Несмотря на то, что не полностью сохранилась, предполагается, что затылочная пластина отступала назад и вверх от затылочных мыщелков (как у некоторых родственников). [ 3 ] Орбито-временная сосудистая система внутри черепа Catopsbaatar не сильно отличалась от таковых из родственных родов. [ 16 ]
Catopsbaatar Мябильность была надежной и очень удлиненной. Диастема ( разрыв между передними и щеками) был вогнутым и продлевался на 20 процентов зубной кости (основная кость нижней челюсти). Сверху видна диастема сформировала широкую полку, которая наклонилась вниз на внутренней стороне челюсти. Небольшое ментальное отверстие было близко к верхнему среднему краю диастемы. Короноидный процесс нижней челюсти, по -видимому, был относительно длиннее и уже, чем в других джадохтатериоидах. Он был отделен от альвеолярного процесса (где содержатся зубы) широким канавкой. Русиночный мыщел (который сочленен с черепом) был немного выше уровня моляров. Передняя часть Мастерического гребня была очень заметной, образуя выпуклость, известную как мастерное выпуклование. Надежность этого гребня и присутствие выпуклости варьируются среди родственных родов. Мастерическая фовеа (яма) перед массетерической ямкой была, вероятно, более отчетливой, чем в других джадохтатериоидах. Каждая половина Медиблютный симфиз (где две половинки нижней челюсти) был сформирован как перевернутая слеза. Птеригоидная ямка на внутренней стороне нижней челюсти была очень большой и заняла большую часть задней части зубца. Нижняя часть этой ямки имела границу, известную как полка Pterygoideus. [ 3 ]
Зубной зуб
[ редактировать ]
Стоматологическая формула (количество зубов каждого типа в зубном ряду млекопитающего) Catopsbaatar составила 2,0.3.2 1,0.2.2 (два резца, без собак , три премоляра и два моляра в половине верхней части зуба и один Резьки, без собак, два премоляра и два моляра в половине нижней части). Для сравнения, зубная формула людей составляет 2,1.2,2–3 2.1.2.2–3 . Каждый зуб в млекопитающем обозначен буквой и номером по положению (i для резца, C для собак, P для Premolar, M для молярного); Буквы капитализированы для зубов верхней челюсти, но не для тех, кто находится в нижней челюсти. Формула Cusp показывает расположение и количество усилий в последовательных рядах зуба, от внешней до внутренней стороны; Каждый ряд разделен толстой кишкой. [ 3 ] [ 4 ] [ 11 ]
Будучи Cimolodont, Catopsbaatar не имел резцов i1. [ 14 ] Учащивания в верхнем фронте I2 Catopsbaatar были очень надежными и имели резко ограниченную полосу эмали . Два резца слегка сходились к середине, касаясь друг друга. Меньший резец i3 позади был конусный. Альвеол Томбаатара (зубной гнездо) резиса Catopsbaatar I3 был сформирован по предчезгилле, а не предчезгиллой и верхней челюсти (в отличие от ) . Передние верхние премоляры P1 и P3 присутствовали только у несовершеннолетних ( лиственных ), исчезая (с их альвеолами) у пожилых людей. P1, по-видимому, имел два шин, был однократным и имел тупой корону , похожей на конус . P3 был однократным и меньше P1. Формула CUSP премонкой P4 составляла 5-4: 1, и центральный порог является самым большим. P4 Catopsbaatar был почти трапециевидной по форме (в отличие от Djadochtatherium и Kryptobaatar , где он находится в форме полумесяца), меньше и не хватает хребтов. Catopsbaatar также отличался тем, что имел только три верхних премоляра, не имея P2 (функция, общая с Tombaatar ). Другие млекопитающие обычно развивают потерю зубов в начале или конце зубного ряда, а не в середине (как в многотуркуляции). Формула Cusp моляра M1 составляла 5–6: 5–6: 4, а внутренний гребень простирался около 75 процентов от длины зуба. Очень остроконечные и невыраженные в несовершеннолетних, но они носили и вогнуты у пожилых животных. Формула Cusp молярного M2 составила 2: 2–3: 2–3. [ 3 ] [ 4 ]
У Catopsbaatar была одна нижняя пара резцов, характерных для многотуркулятов, которые были очень сильными и сжатыми вбок. У него была резко ограниченная полоса эмали, и он постоянно рос. P3 Premolar был очень маленьким и полностью придерживался нижней диастемы под большим P4. Подобный лезвию P4 был примерно трапециевидным по визу сбоку и имел три острых отвода вдоль горизонтального верхнего края и один остров на внешней задней стороне. У P4 не было хребтов на внешней и внутренней стороне, как присутствуют в других многотуркуляциях. Молярный M1 был почти симметричным, и его формула Cusp составляла 4: 4, размер сливочных элементов, уменьшающихся в сторону спины. M2 имел формулу Cusp 2–3: 2, большинство образцов составляли 2: 2. Выловы на внутренней стороне были шире, чем на внешней стороне, внутренний ряд тяжбы был короче, чем внешний, а задний край зуба был устроен наклонно. [ 3 ]
Посткраниальный скелет
[ редактировать ]
Единственный образец Catopsbaatar , который сохраняет посткраниальный скелет, - это PM120/107, который является фрагментарным. Он включает в себя элементы, которые неизвестны или не полностью сохранены, в других многотуркуляциях. Один поясничный позвонок (пятый или шестый, от грудной клетки и таза) имел остистый процесс, который был крепким по визу и долго, когда они видели сверху. Ключица была немного менее изогнутой , чем у криптобатара (напоминающий изогнутый стержень, который расширялся на каждом конце), и измерял около 24,8 мм (0,98 дюйма). Верхняя часть лопатокоракоида была относительно широкой по визу сбоку, а нижняя часть была очень узкой. Вероятно, это было около 60 мм (2,4 дюйма) длиной, когда завершено. Сохраненная часть плечевой кости (кость верхней части руки) составляла около 37,5 мм (1,48 дюйма). Его стержень был треугольным по поперечному сечению, относительно узким, когда они видели сверху, и большая часть его ширины была занята межберкулярной канавкой . Эта канавка была разграничена на стороне гребнем Большого бугорка , средняя часть которого образовала дельтопекторный гребень. Локтевой мыщел, где Ложа нижняя рука, сформулированная с плечами, была более заметной, чем радиальный мыщел (где сформулирован радиус), овальный и разграниченный из радиального мыщелка с помощью канавки. Радиус был длиной около 26 мм (1,0 дюйма) с выдающейся головой. Его вал был гладким, сжатым сверху вниз и овальным по поперечному сечению. Устань сжималась в боковой основе, более плоской, чем радиус, и около 40 мм (1,6 дюйма), как сохранилось. [ 13 ]

Контакт от ишия к подвздошной кости и лобкотике таза не слился, а передний конец ишия образовал морщинистый швар. Лобок был примерно треугольным, с грубым швом для подвздошного подвздошного ткани и глубокой канавкой для ишия в нижнем фронте. Образец PM120/107 тазовых костей отличались от других мультитуберкуляций, не слитых вместе. Присутствие швов в тазе PM120/107 указывает на то, что это был несовершеннолетний, хотя череп кажется взрослым; Значение этого несоответствия неизвестно. Бедра (бедра) была пропорционально похожа на эриф Эукосоммодона и Немегтбаатара - ловца, чем первый, но больше, чем у последних. Бедра была крепкой по сравнению с его длиной, и, возможно, длиной около 56 мм (2,2 дюйма). Цвета голени нижней ноги составляла около 35,8 мм (1,41 дюйма). Из -за верхней стороны у голени были глубокие раскопки (полость), которые могут быть характерны для многотуркуляции. [ 13 ]
В отличие от большинства других многотуркулятов и других млекопитающих, кальцевая кость в задней части стопы имела короткий клубень кальциреи (например, некоторые кенгуру из дерева ), с расширенным проксимальным процессом в форме конва, сильно изогнутой вниз и в стороне. Catopsbaatar имела кость OS Calcaris на внутренней стороне его лодыжки, что также наблюдается у современных мужских монотримов ( утконос и эхидна ) и других мезозойских млекопитающих. ОС Calcaris кость была похожей на пластинку и прямоугольной в контур; Как и в монотриме, он поддерживал Cornu Calcaris в формировании шпоры на внешней стороне лапки (кластер костей ног). В отличие от других мезозойских млекопитающих, эти два элемента не были объединены вместе в многотуркуляциях. Cornu Calcaris был треугольным, с вогнутостью в середине и длиной 13 мм (0,51 дюйма). Шпора была сплющена и была толще в его соединении с ОС Калгарисом (где они соединялись через несколько хребтов). Поскольку подростка PM120/107, возможно, была перемещена с его первоначальной позиции, неизвестно, столкнулся ли он внутрь (как на утконосе). В отличие от утконос, не было впечатления о канале для яд . Cornu Calcaris из Catopsbaatar был окостенен (превращен в кость) и был бы покрыт кератином (возбужденное покрытие, наблюдаемое в ногтях и копытах). Cornu Calcaris утконоса состоит только из кератина и пустоте. [ 13 ] [ 17 ]
Палеобиология
[ редактировать ]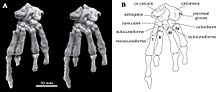
Hurum, Zhe-xi luo и Kielan-jaworowska предположили в 2006 году, что шпоры на лодыжках мезозойских млекопитающих (таких как Catopsbaatar ) были гомологичными с монотриями и были базальными (или «примитивными»), потерянной позже млекопитающие. Самский утконос использует шпобу, чтобы доставить яд из железы, но неизвестно, были ли вымершие группы также ядовиты. размером с лисы Мезозойские млекопитающие были в основном небольшими (за исключением, такими как репеномамус ), и, хотя они были слишком малы, чтобы быть жертвами для крупных динозавров с теропод , меньшие тероподы, большие ящерицы, крокодилы и птицы могли питаться на них. Например, челюсти из млекопитающих были обнаружены в животе образца малого теропод Sinosauropteryx ; Челюсти принадлежали Чангеотерауму , в котором также были шпоры, и многотуркуляция Синобаатара . Поскольку динозавры доминировали в Земле во время мезозойского, этот период был назван « темными веками » истории млекопитающих. Споры, которая была бы более эффективной, если бы ядовилась, вероятно, использовалась в качестве защитного оружия маленькими ранними млекопитающими. Он также мог быть использован во время внутривидовая конкуренция или хищничество. [ 17 ]

Тазовые кости образца Catopsbaatar PM120/107, возможно, не были слиты, потому что слияние произошло поздно в разработке, потому что это была сексуально диморфная особенность, возникающая только у мужчин (невзванные тазовые кости могли позволить расширить родовой канал у женщин), или в пельвике Слияние может быть таксономическим различием между Catopsbaatar и другими многотуркуляциями. [ 13 ] В отличие от других млекопитающих, таз мультитуберкулятов была очень узкой; В других родах, где известен таз, каждая половина лобкового лобка и ишия была объединена вместе, образуя киль. Длина и жесткость киля указывают на то, что таз не мог распространяться во время рождения. Поскольку для прохождения яйца (яйцеклеточные монотрины было мало места (у монотрий с яйцом широкие ишиальные арки), Килан-Джаворовска в 1979 году предположил, что многотуркуляты были вивипарусными (дали живые роды) и что новорожденные были чрезвычайно маленькими-не менее сумчатые. [ 4 ]
Волосы, структурно сходные с волосами современных млекопитающих и связанные с костями монгольских многотуркуляции лямбдопсиса , были идентифицированы в копролитах (ископаемые фекалии) плотоядных млекопитающих из эпохи палеоцена . Это указывает на то, что в мультитуберкулятах были волосы для изоляции, аналогично современным млекопитающим (и, возможно, ископаемым млекопитающим), что, вероятно, связана с гомеотермией (теплокровная). [ 4 ] [ 18 ]
Кормление и диета
[ редактировать ]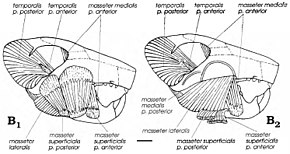
Несмотря на то, что мультитуберкуляты считались плотоядными или травоядными, поскольку американский палеонтолог Уильям А. Клеменс и Килан-Яворовская предложили современные крысы кенгуру в качестве аналогов для группы в 1979 году, они считались всеядными (кормить как растения, так и животных). [ 4 ] Уникально среди млекопитающих многотуркуляты использовали обратный жевательный удар, который приводил к жевальным мышцам - мышцам, которые перемещают нижнюю челюсть - вставлялись больше к фронту, чем в других группах (включая грызунов). Gambaryan и Kielan-Jaworowska реконструировали мистическую мускулатуру различных мультитуркуляций в 1995 году и обнаружили, что Catopsbaatar и его родственники имели очень мощную шагтирующую мускулатуру из-за их высоких зигоматических арков и больших передних и промежуточных зигоматических выступов и короноидных процессов. Их мощные резцы, с ограниченными полосами эмали, были бы хорошо адаптированы к грызу и крезанию твердых семян (аналогично грызунам). Поскольку он был больше, чем некоторые другие многотуркуляции, Catopsbaatar должен был бы открыть рот всего на 25 градусов, чтобы раздавить твердые семена 12–14 мм (0,47–0,55 дюйма) в диаметре; 40-градусный зазерство вызвало бы дислокацию. После того, как резцы сократились, премоляры и моляры начнут измельчать с «силовым ударом». [ 12 ]
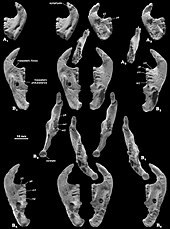
Согласно Гамбарян и Киелан-Джаворовской, иногда адаптация для сокрушительных тяжелых семян-как в Catopsbaatar -нацелена на пользу низкого мыщелкового процесса (который препятствует вывиху нижней челюсти). Передние и промежуточные зигоматические хребты черепа были происхождением поверхностной мышцы массы , которая облегчает жевание. Разделение происхождения этой мышцы на две части, а округлые мышечные шрамы, оставшиеся ими, являются уникальными среди млекопитающих для многотуркуляции. Печатные мышцы многотуркуляции независимо развивались функции, разделяемые с грызунами и мелкими травоядными сумчарыми. Как и в случае с грызунами, мультитуберкуляты могли быть способны к двустороннему желудочке - где оба ряда зубов в нижней челюсти выполняют одновременно одновременно одновременно - и одностороннее желучение (где используются ряды с одной стороны). [ 3 ] [ 12 ]
Осанка и передвижение
[ редактировать ]
Обсуждается позы в Limb Multuberculates. По словам некоторых исследователей, они использовали паразагиттальную позицию с прямыми конечностями под организмом; Другие считают более вероятной обширной позиции. Киелан-Джаворовская и Хурум поддерживали последнюю теорию в 2006 году на основе присутствия шпоров за задние ноги, что, по их мнению, присутствует только у обширных млекопитающих. Они указали, что все ранние млекопитающие, сохранившие в озерных (озеро) отложения, были сжаты сверху вниз, что указывает на обширную позицию, тогда как более поздние млекопитающие сохранились на их флангах. Более ранние аргументы в пользу обширной позиции включают глубокие таза и особенности ног. Они также предположили, что ноги многотуркулятов были бы в покое Plantigrade (подошва, касающаяся земли), но Digitigrade (подошва, не касаясь земли), когда прыгает и работают быстро; Они отклонили идею о том, что передние конечности многотуркулятов и других ранних млекопитающих были более паразагиттальными, чем их задние конечности. Киелан-джаворовская и Хурум изображены Catopsbaatar с Plantigrade, растягивающимися ногами, с мобильными шпорами, которые указывали внутрь при подготовке к атаке. [ 19 ] [ 4 ]
В 2008 году Киелан-Яворовская и Хурум предположили, что длинный остистый процесс на позвонке Catopsbaatar и длинные поперечные процессы в Nemegtbaatar могут указывать на то, что некоторые мультитуберкуляты были солковыми (имели способность прыгать). Catopsbaatar , вероятно, имел сильные мышцы, прикрепляющиеся к клубеньку кальцилеи, что дополнительно подтверждает гипотезу прыжков. [ 13 ] [ 20 ] Хотя было высказано предположение, что многотуркуляты были древесными (жили на деревьях), большинство азиатских таксонов, вероятно, были наземными; Некоторые другие были подачами , копали и жили под землей. [ 4 ]
Палеоэрона
[ редактировать ]
Все образцы Catopsbaatar известны из формирования Баруна Гойота пустыни Гоби, что, вероятно, датируется поздней кампанианской эпохи поздней меловой эпохи (около 72 миллионов лет назад). Образцы были обнаружены в красных пластах области Гермиина Цав, за исключением одного моляра из Хулсана. Когда был обнаружен Catopsbaatar , полагалось, что красные пласты в области Гермиина Цав были отдельной ковальной формированием (одного и того же геологического возраста) с формированием Barun Goyot, поскольку они содержат много одинаковых животных, но теперь считается, что они соответствуют разным Уровни формирования Barun Goyot и Nemegt. [ 3 ] [ 21 ] [ 22 ] Каменные фации красных слоев области гермиин цав состоят из толстого песчаника из оранжевого цвета с тонким переплетенным иеловым камнями и глиняными камнями . [ 23 ] [ 3 ] Считается, что каменные фации формирования Barun Goyot являются результатом засушливой или полузасушливой среды, с эолианскими (откладываемыми ветровыми) кроватями. [ 24 ] [ 22 ] Другие известные млекопитающие из красных слоев Hermiin Tsav включают в себя многотуркуляции Nemegtbaatar , Chulsanbaatar и Nessovbaatar , а также Deltatheridium , Asioryctes и Barunlestes . [ 25 ] Динозавры включают Heyuannia , Velociraptor , Saichania , Platyceratops , Gobiceratops и некоторые неопределенные тероподы . черепахи Рептилии включают в себя монголемы , ящерицы Гобинатус , Чингисавр , Продюя , Гладиденагама и Фриносомимус , а также неопределенный крокодил. и лягушка неопределенная алексиальная Также известны птица. Остракоды включают лимноцитхер , киприду и eucypris . [ 26 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный в Килан-Джаворовский, З. (1974). «Многотуркуляция преемственности в поздней меловой пустыне Гоби (Монголия)» (PDF) . Palaeontologica Polonica . Результаты польских монгольских палеонтологических экспедиций-Часть V. 30 : 23–43. Архивировано (PDF) из оригинала 2017-05-05 . Получено 2018-03-20 .
- ^ Dashzeveg, D.; Новацек, MJ; Норелл, Массачусетс; Кларк, JM; Chiappe, LM; Дэвидсон, А.; МакКенна, MC; Dingus, L.; Swisher, C.; Altagerel, P. (1995). «Необычайное сохранение в новой сборке позвоночных от позднего мела Монголии». Природа . 374 (6521): 446–449. Bibcode : 1995natur.374..446d . doi : 10.1038/3744446A0 . S2CID 4245878 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м Kielan-Jaworowska, Z.; Хурум, JH; Lopatin, AV (2005). «Структура черепа в Catopsbaatar и зигоматические хребты у многотуркуляции млекопитающих» . Acta Palaeontologica Polonica . 50 (5). Архивировано с оригинала 2018-02-24 . Получено 2018-03-03 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Kielan-Jaworowska, Z.; Cifelli, RL; Luo, Z.-X. (2004). Млекопитающие с эпохи динозавров: происхождение, эволюция и структура . Нью -Йорк: издательство Колумбийского университета. С. XIV, 249, 260–261, 279, 294–299. ISBN 978-0-231-11918-4 .
- ^ Kielan-Jaworowska, Z.; Слоан, Re (1979). « Catopsalis (Multituberculata) из Азии и Северной Америки и проблема рассеивания таениолабидидов в позднем меловом периоде» . Acta Palaeontologica Polonica . 24 (2): 187–197. Архивировано с оригинала 2018-04-30 . Получено 2018-04-29 .
- ^ Симмонс, NB; Десуи М. (1986). «Парафила в Catopsalis (Mammalia: Multituberculata) и его биогеографические последствия». Вклад в геологию, Университет Вайоминга . 24 (3): 87–94. doi : 10.2113/gsrocky.24.special_paper_3.87 .
- ^ Jump up to: а беременный в Килан-Джаворовский, З. (1994). «Новое общее название для многотуркуляции млекопитающих« djadochtatherium » Catopsaloides » . Acta Palaeontologica Polonica . 39 (1): 134–136. Архивировано с оригинала 2018-04-30 . Получено 2018-04-29 .
- ^ Jump up to: а беременный Уильямсон, те; Брусатте, SL; Secord, R.; Шелли С. (2016). «Новый таениолабидоидный мультитуберкуляция (млекопитающие) из среднего пуканта формирования Нацимиенто, Нью -Мексико и пересмотр таениолабидоидной систематики и филогения» . Зоологический журнал Линневого общества . 177 (1): 183–208. doi : 10.1111/Zoj.12336 . Архивировано из оригинала 2017-09-22 . Получено 2018-10-22 .
- ^ Lillegraven, Ja; Kielan-Jaworowska, Z.; Клеменс, Вашингтон (1979). Мезозойские млекопитающие: первые две трети истории млекопитающих . Беркли: Университет Калифорнийской прессы. п. 35. ISBN 978-0520035829 .
- ^ Килан-Джовавска, Z.; Palmary, PP (1994). Помимо мультикультурного Летая . 27 (4): 300. 10.111 / j.1502-3931.1994 .
- ^ Jump up to: а беременный в Kielan-Jaworowska, Z.; Хурум, JH; Керри, PJ; Барсболд Р. (2002). «Новые данные об анатомии позднего мела -многотуркуляции млекопитающих Catopsbaatar » . Acta Palaeontologica Polonica . 47 (3): 557–560. Архивировано с оригинала 2018-04-30 . Получено 2018-04-29 .
- ^ Jump up to: а беременный в дюймовый Gambaryan, pp; Килан-Джаворовский, З. (1995). «Печатная мускулатура азиатских таениолабидоидных мультитуберкуляционных млекопитающих» . Acta Palaeontologica Polonica . 40 (1): 45–108. Архивировано с оригинала 2018-02-24 . Получено 2018-03-03 .
- ^ Jump up to: а беременный в дюймовый и фон глин Хурум, JH; Килан-Джаворовская, З. (2008). «Посткраниальный скелет мелового мультитуркуляции млекопитающих Catopsbaatar » . Acta Palaeontologica Polonica . 53 (4): 545–566. doi : 10.4202/app.2008.0401 .
- ^ Jump up to: а беременный в Kielan-Jaworowska, Z.; Hurum, JH (1997). «Джадохтатерия - новый подряд многотуркуляции млекопитающих» . Acta Palaeontologica Polonica . 42 (2): 201–242. Архивировано с оригинала 2018-04-30 . Получено 2018-04-29 .
- ^ Kielan-Jaworowska, Z.; Hurum, JH (2001). «Филогения и систематика многотуркуляции млекопитающих» (PDF) . Палеонтология . 44 (3): 389–429. doi : 10.1111/1475-4983.00185 . S2CID 83592270 .
- ^ Kielan-Jaworowska, Z.; Пресли, Р.; Поплин, Сесиль (1986). «Сосудистая система черепа у таениолабидоидных мультитуберкуляционных млекопитающих». Философские транзакции Королевского общества Лондона. Серия B, биологические науки . 313 (1164): 525–602. BIBCODE : 1986RSPTB.313..525K . doi : 10.1098/rstb.1986.0055 . JSTOR 2396476 .
- ^ Jump up to: а беременный Хурум, JH; Luo, ZX; Килан-Джаворовский, З. (2006). "Были ли млекопитающие изначально ядовитыми?" Полем Acta Palaeontologica Polonica . 51 (1): 1–11. Архивировано из оригинала 2017-10-27 . Получено 2018-03-10 .
- ^ Meng, J.; Wyss, AR (1997). «Многотуркуляция и другие волосы млекопитающих, извлеченные из экскрементов палеогена». Природа . 385 (6618): 712–714. Bibcode : 1997natur.385..712m . doi : 10.1038/385712A0 . PMID 9034186 . S2CID 4325160 .
- ^ Kielan-Jaworowska, Z.; Hurum, JH (2006). «Осанка конечностей у ранних млекопитающих: обширные или парасагиттальные» . Acta Palaeontologica Polonica . 51 (3): 393–406. Архивировано из оригинала 2017-10-08 . Получено 2018-03-13 .
- ^ Чен, М.; Уилсон, GP (2015). «Многомерный подход к выводу локомоторных мод у мезозойских млекопитающих». Палеобиология . 41 (2): 280–312. doi : 10.1017/pab.2014.14 . S2CID 86087687 .
- ^ Gradziński, R.; Kielan-Jaworowska, Z.; Марианска Т. (1977). «Верхние меловые джадохты, барун Гойот верхний меловый и Немегт Формации Монголии, включая замечания о предыдущих подразделениях» . Acta Geologica Polonica . 7 (3). Архивировано с оригинала 2018-03-14 . Получено 2018-03-13 .
- ^ Jump up to: а беременный Эберт, Да (2018). «Стратиграфия и палеоэкологовая эволюция богатой динозаврой-динозавром последовательности Baruungoyot-nemegt (верхний меловой), бассейн Немегта, южная Монголия». Палеогеография, палеоклиматология, палеоэкология . 494 : 29–50. Bibcode : 2018ppp ... 494 ... 29e . doi : 10.1016/j.palaeo.2017.11.018 .
- ^ Gradziński, R.; Jerzykiewicz, T. (1972). «Дополнительные географические и геологические данные от польских монгольских палеонтологических экспедиций» (PDF) . Palaeontologia Polonica . 22 : 17–32. Архивировано (PDF) из оригинала 2014-08-08 . Получено 2018-03-13 .
- ^ Эберт, да; Badamgagarav, D.; Керри, PJ (2009). «Переход Baruantoit-Nemegt (Pupper Cretaceous) в области Немета, бассейн Немегта, Южная Центральная Монголия » 고생 Pro -. 25 (1). ISSN 1225-0 Архивировано с оригинала 2018-06-1 Получено 2018-03-1
- ^ Kielan-Jaworowska, Z.; Новацек, MJ; Трофимов, Б.А.; D., Dashzeveg (2000). «Млекопитающие из мезозоя Монголии». В Бентоне, MJ; Шишкин, Массачусетс; Unwin, DM; Kurochkin, en (eds.). Возраст динозавров в России и Монголии . Кембридж: издательство Кембриджского университета. С. 573–626. ISBN 978-0521545822 .
- ^ Alroy, J. "Khermeen Tsav, Red Breads [SMPE] (PIN Coll. 3142) (меловая из Монголии)" . FossilWorks.org . FossilWorks: Gateway в базу данных палеобиологии . Получено 17 декабря 2021 года .
Внешние ссылки
[ редактировать ] СМИ, связанные с Catopsbaatar в Wikimedia Commons
СМИ, связанные с Catopsbaatar в Wikimedia Commons  Данные, связанные с Catopsbaatar в Wikispecies
Данные, связанные с Catopsbaatar в Wikispecies
| ||||
| ||||

