Outermost of the three layers that make up the skin
Эта статья о коже млекопитающих. Чтобы узнать о других значениях, см. Эпидермис (значения) .
Эпидермис
Микроскопическое изображение эпидермиса, составляющего внешний слой кожи, показано здесь белой полосой.
Микроскопическое изображение, показывающее слои эпидермиса. на Роговой слой этом изображении выглядит более компактным, чем на изображении выше, из-за различной подготовки образца.
Слово «эпидермис» происходит от латинского » слова « эпидермис , от древнегреческого слова «эпи » — «над», и от древнегреческого « дерма» — «кожа». Что-то, связанное с эпидермисом или его частью, называется эпидермальным.
Клетки эпидермиса тесно связаны между собой и служат плотным барьером против внешней среды. Соединения между эпидермальными клетками относятся к типу слипчивых соединений , образованных трансмембранными белками, называемыми кадгеринами . Внутри клетки кадгерины связаны с актиновыми нитями. При иммунофлуоресцентной микроскопии сеть актиновых филаментов выглядит как толстая граница, окружающая клетки. [ 9 ] хотя актиновые нити на самом деле расположены внутри клетки и идут параллельно клеточной мембране. Из-за близости соседних клеток и плотности контактов актиновая иммунофлуоресценция выглядит как граница между клетками. [ 9 ]
Конфокальное изображение рогового слоя Состоит из 10–30 слоев многогранных безъядерных корнеоцитов (последний этап дифференцировки кератиноцитов ), причем наибольшее количество слоев приходится на ладони и подошвы. Корнеоциты содержат белковую оболочку (белки ороговевшей оболочки) под плазматической мембраной, заполнены водоудерживающими кератиновыми белками, соединены между собой через корнеодесмосомы и окружены во внеклеточном пространстве сложенными слоями липидов . [ 11 ] В этом слое локализуется большая часть барьерных функций эпидермиса. [ 12 ]
прозрачный/полупрозрачный слой ( stratum lucidum , только на ладонях и подошвах)
Этот узкий слой имеется только на ладонях и подошвах. Эпидермис этих двух областей известен как «толстая кожа», поскольку благодаря этому дополнительному слою кожа имеет 5 эпидермальных слоев вместо 4.
Конфокальное изображение гранулезного слоя Кератиноциты теряют ядра , а их цитоплазма становится зернистой. Липиды, содержащиеся в кератиноцитах пластинчатых тел , высвобождаются во внеклеточное пространство посредством экзоцитоза , образуя липидный барьер, который предотвращает потерю воды из организма, а также проникновение инородных веществ. Эти полярные липиды затем превращаются в неполярные липиды и располагаются параллельно поверхности клетки. Например, гликосфинголипиды становятся церамидами , а фосфолипиды — свободными жирными кислотами . [ 11 ]
Конфокальное изображение шиповатого слоя, уже показывающее некоторые скопления базальных клеток. Кератиноциты соединяются через десмосомы и производят пластинчатые тельца изнутри Гольджи , обогащенные полярными липидами, гликосфинголипидами , свободными стеролами , фосфолипидами и катаболическими ферментами. [ 3 ] В середине этого слоя расположены клетки Лангерганса, иммунологически активные клетки. [ 11 ]
Как многослойный плоский эпителий , эпидермис поддерживается за счет деления клеток в базальном слое. Дифференцирующиеся клетки отслаиваются от базальной мембраны и перемещаются наружу через слои эпидермиса, проходя несколько стадий дифференцировки, пока в роговом слое не теряют свое ядро и не сливаются с плоскими листками, которые в конечном итоге отслаиваются от поверхности ( десквамация ). Дифференцированные кератиноциты секретируют кератиновые белки, которые способствуют образованию внеклеточного матрикса , который является неотъемлемой частью барьерной функции кожи. В нормальной коже скорость производства кератиноцитов равна скорости их потери. [ 4 ] Клетке требуется около двух недель, чтобы пройти путь от базального слоя до верхушки зернистого слоя, и еще четыре недели, чтобы пересечь роговой слой. [ 2 ] Весь эпидермис заменяется ростом новых клеток в течение примерно 48 дней. [ 13 ]
Дифференциация кератиноцитов по всему эпидермису частично опосредована градиентом кальция , который увеличивается от базального слоя до внешнего зернистого слоя, где он достигает максимума, и снижается в роговом слое. Концентрация кальция в роговом слое очень низкая, отчасти потому, что эти относительно сухие клетки не способны растворять ионы. Этот градиент кальция соответствует дифференцировке кератиноцитов и поэтому считается ключевым регулятором формирования эпидермальных слоев. [ 3 ]
Повышение внеклеточной концентрации кальция индуцирует увеличение внутриклеточной концентрации свободного кальция. [ 14 ] Частично это внутриклеточное увеличение происходит за счет кальция, высвобождаемого из внутриклеточных запасов. [ 15 ] а другая часть поступает за счет трансмембранного притока кальция, [ 16 ] через оба кальций-чувствительных хлоридных канала [ 17 ] и потенциалнезависимые катионные каналы, проницаемые для кальция. [ 18 ] Более того, было высказано предположение, что внеклеточный кальций-чувствительный рецептор (CaSR) также способствует повышению внутриклеточной концентрации кальция. [ 19 ]
Клетки зернистого слоя не делятся, а вместо этого образуют клетки кожи, называемые кератиноцитами, гранул кератина из . Эти клетки кожи в конечном итоге становятся ороговевшим слоем ( роговым слоем ), самым внешним эпидермальным слоем, где клетки превращаются в уплощенные мешочки, ядра которых расположены на одном конце клетки. После рождения эти самые внешние клетки заменяются новыми клетками из зернистого слоя, и на протяжении всей жизни они теряются со скоростью 30–90 миллиграммов чешуек кожи каждый час или 0,720–2,16 граммов в день. [ 21 ]
Кислый pH (около 5,0) и небольшое количество воды делают эпидермис враждебным для многих микроорганизмов. [ 12 ]
Непатогенные микроорганизмы на поверхности эпидермиса помогают защититься от патогенов, конкурируя за пищу , ограничивая ее доступность и производя химические выделения , подавляющие рост патогенной микробиоты. [ 12 ]
Внезапные и значительные изменения влажности рогового слоя изменяют гидратацию таким образом, что это может привести к проникновению патогенных микроорганизмов. [ 23 ]
Количество и распределение меланина пигмента в эпидермисе являются основной причиной изменения цвета кожи у Homo sapiens . Меланин содержится в небольших меланосомах , частицах, образующихся в меланоцитах, откуда они передаются в окружающие кератиноциты. Размер, количество и расположение меланосом различаются в зависимости от расовой группы, но хотя количество меланоцитов может варьироваться в разных частях тела, их количество остается одинаковым в отдельных областях тела у всех людей. В белой и азиатской коже меланосомы упакованы в «агрегаты», а в черной коже они крупнее и распределены более равномерно. Количество меланосом в кератиноцитах увеличивается при воздействии УФ-излучения , в то время как их распределение практически не изменяется. [ 28 ]
Кожа содержит специализированные эпидермальные рецепторные клетки, называемые клетками Меркеля . Исторически роль клеток Меркеля в восприятии прикосновения считалась косвенной из-за их тесной связи с нервными окончаниями. Однако недавние исследования на мышах и других модельных организмах показали, что клетки Меркеля по своей сути преобразуют прикосновение в электрические сигналы, которые передаются в нервную систему. [ 29 ]
Очаговая эпителиальная гиперплазия (болезнь Гека) — бессимптомное доброкачественное опухолевое состояние, характеризующееся множественными папулами от белого до розоватого цвета , которые диффузно возникают в полости рта. [ 32 ] [ 6 ] : 411
Псевдоэпителиоматозная гиперплазия (ПЭГ) — доброкачественное состояние, характеризующееся гиперплазией эпидермиса и эпителия придатков кожи . [ 33 ] с неровными плоскими тяжами, спускающимися в дерму, [ 34 ] и близко имитирует плоскоклеточную карциному (SCC). [ 33 ]
Псевдоэпителиоматозная гиперплазия (ПЭГ), небольшое увеличение, с акантотическим плоским эпителием с неравномерными толстыми пальцевидными разрастаниями в подлежащую дерму.
PEH, большое увеличение, с реактивными плоскоклеточными разрастаниями без значительной цитологической атипии.
^ Jump up to: а б Джеймс В.Д., Бергер Т.Г., Элстон Д.М., Айдемир Э.Х., Одом Р.Б. (2006). Болезни кожи Эндрюса: клиническая дерматология . Сондерс Эльзевир. ISBN 0-7216-2921-0 .
^ Бличманн CW, Серуп Дж (1988). «Оценка влажности кожи. Измерение электропроводности, емкости и трансэпидермальной потери воды». Дерматовенерологический акт . 68 (4): 284–90. doi : 10.2340/0001555568284290 (неактивен 31 января 2024 г.). ПМИД 2459872 . {{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка )
^ Даунинг Д.Т., Стюарт М.Е., Верц П.В., Колтон С.В., Абрахам В., Штраус Дж.С. (март 1987 г.). «Липиды кожи: обновление». Журнал исследовательской дерматологии . 88 (3 дополн.): 2–6 с. doi : 10.1111/1523-1747.ep12468850 (неактивен 01 февраля 2024 г.). ПМИД 2950180 . {{cite journal}}: CS1 maint: DOI неактивен по состоянию на февраль 2024 г. ( ссылка )
^ Бонте Ф., Сонуа А., Пинге П., Мейбек А. (январь 1997 г.). «Существование липидного градиента в верхнем роговом слое и его возможное биологическое значение». Архив дерматологических исследований . 289 (2): 78–82. дои : 10.1007/s004030050158 . ПМИД 9049040 . S2CID 10787600 .
^ Кумар В., Фаусто Н., Аббас А. (2004). Патологическая основа болезней Роббинса и Котрана (7-е изд.). Сондерс. п. 1230. ИСБН 0-7216-0187-1 .
^ Стоун М.С., Рэй Т.Л. (сентябрь 1995 г.). «Акантоз» . ДермПатТутор . Кафедра дерматологии Университета Айовы. Архивировано из оригинала 29 мая 2012 года . Проверено 17 мая 2012 г.
Arc.Ask3.Ru Номер скриншота №: 4e4820ee9b13f34c836082684a2b6a7f__1720966320 URL1:https://arc.ask3.ru/arc/aa/4e/7f/4e4820ee9b13f34c836082684a2b6a7f.html Заголовок, (Title) документа по адресу, URL1: Epidermis - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)






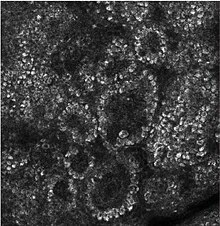






 Псевдоэпителиоматозная гиперплазия (ПЭГ), небольшое увеличение, с акантотическим плоским эпителием с неравномерными толстыми пальцевидными разрастаниями в подлежащую дерму.
Псевдоэпителиоматозная гиперплазия (ПЭГ), небольшое увеличение, с акантотическим плоским эпителием с неравномерными толстыми пальцевидными разрастаниями в подлежащую дерму.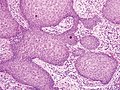 PEH, большое увеличение, с реактивными плоскоклеточными разрастаниями без значительной цитологической атипии.
PEH, большое увеличение, с реактивными плоскоклеточными разрастаниями без значительной цитологической атипии.



 Эпидермис и дерма кожи человека
Эпидермис и дерма кожи человека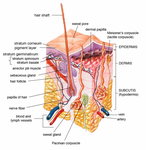 Поперечное сечение всех слоев кожи
Поперечное сечение всех слоев кожи Иллюстрация слоев эпидермиса
Иллюстрация слоев эпидермиса Оптическая когерентная томография кончика пальца
Оптическая когерентная томография кончика пальца


