Полиэлектролитная теория гена
Полиэлектролитная теория гена предполагает, что для того, чтобы линейный генетический биополимер, растворенный в воде, такой как ДНК , претерпел дарвиновскую эволюцию в любой точке Вселенной, он должен быть полиэлектролитом , полимером, содержащим повторяющиеся ионные заряды. [ 1 ] Эти заряды поддерживают одинаковые физические свойства, необходимые для дарвиновской эволюции, независимо от информации, закодированной в генетическом биополимере. [ 1 ] [ 2 ] [ 3 ] ДНК является такой молекулой. Независимо от последовательности нуклеиновой кислоты , отрицательные заряды на ее основной цепи доминируют в физических взаимодействиях молекулы до такой степени, что она сохраняет одинаковые физические свойства, такие как растворимость в воде и структура двойной спирали . [ 1 ]
Полиэлектролитная теория гена была предложена Стивеном А. Беннером и Дэниелом Хаттером в 2002 году. [ 2 ] и в значительной степени остается теоретической основой, которую астробиологи используют для размышлений о том, как можно обнаружить жизнь за пределами Земли. Эта идея позже была связана Беннером [ 3 ] [ 1 ] взгляду Эрвина Шредингера на ген как на « апериодический кристалл ». [ 4 ] создать надежную, универсально обобщенную концепцию генетического биополимера — биополимера, действующего как единица наследования в дарвиновской эволюции.
Беннер и другие, кто основывался на его работе. [ 5 ] предложили методы концентрации и идентификации генетических биополимеров на других планетах и лунах Солнечной системы с помощью электрофореза , который использует электрическое поле для концентрации заряженных соединений. [ 1 ] [ 2 ] [ 3 ]
Хотя мало кто проверял полиэлектролитную теорию гена, в 2019 году лабораторные эксперименты поставили под сомнение универсальность этой идеи. Эта работа позволила создать неэлектролитные полимеры, способные к ограниченной дарвиновской эволюции, но только до длины 72 нуклеотидов. [ 6 ] [ 7 ]
Физическая структура полиэлектролитов
[ редактировать ]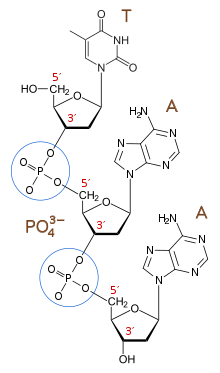
Полиэлектролит заряженными звеньями – это полимер с повторяющимися электростатически . В контексте полиэлектролитной теории гена этот полиэлектролит представляет собой биополимер — полимер, полученный из живой системы — с повторяющейся ионно-заряженной единицей, аналогичный генетическому биополимеру в современной биологии — ДНК. Хотя РНК не действует как генетический биополимерный архив в современной биологии, за исключением некоторых вирусов, таких как коронавирус. [ 8 ] и ВИЧ [ 9 ] — Гипотеза мира РНК предполагает, что РНК, возможно, предшествовала ДНК и стала первым генетическим биополимером жизни. [ 10 ] Строительные блоки нуклеотидов , из которых состоят ДНК и РНК, соединены отрицательно заряженными фосфатными группами. Эти фосфодиэфирные связи создают повторяющиеся отрицательные заряды на основной цепи молекулы, которые придают ДНК и РНК их полиэлектролитную природу. [ 11 ]
Полиэлектролиты в контексте генетических биополимеров
[ редактировать ]Чтобы участвовать в дарвиновской эволюции, которую можно описать как « происхождение с модификациями », [ 12 ] единица наследования должна быть способна к несовершенной репликации, чтобы время от времени создавать новую модифицированную единицу наследования, которая все еще должна быть способна к репликации. Эта несовершенная репликация приводит к вариациям, на которые может воздействовать дарвиновская эволюция. [ 13 ]
Полиэлектролитная теория гена пытается понять современную биологическую единицу наследования — ДНК — на обобщающем уровне. В 2002 году Стивен А. Беннер и Дэниел Хаттер определили, что повторяющиеся заряды в фосфодиэфирных связях ДНК имеют решающее значение для ее функции как генетического биополимера. С помощью полиэлектролитной теории гена они предположили, что повторяющиеся ионные заряды — положительные или отрицательные — являются общим требованием для всех растворенных в воде генетических биополимеров, чтобы пройти дарвиновскую эволюцию в любой точке космоса. [ 2 ]
Эта концепция работает в тандеме с представлением о гене как об «апериодическом кристалле», предложенным Эрвином Шрёдингером в его книге 1944 года « Что такое жизнь? ». [ 4 ] Апериодический кристалл, как его описывает Шредингер, имеет дискретный набор молекулярных строительных блоков, расположенных неповторяющимся образом. ДНК — это апериодический кристалл, состоящий из дискретных нуклеиновых оснований ( A , T , C и G ), которые расположены на основе информации, которую они кодируют, а не в каком-либо повторяющемся формате. Хотя идея «апериодического кристалла» изначально не была связана с полиэлектролитной теорией гена, Беннер в более поздней работе связал их. [ 3 ] [ 1 ]
Полиэлектролиты остаются физически однородными независимо от закодированной информации.
[ редактировать ]
В биохимии структура биомолекулы определяет ее функцию, и поэтому изменения в структуре вызывают изменения в функции. [ 14 ] Чтобы работать как единица наследственности, генетический биополимер должен сохранять форму и, следовательно, физическую и химическую консистенцию, независимо от информации, которую кодирует структура. ДНК является такой молекулой. Независимо от последовательности нуклеиновой кислоты, ДНК сохраняет постоянную структуру двойной спирали и, следовательно, постоянные физические свойства, которые позволяют ей оставаться растворенной в воде и реплицироваться клеточными механизмами. Полиэлектролитная теория гена предполагает, что ДНК может сохранять свою форму независимо от мутаций, поскольку отрицательные заряды фосфатного остова доминируют над физическими взаимодействиями молекулы до такой степени, что изменения в последовательности нуклеиновой кислоты, то есть закодированная информация, не влияют на общее физическое поведение молекулы. [ 2 ]
Например, тимидиновые нуклеотиды (Т) хорошо растворимы в воде, а гуанозиновые нуклеотиды (G) более нерастворимы; однако олигонуклеотид — короткая полинуклеотидная последовательность, состоящая только из тимина и олигонуклеотида, состоящего только из гуанина, имеет одинаковую общую структуру и физические свойства. [ 15 ] Если изменения в последовательности нуклеиновой кислоты , которая кодирует генетическую информацию, изменят физические свойства ДНК, эти изменения могут нарушить механизм репликации ДНК .
Такое физическое единообразие очень редко встречается в природе. Возьмем другой биополимер, например, белки . Последовательность нуклеиновой кислоты в ДНК кодирует последовательность аминокислот , входящих в состав белков. Изменение даже одной аминокислоты в первичной последовательности белка может полностью изменить физические свойства этого белка. Например, признак серповидноклеточной анемии вызван единственной мутацией аденина на тимин в гене гемоглобина , вызывающей переключение с глутаминовой кислоты на валин . [ 16 ] Это полностью меняет трехмерную структуру гемоглобина и, таким образом, меняет физические свойства белка, которые приводят к серповидноклеточному признаку.
Белки чувствительны к изменениям в аминокислотной последовательности, поскольку 20 различных боковых цепей аминокислот образуют связи или частичные связи друг с другом. [ 17 ] Кроме того, основная цепь белка имеет дипольный момент — имеющий частично положительные и частично отрицательные стороны — который может дополнительно создавать взаимодействия внутри молекулы. Эти взаимодействия боковой цепи и основной цепи чувствительны к изменениям в окружающей среде и последовательности аминокислот. Маловероятно, что белок может действовать как генетическая биомолекула, поскольку изменения в последовательности аминокислот приводят к изменениям общей физической структуры и свойств. [ 2 ]
Другой неэлектролитный биополимер столкнулся бы с теми же проблемами, что и белок, действуя как генетическая биомолекула. Изменения физических свойств с изменениями в закодированной информации будут означать, что такая молекула будет с трудом воспроизводиться с определенными последовательностями закодированной информации, поскольку эти последовательности приведут к физическим свойствам, несовместимым с репликацией. Эта проблема означает, что гипотетический ген белка не сможет исследовать все возможные генетические последовательности, поскольку определенные последовательности могут привести к невозможности репликации молекулы из-за физической структуры ее гена, а не из-за приспособленности того, что этот ген кодирует. . [ 2 ]
Беннер и Хаттер первоначально описали это свойство ДНК как «способность выдерживать изменения в конституции без потери свойств, необходимых для репликации» или аббревиатуры COSMIC-LOPER. Эта аббревиатура дает ученым сокращенный способ описания сложной идеи генетического биополимера, обладающего физической однородностью независимо от закодированной информации, которая позволяет его воспроизводить. [ 2 ]
Хотя РНК часто называют генетическим биополимером из-за ее теоретической роли первой единицы наследования жизни (Мира РНК), она не является полностью КОСМИЧЕСКИМ-ЛОПЕРОМ. РНК, особенно последовательности с высоким содержанием гуанина (G), способны сворачиваться и выполнять химические процессы ферментативного типа . [ 2 ] [ 18 ] Сворачивание последовательностей РНК, богатых гуанином, предотвращает способность РНК формировать шаблоны и, следовательно, ее способность реплицироваться в сценарии мира РНК, по той же причине, по которой было бы трудно реплицироваться гену, основанному на белке. [ 2 ]
Повторяющиеся ионные заряды увеличивают растворимость в воде.
[ редактировать ]Повторяющиеся отрицательные заряды увеличивают растворимость ДНК и РНК в воде. Поскольку ионные заряды хорошо растворимы в воде, их расположение на основной цепи молекулы увеличивает растворимость молекулы. [ 19 ] Если бы основная цепь гипотетического генетического биополимера была связана неионным образом, растворимость всей молекулы снизилась бы. [ 20 ] Растворимость важна, потому что для репликации ДНК – или любая другая генетическая биомолекула – должна быть растворимой, чтобы взаимодействовать с репликативным механизмом. [ 11 ]
Повторяющиеся ионные заряды способствуют специфичности спаривания оснований Уотсона-Крика.
[ редактировать ]
Повторяющиеся отрицательные заряды основной цепи ДНК электростатически отталкивают друг друга, предотвращая взаимодействия как внутри, так и между нитями ДНК. Это отталкивание способствует специфическим взаимодействиям вдоль «края» нуклеиновых оснований Уотсона-Крика, способствуя специфичности спаривания оснований Уотсона-Крика : пары A с T и пары C с G. [ 2 ]
Повторяющиеся ионные заряды предотвращают сворачивание.
[ редактировать ]
Повторяющиеся отрицательные заряды на основной цепи удерживают ДНК и многие молекулы РНК от сворачивания и позволяют им действовать как матрицы. В воде молекулы принимают конформацию наиболее энергетически выгодную с наименьшей свободной энергией Гиббса . [ 11 ] Эта конфигурация максимизирует благоприятные взаимодействия ( водородные связи , взаимодействия положительно-отрицательных зарядов, взаимодействия Ван-дер-Ваальса ) и минимизирует неблагоприятные взаимодействия (т. е. гидрофильно-гидрофобные взаимодействия и взаимодействия подобных зарядов). В случае двухцепочечной ДНК и РНК наиболее энергетически выгодной формой является линейная конфигурация двойной спирали, поскольку она максимизирует взаимодействия между парами оснований, а также между отрицательно заряженным остовом и окружающими молекулами воды, одновременно сводя к минимуму взаимодействия между отрицательно заряженными фосфодиэфирными связями позвоночник. [ 11 ] Если двухцепочечная молекула ДНК или РНК сворачивается, она заменяет благоприятные взаимодействия между водой и основной цепью на неблагоприятные взаимодействия между основной цепью. Биополимер без ионно-заряженной основной цепи, подобно белкам, не будет вызывать неблагоприятного взаимодействия между основной цепью во время сворачивания и, таким образом, легко сворачивается и агрегирует . Эта присущая ДНК тенденция к линейности улучшает способность ДНК выступать в качестве матрицы для репликации. [ 2 ] потому что свернутые и агрегированные конформации недоступны механизму репликации.
Лабораторные эксперименты
[ редактировать ]Лабораторные эксперименты, проведенные с неэлектролитными аналогами ДНК и РНК, первоначально вдохновили Беннера и Хаттона на публикацию полиэлектролитной теории гена. [ 3 ] В конце 80-х и 90-х годах ученые разработали синтетические ДНК-подобные молекулы для связывания и подавления нежелательных генных продуктов мРНК в качестве способа лечения заболеваний. [ 2 ] [ 21 ] В рамках этого исследовательского исследования исследователи разработали множество неэлектролитных аналогов РНК и ДНК, которые смогут проникать через клеточную мембрану , на что ДНК и РНК неспособны из-за их заряженных остовов. Один из этих аналогов заменил сульфоном природную фосфодиэфирную связь (PO₂⁻) (SO₂). Хотя первоначальные эксперименты показали, что аналог сульфона имеет очень схожие свойства с ДНК в виде димера (два связанных вместе нуклеотида), когда были синтезированы более длинные аналоги сульфона, они сворачивались, теряли специфичность пары оснований Уотсона-Крика и имели драматические изменения в физических свойствах из-за небольшие изменения в последовательности нуклеиновых кислот. [ 2 ] Снижение качества признаков, которые делают ДНК хорошей генетической молекулой, наблюдалось для всех неионных линкеров, протестированных по состоянию на 2002 год. [ 2 ]
Ближайшим неэлектролитным аналогом для поддержания свойств ДНК был аналог нуклеиновой кислоты с полиамидной связью (ПНК), который заменил фосфодиэфирную связь ДНК незаряженной N-(2-аминоэтил)глициновой связью. Даже Беннер и Хаттер задавались вопросом, может ли ПНА опровергнуть их гипотезу полиэлектролита; однако, хотя ПНА сохраняла качества ДНК до длины 20 нуклеотидов, за пределами этой длины молекулы начали терять специфичность пар оснований Уотсона-Крика, агрегировались и становились чувствительными к изменениям последовательности нуклеиновых кислот. [ 2 ]
Лабораторные эксперименты, бросающие вызов полиэлектролитной теории гена
[ редактировать ]В 2019 году группа под руководством Филиппа Холлигера в Кембридже, Англия, разработала неэлектролитные аналоги ДНК P-алкилфосфонатных нуклеиновых кислот (phNA), которые были способны подвергаться матричному синтезу и направленной эволюции . [ 22 ] Аналоги phNA заменили заряженный кислород на фосфатном остове ДНК незаряженной метильной или этильной группой . В то время как было показано, что другие аналоги ДНК подвергаются шаблонному синтезу и направленной эволюции, это открытие стало первым случаем, когда неэлектролитный аналог ДНК обладал этими свойствами, и первым случаем, когда полиэлектролитная теория гена была подвергнута экспериментальному сомнению. [ 23 ] Однако матричный синтез phNA осуществлялся только до длины 72 нуклеотидов. [ 22 ] Это примерно соответствует длине самого короткого природного гена, тРНК. [ 24 ] но примерно на порядок короче генома самого маленького свободноживущего организма. [ 25 ] Для справки, длина генома человека составляет 3,05×10⁹ пар оснований. [ 26 ]
Как «агностическая биосигнатура»
[ редактировать ]С момента своего создания полиэлектролитная теория гена рассматривалась в контексте поиска жизни во Вселенной. Эта теория в сочетании с представлением Шрёдингера о гене как об апериодическом кристалле обеспечивает так называемую «агностическую биосигнатуру ». [ 1 ] признак жизни, не предполагающий никакой биохимии. [ 27 ] Другими словами, обобщенный взгляд на жизнь должен иметь место в любой точке Вселенной. [ 1 ]
Поскольку теоретические генетические полиэлектролитные биомолекулы могут быть заряжены как положительно, так и отрицательно, как в случае с ДНК и РНК, их можно концентрировать в воде с помощью электрического поля с помощью электрофореза или электродиализа . Это гипотетическое устройство для концентрации было названо агностическим устройством поиска жизни. [ 1 ] Подобно тому, как электрофорез работает для разделения молекул ДНК, отрицательно заряженные молекулы, такие как ДНК или РНК, будут притягиваться к положительно заряженному аноду, а положительно заряженные генетические биомолекулы будут притягиваться к отрицательно заряженному катоду. [ 15 ]
Беннер предлагает протестировать молекулы на однородность размера и формы после того, как биомолекула полиэлектролита будет сконцентрирована. Кроме того, молекулы должны быть проверены на предмет использования ограниченного числа строительных блоков, расположенных неповторяющимся образом, апериодической кристаллической структуры. [ 1 ] Беннер предположил, что это можно сделать с помощью матричной лазерной десорбционной ионизации (MALDI) высокого разрешения с орбитальной ловушкой в сочетании с масс-спектрометром . [ 28 ] Другой предложенный подход заключался в использовании технологии секвенирования нанопор , хотя остаются вопросы о том, повлияет ли солнечное излучение , испытываемое во время транспортировки и на месте, на функциональность устройства. [ 5 ] Хотя космические агентства еще не использовали ни одну из этих предложенных систем для обнаружения жизни, в будущем они могут быть использованы на Марсе. [ 28 ] Энцелад, [ 1 ] и Европа. [ 5 ]
Несмотря на то, что полиэлектролитная теория гена и апериодический кристаллический взгляд на ген описываются как агностические биосигнатуры, эти теории ориентированы на терра- или земную жизнь. Неизвестно, какой может быть жизнь в другом мире; Хотя часто утверждают, что жизнь любого вида нуждается в биомолекулах и воде, это может быть неправдой. [ 19 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к Беннер, Стивен А. (2017). «Обнаружение дарвинизма на основе молекул в шлейфах Энцелада, лунах Юпитера и других планетарных водных лагунах» . Астробиология . 17 (9): 840–851. Бибкод : 2017AsBio..17..840B . дои : 10.1089/ast.2016.1611 . ISSN 1531-1074 . ПМК 5610385 . ПМИД 28665680 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Беннер, Стивен А.; Хаттер, Дэниел (01 февраля 2002 г.). «Фосфаты, ДНК и поиск внеземной жизни: модель второго поколения генетических молекул» . Биоорганическая химия . 30 (1): 62–80. дои : 10.1006/bioo.2001.1232 . ISSN 0045-2068 . ПМИД 11955003 .
- ^ Jump up to: а б с д и Беннер, Стивен А. (27 февраля 2023 г.). «Переосмысление нуклеиновых кислот от их происхождения до их применения» . Философские труды Королевского общества B: Биологические науки . 378 (1871). дои : 10.1098/rstb.2022.0027 . ISSN 0962-8436 . ПМЦ 9835595 . ПМИД 36633284 .
- ^ Jump up to: а б Шредингер, Эрвин (1944). Что такое Жизнь? Физический аспект живой клетки . Великобритания: Кембриджский университет. ISBN 0-521-42708-8 .
- ^ Jump up to: а б с Саттон, Марк А.; Бертон, Аарон С.; Зайкова, Елена; Саттон, Райан Э.; Бринкерхофф, Уильям Б.; Бевилаква, Джули Г.; Венг, Маргарет М.; Мама, Майкл Дж.; Джонсон, Сара Стюарт (29 марта 2019 г.). «Радиационная устойчивость технологии секвенирования нанопор для обнаружения жизни на Марсе и Европе» . Научные отчеты . 9 (1): 5370. Бибкод : 2019НатСР...9.5370С . дои : 10.1038/s41598-019-41488-4 . ISSN 2045-2322 . ПМК 6441015 . ПМИД 30926841 .
- ^ Браун, Аша; Браун, Том (2019). «Ограничение своего негатива» . Природная химия . 11 (6): 501–503. Бибкод : 2019НатЧ..11..501Б . дои : 10.1038/s41557-019-0274-1 . ISSN 1755-4349 . ПМИД 31123348 . S2CID 163166821 .
- ^ Арангунди-Франклин, Себастьян; Тейлор, Александр И.; Поребски, Бенджамин Т.; Генна, Вито; Пик-Жевать, Шить; Вайсман, Александра; Вудгейт, Роджер; Ороско, Модесто; Холлигер, Филипп (2019). «Синтетический генетический полимер с незаряженной основной цепью на основе алкилфосфонатных нуклеиновых кислот» . Природная химия . 11 (6): 533–542. Бибкод : 2019НатЧ..11..533А . дои : 10.1038/s41557-019-0255-4 . ISSN 1755-4349 . ПМК 6542681 . ПМИД 31011171 .
- ^ Брант, Айслан Кастро; Тянь, Вэй; Маерчак Владимир; Ян, Вэй; Чжэн, Чжи-Мин (2021). «SARS-CoV-2: от открытия до структуры генома, транскрипции и репликации» . Клетка и биологические науки . 11 (1): 136. дои : 10.1186/s13578-021-00643-z . ПМЦ 8287290 . ПМИД 34281608 .
- ^ Харвиг, Алекс; Дас, Аце Т.; Берхаут, Бен (2015). «РНК ВИЧ-1: смысловые и антисмысловые, большие мРНК и малые миРНК и микроРНК» . Современное мнение о ВИЧ и СПИДе . 10 (2): 103–109. дои : 10.1097/COH.0000000000000135 . ISSN 1746-630X . ПМИД 25565176 . S2CID 34885126 .
- ^ Мир РНК
- ^ Jump up to: а б с д Нельсон, Дэвид Л.; Кокс, Майкл М.; Нельсон, Дэвид Л. (2013). Ленинджер, Альберт Л. (ред.). Ленингерские принципы биохимии (Шестое изд.). Бейзингсток: Высшее образование Макмиллана. ISBN 978-1-4292-3414-6 .
- ^ Бромхэм, Линделл (21 января 2016 г.), «Отбор» , «Введение в молекулярную эволюцию и филогенетику» , Oxford University Press, doi : 10.1093/hesc/9780198736363.003.0007 , ISBN 978-0-19-873636-3 , получено 9 декабря 2023 г.
- ^ Грегори, Т. Райан (2009). «Понимание естественного отбора: основные понятия и распространенные заблуждения» . Эволюция: образование и информационно-пропагандистская деятельность . 2 (2): 156–175. дои : 10.1007/s12052-009-0128-1 . ISSN 1936-6434 . S2CID 4508223 .
- ^ «2.3: Структура и функции – Белки I» . Свободные тексты по биологии . 21 января 2017 г. Проверено 5 декабря 2023 г.
- ^ Jump up to: а б Беннер, Стивен А. «Применение полиэлектролитной теории гена в агностических поисках марсианской жизни - первозданная сенсация» . Проверено 5 декабря 2023 г.
- ^ «Генетическая мутация | Изучайте науку в Scitable» . www.nature.com . Проверено 7 декабря 2023 г.
- ^ «Сворачивание белка» . Химия LibreTexts . 02.10.2013 . Проверено 7 декабря 2023 г.
- ^ Талини, Джулия; Бранчамор, Серхио; Галлори, Энцо (2011). «Рибозимы: гибкие молекулярные устройства в работе» . Биохимия 93 (11): 1998–2005. дои : 10.1016/j.biochi.2011.06.026 . ПМИД 21740954 .
- ^ Jump up to: а б Беннер, Стивен А. (2010). «Определение жизни» . Астробиология . 10 (10): 1021–1030. Бибкод : 2010AsBio..10.1021B . дои : 10.1089/ast.2010.0524 . ISSN 1531-1074 . ПМК 3005285 . ПМИД 21162682 .
- ^ «4.5: Растворимость ионных соединений» . Химия LibreTexts . 20 января 2022 г. Проверено 7 декабря 2023 г.
- ^ Крук, Стэнли Т. (2000), «Прогресс в антисмысловой технологии: конец начала», Антисмысловая технология, часть A: Общие методы, методы доставки и исследования РНК , Методы в энзимологии, том. 313, Elsevier, стр. 3–45, номер документа : 10.1016/s0076-6879(00)13003-4 , ISBN. 978-0-12-182214-9 , PMID 10595347 , получено 7 декабря 2023 г.
- ^ Jump up to: а б Арангунди-Франклин, Себастьян; Тейлор, Александр И.; Поребски, Бенджамин Т.; Генна, Вито; Пик-Жевать, Шить; Вайсман, Александра; Вудгейт, Роджер; Ороско, Модесто; Холлигер, Филипп (2019). «Синтетический генетический полимер с незаряженной основной цепью на основе алкилфосфонатных нуклеиновых кислот» . Природная химия . 11 (6): 533–542. Бибкод : 2019НатЧ..11..533А . дои : 10.1038/s41557-019-0255-4 . ISSN 1755-4330 . ПМК 6542681 . ПМИД 31011171 .
- ^ Браун, Аша; Браун, Том (2019). «Ограничение своего негатива» . Природная химия . 11 (6): 501–503. Бибкод : 2019НатЧ..11..501Б . дои : 10.1038/s41557-019-0274-1 . ISSN 1755-4330 . ПМИД 31123348 . S2CID 163166821 .
- ^ Кран, Натали; Фишер, Джонатан Т.; Зёлль, Дитер (2020). «Естественно встречающиеся тРНК с неканонической структурой» . Границы микробиологии . 11 . дои : 10.3389/fmicb.2020.596914 . ISSN 1664-302X . ПМК 7609411 . ПМИД 33193279 .
- ^ Джованнони, Стивен Дж.; Трипп, Х. Джеймс; Гиван, Скотт; Подар, Мирча; Вергин, Кевин Л.; Баптиста, Дэймон; Биббс, Лиза; Идс, Джонатан; Ричардсон, Тоби Х.; Нордевир, Мишель; Раппе, Майкл С.; Шорт, Джей М.; Кэррингтон, Джеймс С.; Матур, Эрик Дж. (19 августа 2005 г.). «Оптимизация генома космополитической океанической бактерии» . Наука . 309 (5738): 1242–1245. Бибкод : 2005Sci...309.1242G . дои : 10.1126/science.1114057 . ISSN 0036-8075 . ПМИД 16109880 . S2CID 16221415 .
- ^ Нурк, Сергей; Корень, Сергей; Ри, уголь; Раутиайнен, Микко; Бзикадзе Андрей Владимирович; Михеенко Алла; Фоллгер, Митчелл Р.; Альтемос, Николас; Уральский, Лев; Гершман, Ариэль; Аганезов Сергей; Хойт, Саванна Дж.; Диканс, Марк; Логсдон, Гленнис А.; Алонге, Майкл (2022). «Полная последовательность человеческого генома» . Наука . 376 (6588): 44–53. Бибкод : 2022Sci...376...44N . дои : 10.1126/science.abj6987 . ISSN 0036-8075 . ПМЦ 9186530 . ПМИД 35357919 .
- ^ «Астробиология НАСА» . astrobiology.nasa.gov . Проверено 7 декабря 2023 г.
- ^ Jump up to: а б Шпачек, Ян; Беннер, Стивен А. (01 октября 2022 г.). «Агностический искатель жизни (ALF) для крупномасштабного обследования марсианской жизни во время дозаправки на месте» . Астробиология . 22 (10): 1255–1263. Бибкод : 2022AsBio..22.1255S . дои : 10.1089/ast.2021.0070 . ISSN 1531-1074 . ПМИД 35796703 . S2CID 250336451 .