Телитоки

Телитоки (от греческого θῆλυς thēlys «женщина» и τόκος tókos «рождение») представляет собой тип партеногенеза и представляет собой отсутствие спаривания и последующего производства всех диплоидных потомков женского пола, как, например, у тли . По данным исследования 1984 года , телитокозный партеногенез редко встречается среди животных и зарегистрирован примерно у 1500 видов , примерно у 1 из 1000 описанных видов животных. [ 1 ] Это чаще встречается у беспозвоночных , таких как членистоногие , но может встречаться и у позвоночных , включая саламандр , рыб и рептилий, таких как некоторые ящерицы с хлыстовым хвостом .
Телитокия может возникать по разным механизмам, каждый из которых по-разному влияет на уровень гомозиготности . Он встречается в нескольких группах перепончатокрылых , включая Apidae , Aphelinidae , Cynipidae , Formicidae , Ichneumonidae и Tenthredinidae . [ 2 ] У перепончатокрылых он может быть индуцирован бактериями Wolbachia и Cardinium . [ 3 ]
Преимущества телитоки
[ редактировать ]Виды могут получить несколько преимуществ, используя эту форму системы спаривания. Телитокия позволяет самкам передавать генотипы, обеспечивающие успех в данной конкретной среде; наличие только дочерей увеличивает продуктивность вида, а энергия, которая в противном случае была бы затрачена на поиск или привлечение партнера, может быть непосредственно инвестирована в воспроизводство. [ 4 ]
Телитокия может возникать естественным путем или может быть вызвана учеными в лабораторных условиях. [ 5 ] У некоторых видов телитоки также может возникать в результате слияния двух женских гамет. [ 6 ]
Виды телитокии
[ редактировать ]Факультативная телитокия означает способность человека к размножению половым или бесполым путем в зависимости от условий окружающей среды. Например, мелкозубая рыба-пила в популяциях Флориды может быть факультативно телитокозной, что означает, что они будут размножаться половым путем при благоприятных условиях, но переключаются на телитоки, когда ресурсов и партнеров становится недостаточно. [ 7 ]
Случайная телитокия возникает, когда женский организм производит потомство бесполым путем из-за отсутствия или неудачного оплодотворения самца. Это может произойти у видов, которые обычно размножаются половым путем, но не могут найти себе пару, или у видов, у которых спаривание не удается из-за физических или поведенческих барьеров. Хотя случайная телитокия может обеспечить краткосрочное репродуктивное решение в отсутствие партнера, она обычно не является устойчивой в долгосрочной перспективе из-за потери генетического разнообразия. [ 8 ]
Циклическая телитоки - это форма телитоки, при которой организмы чередуют половое и бесполое размножение в регулярном цикле. Этот тип размножения наблюдается у цинипидных желчных ос , у которых половое размножение происходит в чередующихся поколениях. Бесполое размножение, происходящее между этими половыми поколениями, обычно облегчается наличием определенных факторов окружающей среды, таких как температура или фотопериод. Считается, что генетическое разнообразие, создаваемое половым размножением этих организмов, играет важную роль в их способности адаптироваться к изменяющимся условиям окружающей среды. [ 9 ]
Облигатная телитоки относится к форме бесполого размножения, при которой человек не может размножаться половым путем и должен полагаться на бесполое размножение для размножения. Виды, которые являются облигатно телитокозными, не обладают генетическими или физиологическими механизмами, необходимыми для производства самцов, и поэтому полагаются исключительно на потомство женского пола для продолжения своей линии. Примеры облигатно телитокозных видов включают некоторых представителей муравьев cerapachys и некоторых видов ящериц с хлыстовым хвостом . [ 10 ]
Арренотокия и телитоки у перепончатокрылых
[ редактировать ]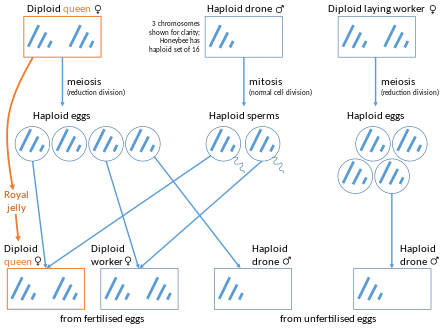
Перепончатокрылые ( муравьи , пчелы , осы и пилильщики ) имеют гаплодиплоидную систему определения пола . Они производят гаплоидных самцов из неоплодотворенных яиц ( арренотокии ), форма партеногенеза . Однако у некоторых социальных перепончатокрылых королевы или рабочие способны производить диплоидное потомство женского пола с помощью телитоки. [ 11 ] Произведенные дочери могут быть или не быть полными клонами своей матери в зависимости от типа происходящего партеногенеза. [ 12 ] [ 13 ] Потомство может развиться как в королев, так и в рабочих особей. Примеры таких видов включают капскую пчелу, Apis mellifera capensis , Mycocepurus smithii и клонального муравья-налётчика Ooceraea biroi .
Автомикс
[ редактировать ]
Аутомиксис является формой телитокии. При аутомиктическом происходит партеногенезе мейоз и диплоидия восстанавливается за счет слияния несестринских ядер первого деления (центральное слияние) или сестринских ядер второго деления (терминальное слияние). [ 14 ] (см. схему).
С центральным слиянием
[ редактировать ]Аутомиксис с центральным слиянием имеет тенденцию поддерживать гетерозиготность при передаче генома от матери к дочери. Эта форма аутомиксиса наблюдалась у нескольких видов муравьев, включая пустынного муравья Cataglyphis курсора . [ 11 ] клональный муравей-налетчик Cerapachys biroi , [ 15 ] хищный муравей Platythyrea punctata , [ 14 ] и электрический муравей (маленький огненный муравей) Wasmannia auropunctata . [ 16 ] Аутомиксис с центральным слиянием встречается также у капской медоносной пчелы Apis mellifera capensis . [ 13 ] артемия Artemia parthenogenetica , [ 17 ] и термит Embiratermes neotenicus . [ 18 ]
Ооциты , подвергающиеся аутомиксису с центральным слиянием, часто демонстрируют пониженную скорость кроссоверной рекомбинации . Низкая скорость рекомбинации в аутомиктических ооцитах благоприятствует поддержанию гетерозиготности и лишь медленному переходу от гетерозиготности к гомозиготности на протяжении последующих поколений. Это позволяет избежать немедленной инбредной депрессии . Виды, демонстрирующие центральное слияние с пониженной рекомбинацией, включают муравьев P. punctata. [ 14 ] и W. auropunctata , [ 16 ] артемия A. parthenogenetica , [ 17 ] и медоносная пчела А. м. капенсис . [ 13 ] В А. М. capensis , скорость рекомбинации во время мейоза, связанного с телитокозным партеногенезом, снижается более чем в 10 раз. [ 13 ] У W. auropunctata снижение происходит в 45 раз. [ 16 ]
Одиночные колонии маток узкоголового муравья Formica exsecta представляют собой наглядный пример возможных вредных последствий повышенной гомозиготности. У этого муравья уровень гомозиготности королевы отрицательно связан с возрастом колонии. [ 19 ] Снижение выживаемости колоний, по-видимому, связано с уменьшением продолжительности жизни маток в результате гомозиготности маток и проявления вредных рецессивных мутаций ( инбредная депрессия ).
С терминальным слиянием
[ редактировать ]Аутомиксис с терминальным слиянием имеет тенденцию способствовать гомозиготности при передаче генома от матери к дочери. Такая форма аутомиксиса наблюдалась у водяной блохи Daphnia magna. [ 20 ] и колумбийский радужный удав Epicrates maurus . [ 21 ] Партеногенез у E. maurus — лишь третий генетически подтвержденный случай последовательных девственных рождений жизнеспособного потомства от одной самки в любой линии позвоночных. [ 21 ] Однако выживаемость потомства в двух последовательных пометах была плохой, что позволяет предположить, что аутомиксис с терминальным слиянием приводит к гомозиготности и экспрессии вредных рецессивных аллелей ( инбредная депрессия ).
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Уайт, Майкл Джей Ди (1984). «Хромосомные механизмы размножения животных». Боллетино ди Зоологии . 51 (1–2): 1–23. дои : 10.1080/11250008409439455 . ISSN 0373-4137 .
- ^ Суомалайнен, Эско; Ансси Саура; Юхани Локки (31 августа 1987 г.). Цитология и эволюция в партеногенезе . ЦРК Пресс. п.п. 29–31, 51. ISBN. 978-0-8493-5981-1 .
- ^ Чон, Дж; Р. Стаутхамер (3 ноября 2004 г.). «Генетика функциональной девственности самок в партеногенезе - зараженной Wolbachia паразитоидной осе Telenomus nawai (Hymenoptera: Scelionidae)» . Наследственность . 94 (4): 402–407. дои : 10.1038/sj.hdy.6800617 . ISSN 0018-067X . ПМИД 15523503 .
- ^ Дж., Белл, Уильям (2007). Тараканы: экология, поведение и естественная история . Издательство Университета Джонса Хопкинса. ISBN 978-1-4356-9271-8 . OCLC 646769575 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Чжу, Дао-Хун; Он, И-Юань; Фань, Юн-Шэн; Ма, Мин-Ён; Пэн, Де-Лян (сентябрь 2007 г.). «Отрицательные доказательства индукции партеногенеза Wolbachia у вида желчной осы Dryocosmus kuriphilus» . Энтомология экспериментальная и прикладная . 124 (3): 279–284. дои : 10.1111/j.1570-7458.2007.00578.x . ISSN 0013-8703 . S2CID 84922476 .
- ^ Бенджамин П. Олдройд; Майкл Х. Олсопп; Розалин С. Глоаг; Джулианна Лим; Линдон А. Джордан; Мадлен Бикман (1 сентября 2008 г.). «Телитокозный партеногенез у непарных королев медоносных пчел (Apis mellifera capensis): центральное слияние и высокие скорости рекомбинации» . Генетика . 180 (1): 359–366. doi : 10.1534/genetics.108.090415 . ПМЦ 2535687 . ПМИД 18716331 .
- ^ Филдс, Эндрю Т.; Фельдхайм, Кевин А.; Пулакис, Грегг Р.; Чепмен, Демиан Д. (01 июня 2015 г.). «Факультативный партеногенез у находящихся под угрозой исчезновения диких позвоночных» . Современная биология . 25 (11): Р446–Р447. дои : 10.1016/j.cub.2015.04.018 . ISSN 0960-9822 . ПМИД 26035783 .
- ^ Пардо, MC; Лопес-Леон, доктор медицины; Кабреро, Дж; Камачо, JPM (ноябрь 1995 г.). «Цитологический анализ и анализ развития тихопартеногенеза Locusta migratoria» . Наследственность . 75 (5): 485–494. дои : 10.1038/hdy.1995.165 . ISSN 0018-067X .
- ^ «Поправка к: «Замыканию жизненного цикла» Andricus quercuslanigera (Hymenoptera: Cynipidae)» . Анналы Энтомологического общества Америки . 116 (1): 72–73. 22 октября 2022 г. дои : 10.1093/aesa/saac020 . ISSN 0013-8746 .
- ^ Куэльяр, Орландо (1968). «Дополнительные доказательства истинного партеногенеза у ящериц рода Cnemidophorus» . Герпетологика . 24 (2): 146–150. ISSN 0018-0831 . JSTOR 3891303 .
- ^ Jump up to: а б Пирси, М. (2004). «Условное использование пола и партеногенеза для производства рабочих и маток у муравьев» (PDF) . Наука . 306 (5702): 1780–1783. Бибкод : 2004Sci...306.1780P . дои : 10.1126/science.1105453 . ПМИД 15576621 . S2CID 37558595 .
- ^ Фурнье, Дени; Эступ, Арно; Оривел, Жером; Фуко, Жюльен; Журдан, Эрве; Бретон, Жюльен Ле; Келлер, Лоран (2005). «Клональное размножение самцов и самок у маленького огненного муравья» (PDF) . Природа . 435 (7046): 1230–1234. Бибкод : 2005Natur.435.1230F . дои : 10.1038/nature03705 . ПМИД 15988525 . S2CID 1188960 .
- ^ Jump up to: а б с д Бодри Э., Крайгер П., Оллсопп М., Кенигер Н., Вотрен Д., Мугель Ф., Корнуэ Ж.М., Солиньяк М. (2004). «Сканирование всего генома у телитокозных откладывателей капской медоносной пчелы ( Apis mellifera capensis ): центральное слияние, снижение скорости рекомбинации и картирование центромер с использованием полутетрадного анализа» . Генетика . 167 (1): 243–252. дои : 10.1534/генетика.167.1.243 . ПМЦ 1470879 . ПМИД 15166151 .
- ^ Jump up to: а б с Келлнер, Катрин; Хайнце, Юрген (2010). «Механизм факультативного партеногенеза у муравья Platythyrea punctata ». Эволюционная экология . 25 (1): 77–89. дои : 10.1007/s10682-010-9382-5 . S2CID 24645055 .
- ^ Оксли П.Р., Джи Л., Феттер-Прунеда И., Маккензи С.К., Ли С., Ху Х., Чжан Г., Кронауер DJ (2014). «Геном клонального муравья-налётчика Cerapachys biroi » . Современная биология . 24 (4): 451–8. дои : 10.1016/j.cub.2014.01.018 . ПМК 3961065 . ПМИД 24508170 .
- ^ Jump up to: а б с Рей О, Луазо А, Факон Б, Фуко Дж, Оривел Дж, Корнуэ ЖМ, Роберт С, Добиньи Дж, Делаби Дж. Х., Мариано Кдос С., Эступ А (2011). «Мейотическая рекомбинация резко снизилась у телитокозных королев маленьких огненных муравьев и их рабочих, полученных половым путем» . Молекулярная биология и эволюция . 28 (9): 2591–601. дои : 10.1093/molbev/msr082 . ПМИД 21459760 .
- ^ Jump up to: а б Нуге О, Роде Н.О., Джаббур-Захаб Р., Сегар А., Шевен Л.М., Хааг Ч.Р., Ленорман Т. (2015). «Автомиксис в Артемии : решение векового спора» . Журнал эволюционной биологии . 28 (12): 2337–48. дои : 10.1111/jeb.12757 . ПМИД 26356354 .
- ^ Фугейроллас Р., Долейшова К., Силлам-Дюссес Д., Рой В., Пото С., Ханус Р., Ройзен Ю. (2015). «Бесполая смена королев у высших термитов Embiratermes neotenicus » . Труды Лондонского королевского общества B: Биологические науки . 282 (1809): 20150260. doi : 10.1098/rspb.2015.0260 . ПМК 4590441 . ПМИД 26019158 .
- ^ Хааг-Лиаутард С., Витикайнен Э., Келлер Л., Сундстрем Л. (2009). «Приспособленность и уровень гомозиготности общественного насекомого» (PDF) . Журнал эволюционной биологии . 22 (1): 134–42. дои : 10.1111/j.1420-9101.2008.01635.x . ПМИД 19127611 .
- ^ Свендсен Н., Рейссер С.М., Дукич М., Тюилье В., Сегард А., Лиотард-Хааг С., Фазель Д., Хюрлиманн Е., Ленорман Т., Галимов Ю., Хааг С.Р. (2015). «Раскрытие загадочной асексуальности у Daphnia magna с помощью RAD-секвенирования» . Генетика . 201 (3): 1143–55. дои : 10.1534/genetics.115.179879 . ПМЦ 4649641 . ПМИД 26341660 .
- ^ Jump up to: а б Бут В., Миллион Л., Рейнольдс Р.Г., Бургхардт Г.М., Варго Э.Л., Шал С. , Цика А.С., Шуэтт Г.В. (2011). «Последовательные непорочные роды в новом мире — змея-боа, колумбийский радужный удав, Epicrates maurus » . Журнал наследственности . 102 (6): 759–63. doi : 10.1093/jhered/esr080 . ПМИД 21868391 .
| Темы |  | |
|---|---|---|
| Группы | ||
| В культуре | ||
| Пионеры, работы |
| |