Развитие растений
Важными структурами в развитии растений являются почки , побеги , корни , листья и цветы ; Растения производят эти ткани и структуры на протяжении всей своей жизни из меристем [ 1 ] расположен на кончиках органов или между зрелыми тканями. Таким образом, живое растение всегда имеет эмбриональные ткани. Напротив, эмбрион животных очень рано произведет все части тела, которые он когда -либо будет иметь в своей жизни. Когда животное рождается (или выводит из яйца), оно имеет все части тела и с этого момента будет только больше и более зрелым. Однако как растения, так и животные проходят через филотипическую стадию , которая эволюционировала независимо [ 2 ] И это приводит к ограничению ограничения развития, ограничивающей морфологическую диверсификацию. [ 3 ] [ 4 ] [ 5 ] [ 6 ]
По словам физиолога растений А. Карла Леопольда , свойства организации, наблюдаемые на заводе, представляют собой возникающие свойства , которые являются больше, чем сумма отдельных частей. «Сборка этих тканей и функций в интегрированный многоклеточный организм дает не только характеристики отдельных частей и процессов, но и довольно новый набор характеристик, которые не были бы предсказуемыми на основе изучения отдельных частей». [ 7 ]
Рост
[ редактировать ]начинается Сосудистое растение с одноклеточной зиготы , образованной оплодотворением яичной клетки сперматозоидами. С этого момента он начинает делиться, чтобы сформировать эмбрион растения в процессе эмбриогенеза . Поскольку это происходит, полученные клетки будут организоваться так, чтобы один конец стал первым корнем, в то время как другой конец образует кончик побега. У семенных растений у эмбриона будет развиваться один или несколько «семян листьев» ( семядоли ). К концу эмбриогенеза молодое растение будет иметь все части, необходимые для начала своей жизни. [ Цитация необходима ]
Как только эмбрион прорастает от своего семян или родительского растения, он начинает производить дополнительные органы (листья, стебли и корни) в процессе органогенеза . Новые корни растут от корневых меристем, расположенных на кончике корня, а новые стебли и листья растут от меристем, расположенных на кончике побега. [ 8 ] Разветвление возникает, когда небольшие сцепления клеток, оставленных меристемой, и которые еще не подвергались клеточной дифференцировке с образованием специализированной ткани, начинают расти в качестве кончика нового корня или побега. Рост от любой такой меристемы на кончике корня или побега называется первичным ростом и приводит к удлинению этого корня или побега. Вторичный рост приводит к расширению корня или побега от подразделений клеток в камбия . [ 9 ]
В дополнение к росту делением клеток , растение может расти путем удлинения клеток . Это происходит, когда отдельные клетки или группы клеток растут дольше. Не все растительные клетки растут до той же длины. Когда клетки на одной стороне ствола растут дольше и быстрее, чем клетки с другой стороны, в результате стебель склоняется в сторону более медленных растущих клеток. Этот направленный рост может происходить через реакцию растения на конкретный стимул, такой как свет ( фототропизм ), гравитация ( гравитропизм ), вода, ( гидротропизм ) и физический контакт ( тигмотропизм ). [ Цитация необходима ]
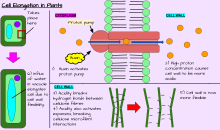

Рост и развитие растений опосредована специфическими гормонами растений и регуляторами роста растений (PGR) (Ross et al. 1983). [ 10 ] Уровень эндогенного гормона влияет на возраст растений, холодная устойчивость, покоя и другие метаболические условия; фотопериод, засуха, температура и другие внешние условия окружающей среды; и экзогенные источники PGR, например, применяемые извне и ризосферное происхождение.
Морфологические различия во время роста
[ редактировать ]Растения демонстрируют естественные различия в их форме и структуре. В то время как все организмы варьируются от индивидуального к индивидуумам, растения демонстрируют дополнительный тип вариаций. Внутри одного человека повторяются части, которые могут отличаться по форме и структуре от других подобных частей. Этот вариант легче всего увидеть в листьях растения, хотя другие органы, такие как стебли и цветы, могут показывать одинаковые вариации. Существуют три основные причины этого изменения: позиционные эффекты, воздействие на окружающую среду и несоответствие. [ Цитация необходима ]

Существует различия между частями зрелого растения, возникающего в результате относительного положения, где производится орган. Например, вдоль новой ветви листья могут варьироваться по постоянному рисунку вдоль ветви. Форма листьев, полученных вблизи основания ветви, отличается от листьев, производимых на кончике растения, и это различие согласуется от ветви к ветвью на данном растении и у данного вида.
То, как новые структуры созревают по мере их производства, может зависеть от точки в жизни растений, когда они начинают развиваться, а также в окружающей среде, в которой разоблачены структуры. Температура обладает множеством воздействий на растения в зависимости от множества факторов, включая размер и состояние растения, а также температуру и продолжительность воздействия. Чем меньше и сочное растение, тем больше восприимчивость к повреждению или смерти от температур, которые слишком высокие или слишком низкие. Температура влияет на скорость биохимических и физиологических процессов, скорости, как правило, (в пределах) увеличиваются с температурой.
Ювенильность или гетеробластия - это когда органы и ткани, полученные молодым растением, таким как рассада , часто отличаются от тех, которые производятся одним и тем же растением, когда оно старше. Например, молодые деревья будут производить более длинные, более скудные ветви, которые растут больше, чем ветви, которые они будут производить в качестве полностью выращенного дерева. Кроме того, листья, полученные во время раннего роста, имеют тенденцию быть больше, тоньше и нерегулярны, чем листья на растении для взрослых. Образцы ювенильных растений могут выглядеть настолько отличающимися от взрослых растений из того же вида, что насекомые с яйцами не распознают растение как пищу для своих молодых. Переход от ранних и поздних форм роста иногда называют изменением вегетативной фазы . [ 11 ]
Дятельные структуры
[ редактировать ]Растительные сооружения, в том числе корни, почки и побеги, которые развиваются в необычных местах, называются случайными . Такие структуры распространены в сосудистых растениях. [ Цитация необходима ]
Авентаризные корни и почки обычно развиваются вблизи существующих сосудистых тканей, так что они могут соединиться с ксилемой и флоэмой . Однако точное местоположение сильно варьируется. В молодых стебли случайные корни часто образуются из паренхимы между сосудистыми пучками . В стебли со вторичным ростом, случайные корни часто возникают в паренхиме флоэмы возле камбия сосудов . В Stem черенки, случайные корни иногда также возникают в мозоль -клетках, которые образуются на поверхности среза. Листовые черенки из крассовой формы образуют эквалентные корни в эпидермисе. [ 12 ]
Бутоны и побеги
[ редактировать ]Авентаризные бутоны развиваются из мест, отличных от апикальной меристемы , которая встречается на кончике стебля или на узеле побега , у листового пазуха, почки оставляют там во время первичного роста. Они могут развиваться на корнях или листьях или на побегах в качестве нового роста. Стреляют апикальные меристемы производят одну или несколько подмышечных или боковых почек в каждом узле. Когда стебли производят значительный вторичный рост , подмышечные почки могут быть уничтожены. Авентативные бутоны могут затем развиваться на стеблях с вторичным ростом. [ Цитация необходима ]
Авентаризные бутоны часто образуются после того, как стебель ранен или обрезан . Авентаризные почки помогают заменить потерянные ветви. Адвентитные бутоны и побеги также могут развиваться на зрелых стволах деревьев, когда затененный ствол подвергается воздействию яркого солнечного света, потому что окружающие деревья вырубают. Деревья Redwood ( Sequoia sempervirens ) часто разрабатывают много экстренных бутонов на своих нижних стволах. Если главный ствол умирает, новый из них часто прорастает от одного из Adventious Buds. Маленькие кусочки красного туловища продаются в виде сувениров, называемых Redwood Burls. Они помещаются в кастрюлю с водой, а случайные бутоны прорастают, чтобы сформировать побеги. [ Цитация необходима ]
У некоторых растений обычно развиваются случайные бутоны на своих корнях, которые могут простираться на расстояние от растения. Съемки, которые развиваются из Adventious Buds на корнях, называются присоски . Они являются типом естественного вегетативного размножения во многих видах , например, многие травы, Quaring Aspen и The Canada Thistle . Пандо , Quicaling Aspen, вырос с одного туловища до 47 000 стволов через Adventitious Bud Formation в одной корневой системе. [ Цитация необходима ]
Некоторые листья развивают случайные бутоны, которые затем образуют случайные корни, как часть вегетативного размножения ; Например, растение Piggyback ( Tolmiea menziesii ) и мать тысяч ( Kalanchoe daigremontiana ). Затем случайные заводы снимают родительское растение и развиваются как отдельные клоны родителя. [ Цитация необходима ]
Коппинг - это практика разрезания стеблей деревьев на землю, чтобы способствовать быстрому росту случайных побегов. Традиционно используется для производства поляков, материала забора или дров. Это также практикуется для культур биомассы , выращиваемых для топлива, таких как тополь или ива.
Корни
[ редактировать ]
Авентаризация укоренения может быть акклимацией уклонения от стресса для некоторых видов, обусловленных такими входами, как гипоксия [ 13 ] или дефицит питательных веществ. Другой экологически важной функцией случайного укоренения является вегетативное воспроизведение видов деревьев, таких как Salix и Sequoia в прибрежных условиях. [ 14 ]
Способность стеблей растений формировать случайные корни используется в коммерческом распространении с помощью черенков . Понимание физиологических механизмов, лежащих в основе Adventitious Crooting, позволило добиться некоторого прогресса в улучшении укоренения черенков путем применения синтетических ауксинов в качестве порошков укоренения и использования селективного базального ранения. [ 15 ] Дальнейший прогресс может быть достигнут в последующие годы, применяя исследования в других регуляторных механизмах для коммерческого распространения и сравнительного анализа молекулярного и экофизиологического контроля случайного укоренения в «сложно -корневом» по сравнению с «простыми для корня» вида. [ Цитация необходима ]
Авентаризные корни и почки очень важны, когда люди распространяют растения через черенки, наслоение , культуру тканей . Растительные гормоны , называемые ауксинами , часто применяются к стеблям, стрельбам или листовым черенкам, чтобы способствовать образованию экстралия корня, например, африканских фиолетовых листьев и листьев Sedum и побегов Пуансеттии и Колея . Распространение с помощью корневых черенков требует образования для случайного бутона, например, в хренах и яблоке . В слои, на воздушных стеблях образуются случайные корни, прежде чем участок стебля будет удалена, чтобы сделать новое растение. Большие комнатные растения часто распространяются в результате слоя воздуха . Добросовестные корни и почки должны развиваться при распространении тканевой культуры растений.
Модифицированные формы
[ редактировать ]- КУБОРТАНИЕ КОНЕРСА НЕДОСТАВЛЯЕТ ОДНАКАЯ ФОРМА; Пример: сладкий картофель .
- Фирменной корень (бугристый корень) встречается в кластерах у основания стебля; Примеры: спаржа , Далия .
- Узеллевые корни распухли возле кончиков; Пример: куркума .
- Корни снятия возникают из первых нескольких узлов стебля. Они проникают косо вниз в почву и оказывают поддержку растению; Примеры: кукуруза , сахарный тростник .
- PROP -корни придают механическую поддержку воздушных ветвях. Боковые ветви растут вертикально вниз в почву и действуют как столбы; Пример: Баньян .
- Восходные корни, возникающие из -за узлов, прикрепляются к некоторой поддержке и поднимаются через него; Пример: epipremnum aureum .
- Moniliform или Beaded Crots Мятистые корни придают бисера, например: горькая тыква , Портулака .
Развитие листьев
[ редактировать ]Генетика развития формы листьев в Arabidopsis thaliana была разбита на три этапа: начало зачатка листьев , установление дорсивентральности и развитие маргинальной меристемы . Перчарчик листьев инициируется подавлением генов и белков семейства KNOX класса I (таких как апикальная меристема ). Эти белки класса I Knox напрямую подавляют биосинтез гиббеллина в примидие листьев. Было обнаружено, что многие генетические факторы участвуют в подавлении этих генов в перчатках листьев (таких как асимметричные листья1, лезвия-на-петиоле1 , пилот-1 и т. Д.). Таким образом, с этим подавлением уровни увеличения гиббеллина и роста листьев инициируют рост. [ Цитация необходима ]
Развитие цветов
[ редактировать ]
Развитие цветов - это процесс, с помощью которого покрытосеменные производят рисунок экспрессии генов в меристемах , который приводит к появлению органа, ориентированного на сексуальное размножение , цветок. Существует три физиологических события, которые должны происходить, чтобы это имело место: во -первых, растение должно переходить от сексуальной незрелости в сексуально зрелое состояние (то есть переход к цветению); Во -вторых, превращение функции апикальной меристемы из вегетативной меристемы в цветочную меристему или соцветие ; и, наконец, рост индивидуальных органов цветка. Последняя фаза была смоделирована с использованием модели ABC , которая описывает биологическую основу процесса с точки зрения молекулярной генетики и генетики развития . [ Цитация необходима ]

Внешний стимул требуется для того, чтобы вызвать дифференциацию меристемы в цветочную меристему. Этот стимул активирует деление митотических клеток в меристеме, особенно по бокам, где новые зачатки образуются . Этот же стимул также заставит меристему следовать схеме развития , которая приведет к росту цветочных меристем, а не к вегетативным меристемам. Основное различие между этими двумя типами меристемы, помимо очевидного несоответствия между объективным органом, заключается в филлотаксисе вертициллатного (или скрученного) , то есть отсутствием удлинения стебля среди последовательных оборотов или версий перчатка. Эти вертины следуют за акропетальной разработкой, вызывая чашелистики , лепестки , тычинки и ковры . Другое отличие от вегетативных подмышечных меристемов заключается в том, что цветочная меристема «определяется», что означает, что после дифференциации ее клетки больше не будут делиться . [ 16 ]
Идентичность органов, присутствующих в четырех цветочных вертиках, является следствием взаимодействия как минимум трех типов генных продуктов , каждый с различными функциями. Согласно модели ABC, необходимы функции A и C, чтобы определить идентичность вертиков перианта и репродуктивных вертиков, соответственно. Эти функции являются исключительными, и отсутствие одного из них означает, что другой будет определять идентичность всех цветочных вертиков. Функция B позволяет дифференцировать лепестки от чашелистиков во вторичном вертике, а также дифференцировку вычин от ковра на третичном вертике.
Цветочный аромат
[ редактировать ]Растения используют цветочную форму, цветок и аромат, чтобы привлечь различных насекомых для опыления . Определенные соединения в рамках испускаемого аромата обращаются к конкретным опылителям . В Petunia Hybrida образуются летучие бензолистые бензолиты , чтобы искать цветочный запах. В то время как компоненты бензольного биосинтетического пути известны, ферменты в пути и последующая регуляция этих ферментов еще предстоит обнаружены. [ 17 ]
Чтобы определить регуляцию пути, цветы P. hybrida Mitchell использовались в специфическом микрочипах с лепестком для сравнения цветов, которые были только что создали аромат, с цветами сорта P. hybrida W138, которые производят немного летучих бензолоидов. кДНК генов обоих растений были секвенированы. Результаты показали, что в цветах Митчелла активируется транскрипционный фактор, но не в цветах W138, в которых отсутствует цветочный аромат. Этот ген был назван Odorant1 (ODO1). Чтобы определить экспрессию ODO1 в течение дня, анализ РНК -гель -блоттинга был проведен . Гель показал, что уровни транскриптов ODO1 начали увеличиваться между 1300 и 1600 ч, достигали пика через 2200 ч и были самыми низкими через 1000 часов. Эти уровни транскриптов ODO1 непосредственно соответствуют временной шкале летучего бензеноидного излучения. Кроме того, гель подтвердил предыдущий вывод о том, что у не-слабых цветов W138 есть только одну десятую уровни транскрипта ODO1 цветов Митчелла. Таким образом, количество выполненного ODO1 соответствует количеству испускаемого бензеноида, что указывает на то, что ODO1 регулирует биосинтез бензеноидов. [ 17 ]
Дополнительными генами, способствующими биосинтезу основных ароматических соединений, являются OOMT1 и OOMT2. OOMT1 и OOMT2 помогают синтезировать орцинол о-метилтрансферазы (OOMT), которые катализируют последние два этапа пути DMT, создавая 3,5-диметокситолуол (DMT). DMT - это ароматическое соединение, производимое множеством разных роз, некоторые сорта роз, такие как Роза Галлика и Дамаска Роза Роза Дамаскен , не испускают DMT. Было высказано предположение, что эти сорта не делают ДМТ, потому что у них нет генов OOMT. Однако после эксперимента по иммунолокализации в лепестком эпидермисе был обнаружен OOMT. Чтобы изучить это, лепестки роз подвергались ультрацентрифугированию . Супернатанты и гранулы были проверены западным блотом . Обнаружение белка OOMT при 150 000 г в супернатанте, и осадка позволило исследователям сделать вывод, что белок OOMT тесно связан с мембранами эпидермиса лепестков. Такие эксперименты определили, что гены OOMT существуют в Роза Галлике и Дамаске Роза Rosa Damascene Разнообразии, но гены OOMT не экспрессируются в цветочных тканях, где производится DMT. [ 18 ]
Ссылки
[ редактировать ]- ^ Bäurle, я; Laux, T (2003). «Апикальные меристемы: фонтан молодости растения». Биологии . 25 (10): 961–70. doi : 10.1002/bies.10341 . PMID 14505363 . Обзор.
- ^ Дрост, Хайк-Георг; Джанитза, Филипп; Гросс, Иво; Quint, Marcel (2017). «Сравнение кросс-цалок с часами развития» . Текущее мнение в области генетики и развития . 45 : 69–75. doi : 10.1016/j.gde.2017.03.003 . PMID 28347942 .
- ^ Ири, Наоки; Куратани, Шигеру (2011-03-22). «Сравнительный анализ транскриптома выявляет филотипический период позвоночных во время органогенеза» . Природная связь . 2 : 248. Bibcode : 2011natco ... 2..248i . doi : 10.1038/ncomms1248 . ISSN 2041-1723 . PMC 3109953 . PMID 21427719 .
- ^ Domazet-Lousho, Tomislav; Tautz, Diethard (2010-12-09). «Филогенетически основанный на транскриптомом возрастной индекс отражает онтогенетическую дивергенцию». Природа . 468 (7325): 815–818. Bibcode : 2010natur.468..815d . doi : 10.1038/nature09632 . ISSN 0028-0836 . PMID 21150997 . S2CID 1417664 .
- ^ Квинт, Марсель; Дрост, Хайк-Георг; Габель, Александр; Улрих, Кристиан Карстен; Бенн, Маркус; Гросс, Иво (2012-10-04). «Транскриптомные песочные часы в эмбриогенезе растений». Природа . 490 (7418): 98-101. Bibcode : 2012 Natur.490 ... 98q . Doi : 10.1038/nature11394 . ISSN 0028-0836 . PMID 22951968 . S2CID 4404460 .
- ^ Дрост, Хайк-Георг; Габель, Александр; Гросс, Иво; Quint, Marcel (2015-05-01). «Свидетельство активного поддержания филотранскрипционных паттернов песочных часов у эмбриогенеза животных и растений» . Молекулярная биология и эволюция . 32 (5): 1221–1231. doi : 10.1093/molbev/msv012 . ISSN 0737-4038 . PMC 4408408 . PMID 25631928 .
- ^ Леопольд, А. Карл (1964). животное и там молодой . МакГроу-Хилл . п. 183.
- ^ Бренд, U; Хобе, м; Саймон, Р. (2001). «Функциональные домены в меристемах съемки растений». Биологии . 23 (2): 134–41. doi : 10.1002/1521-1878 (200102) 23: 2 <134 :: AID-bies1020> 3.0.co; 2-3 . PMID 11169586 . S2CID 5833219 . Обзор.
- ^ Барлоу, П (2005). «Определение узорчатых клеток в ткани растения: вторичная флоэма деревьев». Биологии . 27 (5): 533–41. doi : 10.1002/bies.20214 . PMID 15832381 .
- ^ Росс, SD; Pharis, RP; Биндер, WD 1983. Регуляторы и хвойные деревья роста: их физиология и потенциальное использование в лесном хозяйстве. п. 35–78 в Никелле, LG (ред.), Химические вещества, регулирующие рост растений. Тол. 2, CRC Press, Boca Raton FL.
- ^ Джонс, Синтия С. (1999-11-01). «Эссе о несовершеннолетних, изменении фазы и гетеробластике в семенных растениях». Международный журнал растительных наук . 160 (S6): 105 - S111. doi : 10.1086/314215 . ISSN 1058-5893 . PMID 10572025 . S2CID 21757481 .
- ^ McVeigh, I. 1938. Регенерация в Crassula Multicava . Американский журнал ботаники 25: 7-11. [1]
- ^ Дрю, MC; Джексон, MB; Гиффард С. (1979). «Профессиональный этилен, случайный укоренение и развитие корковых воздушных пространств (Aerenchyma) в корнях может быть адаптивным реакцией на наводнения в Зеа Мэйс L» . Планта 147 (1): 83–88. doi : 10.1007/bf00384595 . PMID 24310899 . S2CID 7232582 .
- ^ Найман, Роберт Дж.; Decamps, Anri (1997). «Экология интерфейсов: прибрежные зоны» . Ежегодный обзор экологии и систематики . 28 : 621–658. doi : 10.1146/annurev.ecolsys.28.1.621 . JSTOR 2952507 . S2CID 86570563 .
- ^ Церковь, грек-ян; Из Тира Крика, Wim; Молодой, шутка С. (1999). «Beview the Formation или Addentious Routes: новые концепции, новые позиции» . In vitro Cellular & Development Bology - растение . 35 (3): 189–199. doi : 10,1007/S11627-999-0076-z . S2CID 44027145 .
- ^ Azcón-bieto; и др. (2000). Основы физиологии растений . McGraw-Hill/Inter-American из Испании, Сау. ISBN 84-486-0258-7 . [ страница необходима ]
- ^ Подпрыгнуть до: а беременный Шуринк, Роберт С.; Харинг, Мишель А.; Кларк, Дэвид Г. (2006). «Регуляция летучего биосинтеза бензоидов в цветах петунии». Тенденции в науке о растениях . 11 (1): 20–25. doi : 10.1016/j.tlants.2005.09.009 .
- ^ Scalliet, Габриэль; и др. (2006-01-01). «Роль лепестков -специфических орцинолов ометилтрансферазы в эволюции аромата розы» . Физиология растений . 140 (1): 18–29. doi : 10.1104/pp.105.070961 . ISSN 1532-2548 . PMC 1326028 . PMID 16361520 .