Древесная ласточка
| Древесная ласточка | |
|---|---|
 | |
| Взрослый в заповеднике дикой природы Ямайка-Бэй в Квинсе, Нью-Йорк | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Птицы |
| Заказ: | Воробьинообразные |
| Семья: | Хирундиниды |
| Род: | Тахицинета |
| Разновидность: | Т. биколор |
| Биномиальное имя | |
| Тахицинета биколор ( Вьейе , 1808 г.) | |
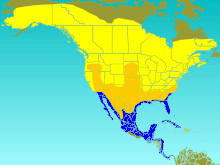 | |
| Ареал Т. биколор Размножающийся летний гость Посетитель миграции Зимний гость | |
| Синонимы | |
Хирундо биколор Вьейо, 1808 г. | |

Древесная ласточка ( Tachycineta bicolor ) — перелетная птица семейства Hirundinidae . Обнаруженная в Америке древесная ласточка была впервые описана в 1807 году французским орнитологом Луи Вьейо как Hirundo bicolor . С тех пор он был перенесен в нынешний род Tachycineta , внутри которого обсуждается его филогенетическое положение. У древесной ласточки верхняя часть тела блестящая сине-зеленая , за исключением черноватых крыльев и хвоста, а нижняя часть тела белая . Клюв черный, глаза темно-коричневые, ноги и ступни бледно-коричневые. Самка обычно более тусклая, чем самец, а у самки-первокурсника верхняя часть тела в основном коричневая, с некоторыми синими перьями. У молодых особей коричневый верх и серо-коричневая грудь. Древесная ласточка гнездится в США и Канаде. Зимует вдоль южных побережий США на юг, вдоль побережья Мексиканского залива, до Панамы и северо-западного побережья Южной Америки, а также в Вест-Индии .
Гнездится древесная ласточка либо изолированными парами, либо рыхлыми группами, как в естественных, так и в искусственных полостях. Размножение может начаться уже в начале мая, хотя из-за изменения климата эта дата наступает раньше , и может закончиться уже в июле. Эта птица в целом социально моногамна (хотя около 8% самцов полигинны ), с высоким уровнем внепарного отцовства . Это может принести пользу самцу, но, поскольку самка контролирует совокупление, отсутствие четкого представления о том, какую пользу такое поведение приносит самкам, делает высокий уровень отцовства внепарной пары загадочным. Самка насиживает кладку из двух-восьми (но обычно от четырех до семи) чисто-белых яиц в течение примерно 14-15 дней. Птенцы вылупляются немного асинхронно, что позволяет самке расставлять приоритеты, каких птенцов кормить во время нехватки еды. Обычно они оперяются примерно через 18–22 дня после вылупления. Древесную ласточку иногда считают модельным организмом из-за большого количества исследований, проведенных с ней.
Воздушное насекомоядное дерево глотает корм как в одиночку, так и группами, питаясь в основном насекомыми , а также моллюсками , пауками и фруктами. Птенцы, как и взрослая особь, питаются преимущественно насекомыми, которых им скармливают оба пола. Эта ласточка уязвима для паразитов, но, находясь на птенцах, они не причиняют особого вреда. Эффект болезни может усиливаться по мере взросления древесной ласточки, поскольку с возрастом некоторые части иммунной системы ослабевают. приобретенный Т-клеточный Например, иммунитет снижается с возрастом, тогда как как врожденный, так и приобретенный гуморальный иммунитет этого не делают. считает, что из-за большого ареала и стабильной популяции древесная ласточка вызывает наименьшее беспокойство Международный союз охраны природы . В США он защищен Законом о перелетных птицах 1918 года , а в Канаде – Законом о Конвенции о перелетных птицах . На эту ласточку негативно влияет деятельность человека, например, вырубка лесов; закисленные озера могут заставить гнездящуюся древесную ласточку преодолевать большие расстояния в поисках богатых кальцием продуктов питания, которыми они могли бы накормить своих птенцов.
Таксономия и этимология
[ редактировать ]Древесная ласточка была описана как Hirundo bicolor Луи Пьером Вьейо в его «Естественной истории птиц Северной Америки» , опубликованной в 1807 году. [2] Затем его отнесли к нынешнему роду Tachycineta , когда Жан Кабанис основал его в 1850 году. [3] В 1878 году Эллиот Куэс предложил по признаку оперения отнести древесную ласточку, по крайней мере, к отдельному подроду Iridoprocne , наряду с белокрылой ласточкой , чилийской ласточкой , белохвостовой ласточкой и мангровой ласточкой . глотать . [4] К 1882 году он повысил статус этого рода до полноценного. [5] Некоторые авторы продолжали использовать эту классификацию, добавив ласточку Тумбеса ; однако генетические данные подтверждают существование единственного рода Tachycineta . [6] Древесную ласточку еще называют белобрюхой из-за ее белой нижней части тела. [7]
Общепринятое название рода происходит от древнегреческого takhykinetos , «быстро движущийся», а видовое bicolor — латинское и означает «двухцветный». [8] Другое название рода, Iridoprocne , происходит от греческого iris , что означает радуга, и Procne , фигуры, которая предположительно превратилась в ласточку. [5]
Как именно древесная ласточка связана с другими представителями Tachycineta , неясно. В исследованиях, основанных на митохондриальной ДНК , он был помещен в базальное (то есть первое ответвление в древе видов) североамериканско-карибскую кладу, состоящую из фиолетово-зеленой ласточки , золотой ласточки и багамской ласточки . [9] [10] Хотя митохондриальная ДНК считается лучшим индикатором эволюционных изменений, поскольку она быстро развивается, анализ, основанный на ней, может пострадать, поскольку она наследуется только от матери, что делает ее хуже, чем ядерная ДНК из нескольких локусов , при представлении филогении целой группы. [11] Исследование, основанное на такой ядерной ДНК, поместило древесную ласточку в самое базальное положение среди Tachycineta в целом (как сестринскую группу по отношению к остальной части рода). [12]
Описание
[ редактировать ]
Древесная ласточка имеет длину от 12 до 14 см (от 4,7 до 5,5 дюйма) и вес примерно от 17 до 25,5 г (от 0,60 до 0,90 унции). Размах крыльев колеблется от 30 до 35 см (от 12 до 14 дюймов). [13] У самца верхняя часть тела преимущественно блестящая, сине-зеленая , крылья и хвост черноватые. Нижняя часть тела [14] и пятно на щеке белое, [7] хотя подкрыльевые кроющие серо-коричневые. [14] Клюв черный, глаза темно-коричневые, ноги и ступни бледно-коричневые. [7] Самка более тусклая, чем самец, и иногда имеет коричневый лоб. [14] Второй год [примечание 1] у самки также коричневая верхняя часть тела с переменным количеством синих перьев; некоторые самки третьего года жизни также сохраняют часть этого полувзрослого оперения. [15] Согласно исследованию 1987 года, это, вероятно, позволяет молодой самке исследовать места гнездования, поскольку местный самец обычно менее агрессивен по отношению к несовершеннолетней самке. [16] Исследование 2013 года показало, что постоянные женщины были менее агрессивны по отношению к моделям-второкурсницам, когда они были представлены отдельно от моделей старшего возраста. Почему самка в конечном итоге меняет свое полувзрослое оперение, неизвестно; это может позволить самцам оценить качество самок, поскольку пары спариваются ассортативно в зависимости от яркости оперения. [17] Молодую древесную ласточку можно отличить по коричневому верху и серо-коричневой груди. [14]
Голос
[ редактировать ]

Песня ласточки состоит из трех частей: стрекотания, визга и бульканья. Эти разделы могут повторяться или опускаться, и все они могут стоять отдельно. Первый, чирикающий призыв (иногда разделенный на контактный призыв и призыв-призыв), издается самкой во время совокупления. [18] и у обоих полов, чтобы стимулировать птенцов к попрошайничеству. [19] или (в некоторых популяциях), когда их партнерша покидает полость гнезда или входит в нее. Вой, обычно состоящий из понижения частоты, за которым следует повышение, может быть подан отдельно как тревожный сигнал. [18] иногда производится в ответ на действия определенных хищников. [20] Бульканье, например, когда оно появляется в конце песни, обычно произносится дважды. Вероятно, он участвует в парном соединении. Разговорный звонок используется для рекламы мест гнездования (по этой причине он также известен как «рекламный звонок места гнездования»), а также передаётся вторгающимся сородичам. Короткий пронзительный призыв к подчинению иногда раздается после агрессивной встречи с другой древесной ласточкой. Если человек физически ограничен или испытывает боль, может быть подан сигнал бедствия. Самец часто издает тикающий (или грубый) агрессивный крик во время совокупления, и представители обоих полов используют его в конце ныряний. Сигнал тревоги подается в ответ на хищников и других злоумышленников. [18] и может побудить старших птенцов присесть и перестать просить милостыню, когда хищник находится рядом. [21]
Общение между родителями и потомством может быть нарушено из-за шума, создаваемого человеком. Исследование 2014 года, например, показало, что выводки, для которых воспроизводился белый шум, с меньшей вероятностью приседали или прекращали просить милостыню в ответ на сигналы тревоги. Родители не изменили свои призывы в качестве компенсации, что, вероятно, увеличило риск нападения хищников. [22] Шум также может повлиять на реакцию родителей на попрошайничество, но это может быть уравновешено более громкими криками, которые издают птенцы, когда они подвергаются его воздействию. Однако увеличение усилий попрошайничества может оказаться неэффективным или дорогостоящим из-за более громкого шума. [23]
Распространение и среда обитания
[ редактировать ]Древесная ласточка гнездится в Северной Америке. Его ареал простирается до северо-центральной Аляски и до границы деревьев в Канаде. Он встречается на юге до Теннесси в восточной части своего ареала, Калифорнии и Нью-Мексико на западе и Канзаса в центре. Иногда он гнездится южнее США. [7] а бродяги иногда встречаются за Полярным кругом , в северной части Тихого океана , в Гренландии и Европе. [14] Ареал зимовки — от Калифорнии и юго-западной Аризоны на западе и юго-восточной Вирджинии на востоке-юге вдоль побережья Мексиканского залива до Вест-Индии , Панамы и северо-западного побережья Южной Америки. [7] Во время миграции эта ласточка часто останавливается, проводя в этих местах осенью в среднем 57 дней. Чтобы добраться до места зимовки, он часто использует один из трех пролетных путей: западный пролетный путь, к западу от Скалистых гор ; Центральный пролетный путь между Скалистыми горами и Великими озерами , простирающийся на юг до Восточной Мексики; и восточный пролетный путь, ведущий от Великих озер на восток. [24] Когда ласточка возвращается в гнездо, она обычно не меняет места гнездования. [25]
Средой размножения этой птицы являются преимущественно открытые и лесистые местности, особенно вблизи воды. [7] Он ночует каждую ночь в период отсутствия размножения, предпочитая отдыхать в тростника или зарослях тростника , а не в воде, но его также можно встретить на суше, на деревьях и проводах. Места для ночлега обычно находятся на расстоянии от 100 до 150 километров (от 62 до 93 миль) друг от друга. [26]
Поведение
[ редактировать ]
Из-за большого количества исследований древесной ласточки и того, как она охотно размножается в гнездовых ящиках, биолог Джейсон Джонс рекомендовал считать ее модельным организмом . [27] Хотя в период размножения она агрессивна, за ее пределами эта ласточка общительна, образуя стаи, иногда насчитывающие тысячи птиц. [7]
Разведение
[ редактировать ]
Гнезда древесных ласточек строятся в сооружениях с уже существовавшими норами, как естественными, так и искусственными. Когда-то их можно было встретить только в лесных районах, но строительство гнездовых ящиков позволило этой птице распространиться на открытые места обитания. [7] Эта ласточка обычно гнездится на месте, где она выплодилась годом ранее; только около 14% самок и 4% самцов ежегодно расходятся для размножения на новом участке. Большинство из них не уходят далеко, обычно гнездясь на участках менее чем в 10 километрах (6,2 мили) от своего первоначального места обитания. На расселение влияет успех размножения; из взрослых самок, которым не удается оперить птенца, около 28% рассеиваются по сравнению с 5% успешных производителей. [25] Натальное расселение (когда птица не возвращается на место, где она родилась для размножения) является обычным явлением для древесной ласточки и происходит чаще, чем гнездовое расселение. [28] Гнездится как рыхлыми группами, так и изолированными парами. При гнездовании рыхлыми группами гнезда обычно располагаются на расстоянии не менее 10–15 метров (от 33 до 49 футов) друг от друга. [14] а те, что ближе по расстоянию, обычно находятся дальше друг от друга с точки зрения даты кладки. [29] В естественных полостях древесная ласточка гнездится на расстоянии примерно 27 метров (89 футов) от своей соседки. Гнездо в таких ситуациях находится в среднем на высоте 3,4 метра (11 футов) над уровнем земли, хотя около 45% из них находятся на высоте менее 2 метров (6,6 футов) над землей. Вероятно, предпочтение отдается более высоким полостям, поскольку они уменьшают хищничество, тогда как более низкие отверстия для гнезд можно выбрать, чтобы избежать конкуренции. Ширина входа часто составляет от 4 до 5 сантиметров (от 1,6 до 2,0 дюйма), тогда как высота входа более варьируется: исследование 1989 года выявило проемы в диапазоне от 3,5 до 26 сантиметров (от 1,4 до 10,2 дюйма). Объем полости обычно ниже 1000 см3. 3 (61 куб. Дюймов). [30] Найдя подходящее место для гнезда, самец садится возле него и часто кричит. Отсутствие мест может вызвать драки между птицами, иногда приводящие к гибели. Эта ласточка обычно защищает территорию вокруг гнезда радиусом около 4,6 метра (15 футов). [7] а также дополнительные гнезда на этой территории , [14] блокируя вход в гнездо и преследуя злоумышленников. [7] Сама чашка гнезда сделана из травы, мха, хвои и водных растений, собираемых в основном самкой. [14] и выстлан перьями, собранными в основном самцами в боях. [31] Перья могут изолировать гнездо, сокращая время инкубации. [32] и, вероятно, предотвращение переохлаждения у цыплят. Помимо более быстрого роста цыплят, [31] яйца в гнездах с перьями остывают медленнее, чем в гнездах без перьев. [33] Однако исследование, опубликованное в 2018 году, не обнаружило значимой корреляции между количеством перьев в гнездах, которые были искусственно утеплены, и количеством перьев, которые не были утеплены. Кроме того, было обнаружено, что в гнездах в Сен-Дени, Саскачеван, использовалось значительно меньше перьев, чем в гнездах в долине Аннаполис , несмотря на то, что первые находились севернее. Однако температура в Новой Шотландии (где находится долина Аннаполис) обычно ниже, чем в Саскачеване, что, возможно, объясняет неожиданный результат. [34]

Во время ухаживания самец древесной ласточки нападает на неизвестную самку. Это можно стимулировать полетом самки с трепещущими крыльями, что может быть приглашением ко двору. Затем самец может принять вертикальную позу с поднятым и слегка расправленным хвостом, а крылья взмахивают и слегка опускаются. Это побуждает самку попытаться приземлиться на спину самцу, но он летит, чтобы предотвратить это; это повторяется. После ухаживания за самкой самец летит к выбранному им месту гнезда, которое самка осматривает. Во время совокупления самец нависает над самкой, а затем садится на нее, издавая тикающие крики. Затем он вступает в клоакальный контакт с самкой, держа перья ее шеи в своем клюве и стоя на ее слегка расправленных крыльях. Копуляция происходит несколько раз. [7]
Яйца откладываются с начала мая до середины июня (правда, из-за изменения климата это происходит раньше). [35] ), а птенцы оперяются в период с середины июня по июль. [14] Широта положительно коррелирует с датой яйцекладки. [14] в то время как возраст самки и длина крыльев (более длинные крылья позволяют более эффективно добывать пищу [36] ) отрицательно коррелируют. [37] Древесная ласточка, вероятно, является прибыльным производителем , поскольку ее размножение зависит от обилия пищи и температуры в период яйцекладки. [36] Этот вид в целом социально моногамен , но до 8% размножающихся самцов полигинны . [14] На полигинию влияет территория: самцы, имеющие территории с гнездовыми ящиками на расстоянии не менее 5 метров (16 футов) друг от друга, с большей вероятностью будут полигинными. [38] Предполагается, что эта полигиния зависит от условий в период яйцекладки: лучшие условия, такие как обилие пищи, позволяют самкам в полигинии, которые не получают помощи в поисках пищи, откладывать больше яиц. [39]

У древесной ласточки высокий уровень внепарного отцовства: от 38% до 69% птенцов являются продуктом внепарного отцовства и от 50% до 87% выводков, содержащих хотя бы одного птенца, появившегося в результате внепарного отцовства. совокупление. [14] Одним из факторов, который может способствовать этому, является то, что самки контролируют совокупление, что делает защиту отцовства неэффективной. [40] Это можно смягчить за счет более частых совокуплений непосредственно перед откладкой яиц, согласно исследованию 2009 года, которое показало, что попытки внутрипарного совокупления достигают пика за три-один день до откладки первого яйца и что более успешные попытки в этот период увеличивают долю внутрипарных совокуплений. -пара молодых самцов. Этот последний вывод противоречит выводам исследований 1993 и 1994 годов. [41] Отцовство внепарной пары не меняет уровень родительской заботы, которую самец вносит в ласточку. [42] Значительное количество отцов дополнительных пар могут быть плавающими (присутствующими в местах размножения, которые предположительно не размножаются). Исследование 2001 года показало, что из 35 дополнительных пар птенцов 25 были получены от местных жителей, трое - от жителей близлежащих участков и семь - от плавающих самцов. Таким образом, у древесной ласточки плавание помогает самцам в хорошем состоянии производить больше птенцов, а самцам в плохом состоянии добиться успеха, вкладывая средства в родительскую заботу. [43] Существует также значительная популяция женщин с поплавками; Исследование 1985 года показало, что от 23% до 27% женщин имели плавающие помутнения, из которых от 47% до 79% были несовершеннолетними. [44]
Почему самки участвуют в совокуплении дополнительных пар и как они выбирают себе партнеров, остается спорным. Одна теория, называемая гипотезой генетической совместимости, утверждает, что повышенная приспособленность потомства является результатом повышенной гетерозиготности , и, таким образом, самки древесных ласточек предпочитают спариваться с самцами, которые менее генетически похожи на них. Самки также могут выбирать сперму после совокупления, чтобы обеспечить совместимого партнера. В подтверждение этой теории исследование 2007 года показало, что потомство вне пары было более гетерозиготным, чем потомство внутри пары. [45] Однако в статье 2005 года была обнаружена небольшая отрицательная корреляция между генетическим сходством пары и долей детенышей дополнительной пары в их гнезде. [46] Теория хороших генов гласит, что самки выбирают самцов из дополнительной пары на основе качества их генов. Это могло бы объяснить, почему у некоторых древесных ласточек нет лишней пары детенышей, а у других есть. [40] Однако большинство исследований не обнаружили фенотипических различий между самцами вне пары и внутри пары (хотя исследование 2007 года действительно показало, что более старые самцы с более ярким оперением с большей вероятностью спаривались вне парной связи). [47] Кроме того, согласно тезису 2017 года, потомство вне пары с большей вероятностью присоединится к своей натальной популяции, чем потомство внутри пары. Другая теория предполагает, что отцовство внепарной пары зависит от контекста: детеныши внепарной пары превосходят детенышей внутри пары в определенных ситуациях и уступают в других средах. В диссертации 2017 года, например, было обнаружено, что молодь внепарной пары была крупнее, тяжелее и более длиннокрылой, чем молодь внутри пары, когда оба подвергались воздействию хищников, в то время как молодь внутри пары была тяжелее, чем молодь внепары, когда они были показаны средства передвижения, не относящиеся к хищникам. В этом тезисе также было обнаружено, что детеныши внутри пары превосходили детенышей вне пары с точки зрения приспособленности на протяжении всей жизни, когда они росли в менее изменчивой среде, что позволяет предположить, что потомство вне пары имеет меньшую пластичность развития, чем потомство внутри пары. [48] Исследование 2018 года слабо подтвердило эту контекстно-зависимую гипотезу, обнаружив, что потомство дополнительной пары с большей вероятностью оперяется, чем потомство внутри пары в экспериментально увеличенных выводках; однако ни длина теломер (коррелят выживаемости и репродуктивного успеха), ни размер через 12 дней после вылупления существенно не различались среди этих детенышей, и не было обнаружено существенных различий между двумя типами в неувеличенных выводках. [49]
Исследования, пытающиеся доказать адаптивность отцовства внепарной пары для самок, подвергались критике за отсутствие положительного эффекта, который могло бы иметь повышение приспособленности потомства по сравнению с потенциальными издержками снижения приспособленности самки. [50] например, усиление хищничества в результате поиска партнеров. [51] Таким образом, постулируются теории, основанные на неадаптивности внепарного отцовства самок. Эти теории основаны на генетическом ограничении , когда аллель, приводящая к неадаптивному поведению, сохраняется, поскольку она также способствует благоприятному фенотипу. Теория интерсексуальной антагонистической плейотропии утверждает, что сильный отбор по отцовству внепарной пары у самцов (как видно на примере этой птицы) [50] ) отменяет слабый отбор против отцовства внепары у самок. Между тем, гипотеза внутриполовой антагонистической плейотропии утверждает, что отцовство вне пары присутствует, потому что гены, регулирующие его, оказывают плейотропное воздействие на аспекты женской приспособленности. [52] как скорость совокупления внутри пары. [50]

Древесная ласточка кладет кладку от двух до восьми человек, хотя обычно от четырех до семи. [14] чисто-белые и полупрозрачные при кладке яйца размером примерно 19 на 14 мм (0,75 на 0,55 дюйма). [53] Эти яйца насиживает самка. [14] обычно после откладки предпоследнего яйца, [54] от 11 до 20 дней, [53] хотя большинство из них вылупляются через 14–15 дней. Около 88% гнезд дают хотя бы одного птенца, но это может быть снижено из-за плохой погоды и более молодой гнездящейся самки. [14] Яйца обычно вылупляются в том порядке, в котором они были отложены. Они также вылупляются немного асинхронно: в среднем между появлением первого и последнего птенца проходит 28 часов. Это может привести к весовой иерархии, при которой цыплята, вылупившиеся раньше, весят больше (особенно в начале периода вылупления), чем цыплята, вылупившиеся позже, что позволяет самке определять приоритетность того, кому цыпленок давать еду во время нехватки еды. Вероятно, наибольший эффект это оказывает в начале периода вылупления птенцов, поскольку к 12 дням после вылупления существенных различий в весе обычно нет. [54] Детоубийство птенцов и яиц иногда происходит, когда самца заменяют другим самцом. Детоубийство обычно не происходит, когда кладка не завершена, поскольку тогда у сменных самцов есть возможность оплодотворить хотя бы одно яйцо. Когда самец прибывает во время инкубации, он иногда совершает детоубийство, но в других случаях усыновляет яйца, поскольку есть вероятность, что некоторые яйца были получены от нового самца. Если новый самец прибывает после вылупления птенцов, обычно совершается детоубийство, хотя иногда самка предотвращает это. [55]
В гнездах, созданных самками в лучшем состоянии, соотношение полов часто смещено в сторону высококачественных самцов. Исследование 2000 года предположило, что это связано с тем, что репродуктивный успех самцов варьируется, и, следовательно, самец высокого качества производит больше потомства, чем самка аналогичного качества. [56]
На рост и выживание птенцов древесных ласточек влияет окружающая среда. Как у молодых, так и у старших птенцов (в возрасте от двух до четырех дней и от девяти до одиннадцати дней соответственно) на рост положительно влияет более высокая максимальная температура, особенно у первых. Более поздний срок вылупления отрицательно влияет на рост, особенно на молодых птенцов. Птенцы постарше растут несколько быстрее, когда насекомых много. Рост молодых птенцов увеличивается с возрастом, а у старых птенцов он уменьшается по мере взросления. [57] Молодые древесные ласточки способны осуществлять терморегуляцию по крайней мере на 75% так же эффективно, как и взрослые особи, в среднем в возрасте 9,5 дней вне гнезда и от четырех до восьми дней в гнезде (в зависимости от размера выводка). [58] Птенцы оперяются примерно через 18–22 дня, при этом успех оперения составляет около 80%. Как и на успех высиживания, на это негативно влияют неблагоприятная погода и более молодая самка. [14] На птенцов могут охотиться змеи [59] и еноты. [60] Это хищничество может усугубляться звонками с попрошайничеством. [61]
Кормление
[ редактировать ]
Древесная ласточка добывает корм на высоте до 50 метров (160 футов) над землей поодиночке или группами. Его полет представляет собой смесь взмахов и скольжения. В период размножения это место обычно находится в пределах 4 километров (2,5 миль) от места гнездования. Однако в поисках птенцов он обычно уходит на расстояние до 200 метров (660 футов) от гнезда. [14] в основном остается на виду и добывает корм на высоте до 12 метров (39 футов). [62] Помимо того, что его поймали в полете, [7] насекомых иногда добывают с земли, воды, растительности и вертикальных поверхностей. [14]
Древесная ласточка питается в основном насекомыми, иногда моллюсками, пауками и фруктами. В Северной Америке мухи составляют около 40% рациона, дополненного жуками и муравьями. В остальном рацион примерно на 90% состоит из мух. [14] Выловленные насекомые представляют собой смесь водных и наземных организмов; первые являются важным источником высоконенасыщенных жирных кислот омега-3 . Это связано с тем, что, хотя древесная ласточка и может превращать предшественник α-линоленовой кислоты в высоконенасыщенные жирные кислоты, такие как докозагексаеновая кислота , она не может делать это в необходимых количествах. [63] Семена и ягоды в основном принадлежат к роду Myrica , который в основном потребляется во все четыре сезона Северного полушария, кроме лета. Исследование, проведенное на Лонг-Айленде, штат Нью-Йорк , также показало, что ракообразные играют важную роль в зимнем рационе. [14]

Оба пола кормят птенцов (хотя самец кормит птенцов меньше, чем самки), в результате чего получается от 10 до 20 кормлений в час. [14] Родители часто используют щебетание, чтобы побудить птенцов просить милостыню. Это чаще используется с молодыми цыплятами, поскольку они просят меньше, чем старшие цыплята, когда родитель приходит с едой, но не зовет. [19] Вероятность попрошайничества в отсутствие родителей также увеличивается с возрастом. [64] Порядок вылупления влияет на то, сколько цыпленка кормят; птенцов, вылупившихся последними (в случаях, когда вылупление асинхронное), скорее всего, кормят меньше, чем вылупившихся ранее. [65] Птенцы, находящиеся ближе к входу в гнездо, также с большей вероятностью будут накормлены, как и те, кто попрошайничает первым. [66] и чаще. Общая скорость кормления выводка также увеличивается с увеличением количества попрошайничества. [67] Сам рацион состоит в основном из насекомых, причем отряды Diptera , Hemiptera и Odonata . большую часть рациона составляют [14] Эти насекомые обычно имеют размер до 10 мм (0,39 дюйма), но иногда достигают 60 мм (2,4 дюйма) в длину. [62] В гнездах вблизи закисленных человеком озер кальциевые добавки, в первую очередь рыбьи кости, экзоскелеты раков , панцири моллюсков и скорлупу птичьих яиц, труднее найти. Это вынуждает взрослую древесную ласточку путешествовать дальше, чем обычно — иногда на расстояние до 650 м (2130 футов) от гнезда — чтобы получить эти добавки кальция. [68]
Выживание
[ редактировать ]Средняя продолжительность жизни древесной ласточки составляет 2,7 года. [69] и максимум 12 лет. Около 79% людей не доживают до первого года жизни, а те, кто выживает, сталкиваются с ежегодной смертностью от 40% до 60%. [14] Большинство смертей, вероятно, является результатом холодной погоды, которая снижает доступность насекомых, что приводит к голоду. [7] Продолжительность жизни связана с длиной теломер: исследование 2005 года, в котором использовалась скорость возвращения (к месту размножения в предыдущем году) в качестве показателя выживаемости, показало, что люди с самыми длинными теломерами в возрасте одного года имели прогнозируемую продолжительность жизни 3,5 года по сравнению с до 1,2 лет для людей с самыми короткими теломерами. [70] Неизвестно, вызывают ли короткие теломеры снижение физической формы или являются просто индикатором этого. Тем не менее, диссертация 2016 года показала, что показатели физического состояния положительно коррелируют с длиной теломер. У самцов также обычно были более длинные теломеры, чем у самок, как и у птиц с меньшими крыльями. Особи с более короткими теломерами могут компенсировать потенциальные потери приспособленности за счет увеличения репродуктивных усилий, тогда как особи с более длинными теломерами могут уменьшить свои вложения, о чем свидетельствует меньшая доля оперившихся самок птенцов с более длинными теломерами. [71] Длина теломер сильно наследуется и особенно зависит от длины теломер. [72]
Хищничество
[ редактировать ]Древесная ласточка уязвима для широкого круга хищников. Яйца, птенцы и взрослые особи в гнезде становятся жертвами черных крысиных змей , американских ворон , американских пустельг , обыкновенных граклей северных мерцаний , бурундуков , мышей-оленей , домашних кошек , ласк , [73] Американские черные медведи , [74] и еноты . [75] Во время полета или сидя к древесным ласточкам относятся следующие хищники: американская пустельга, черноклювая сорока , [73] полосатые совы , [76] большие рогатые совы , кречеты , сапсаны и остроногие ястребы . Хотя у свободно летающих ласточек бегство является обычной реакцией на хищников, вокруг гнезда распространено поведение толпы. [73] и направлен не только на хищников, но и на конкурентов места гнездования, которых он может отпугнуть. [20] Такое поведение предполагает, что ласточка роится и ныряет к злоумышленнику (но не наносит ему удар). [73] на высоте примерно от 5 до 20 м (от 16 до 66 футов) над землей, обычно издавая тихие тикающие крики ближе к концу и приближаясь к хищнику на расстояние от 0,5 до 2 м (от 1,6 до 6,6 футов). [20] Кажется, он меняет интенсивность своих атак в зависимости от того, какой хищник приближается; [73] Исследование 1992 года показало, что хорьки вызывают более энергичную защиту, чем змеи-черные крысы. [20] а диссертация 2019 года аналогичным образом обнаружила, что модели змей черной крысы ныряли меньше всего, а модели восточного бурундука - больше всего. [77] Предполагается, что змея вызвала более слабую реакцию, поскольку защитное поведение может быть менее эффективным. [20] и опаснее выступать против него. [77]
Паразиты
[ редактировать ]
Древесная ласточка уязвима для различных паразитов, таких как кровяной паразит Trypanosoma . Он также восприимчив к блохе Ceratophyllus idius и перьевым клещам Pteronyssoides tyrrelli , Trouessartia и (вероятно) Hemialges . Вероятно, он также поражается вшами родов Brueelia и Myrsidea . Существует корреляция между количеством блох на птице и количеством птенцов, о которых она заботится. Предполагается, что эта связь возникает из-за улучшения микроклимата для блох из-за более крупной кладки. [78] Птенцы также страдают от паразитов, например, мух рода Protocalliphora . [79] что приводит к потере крови птенцами. [80] Однако эти паразиты встречаются в большинстве гнезд и, похоже, не оказывают большого влияния на птенцов. Исследование, опубликованное в 1992 году, показало, что последствия паразитизма мух-мух объясняют лишь около 5,5% различий в массе птенцов. [79]
Иммунология
[ редактировать ]У размножающихся самок древесной ласточки гуморальная иммунокомпетентность (HIC) обратно коррелирует с датой кладки. Это означает, что в среднем птица, откладывающая яйца раньше, имеет более сильную реакцию антибиотика на антиген, чем птица, откладывающая яйца позже. Древесная ласточка, страдающая от подрезания крыльев, обычно имеет более низкий HIC. Эти взаимосвязи можно было бы интерпретировать как подтверждение вывода о том, что самка, которая откладывает яйца раньше, приобретает более высокий HIC, но авторы исследования, обнаружившего корреляцию, считали это маловероятным из-за более низких температур в начале сезона размножения. Вместо этого они подумали, что HIC может быть мерой качества и что самка более высокого качества способна отложить потомство раньше. Авторы также предположили, что это показатель рабочей нагрузки, о чем свидетельствует более низкий HIC у птиц-инвалидов. [81]
Самки древесных ласточек более высокого качества (по дате кладки) способны поддерживать свои репродуктивные усилия, отвлекая ресурсы на борьбу с иммунными проблемами. Ласточки более низкого качества менее способны на это; Исследование 2005 года в Итаке, штат Нью-Йорк, показало, что самки, откладывающие поздно, с искусственно увеличенным выводком, хотя и были способны поддерживать качество потомства, имели более низкую реакцию на иммунный вызов, чем те, которые были более высокого качества или не имели увеличенного выводка. [82] Выбор самки в качестве приоритета качества потомства или иммунокомпетентности, вероятно, связан с вероятностью выживания; Исследование 2005 года показало, что самки с увеличенным выводком на Аляске, где показатели выживаемости ниже, имели более слабые иммунные реакции, но сохраняли стабильные репродуктивные усилия, тогда как самки в Теннесси, с более высокими показателями выживаемости, имели более сильную реакцию, но потомство более низкого качества. [83]
У древесной ласточки с возрастом ухудшаются некоторые компоненты иммунной системы. приобретенный Т-клеточный Например, иммунитет снижается с возрастом у самок древесной ласточки. Но возраст самки не влияет ни на приобретенный, ни на врожденный гуморальный иммунитет ; отсутствие ухудшения показателей первых контрастирует с исследованиями ласточек и самок мухоловок . [84] Из-за этого иммуностарения (снижения иммунной функции с возрастом) пожилые самки, инфицированные этим заболеванием, обычно меньше посещают свое гнездо, в результате чего их птенцы растут медленнее. Они также могут потерять вес из-за инфекции. [85]
Статус
[ редактировать ]считает, что древесная ласточка вызывает наименьшее беспокойство Международный союз охраны природы . Это связано с большим радиусом действия птицы - около 834 000 км. 2 (322 000 квадратных миль), [1] и его стабильная популяция, по оценкам, составляет около 20 000 000 особей. [14] В США он охраняется Законом о перелетных птицах 1918 года . [86] а в Канаде – Законом о Конвенции о перелетных птицах . [87] В некоторых частях США ареал этой ласточки расширился на юг, вероятно, из-за изменений в землепользовании, реинтродукции бобров и установки гнездовых ящиков для синих птиц . На древесную ласточку негативно влияют вырубка лесов и сокращение болот, что приводит к сокращению среды обитания, доступной для зимовки. Этой ласточке приходится конкурировать за места гнездования с обыкновенным скворцом , домашним воробьем (оба завезены в Северную Америку ), [14] синие птицы, [88] и домовой крапивник (который также разрушает гнезда, не занимая их). [89] Закисление озер может заставить эту ласточку преодолевать относительно большие расстояния в поисках предметов, богатых кальцием, а птенцы могут поедать пластик. [68] Другие химические вещества, такие как пестициды и другие загрязнители, могут иметь высокую концентрацию в яйцах, а ПХБ связаны с отказом от кладки пары. [14] Загрязнение с участков добычи нефтеносных песков может отрицательно повлиять на древесных ласточек, увеличивая количество токсинов, что измеряется по активности этоксирезоруфино - деэтилазы (фермента детоксикации) у птенцов. Обычно это мало влияет на птенцов и оперение. [90] хотя экстремальные погодные условия могут выявить последствия: исследование 2006 года показало, что птенцы из водно-болотных угодий, наиболее загрязненных материалами переработки нефтеносных песков, имели более чем в 10 раз больше шансов умереть, чем птенцы из контрольного участка в периоды синхронизированных низких температур и сильных дождей, по сравнению с отсутствие разницы в смертности между группами, когда погода была менее экстремальной. [91] Однако в статье 2019 года было обнаружено, что увеличение количества осадков привело к аналогичному снижению успешности вылупления и вылупления птенцов как вблизи, так и вдали от участков нефтеносных песков. [92] В другом исследовании птицы, подвергшиеся воздействию ртути, оперились в среднем на одного птенца меньше, чем те, кто не подвергался воздействию ртути, и этот эффект усиливался в теплую погоду. [93] Кроме того, холодные погодные явления могут быстро сократить доступность воздушных насекомых-жертв. [94] а в некоторых популяциях с ускоренным воспроизводством это может привести к снижению выживаемости потомства. [95]
Ссылки
[ редактировать ]Примечания
[ редактировать ]Цитаты
[ редактировать ]- ^ Перейти обратно: а б BirdLife International (2016). « Тахицинета биколор » . Красный список видов, находящихся под угрозой исчезновения МСОП . 2016 : e.T22712057A94316797. doi : 10.2305/IUCN.UK.2016-3.RLTS.T22712057A94316797.en . Проверено 12 ноября 2021 г.
- ^ Вьейо, Луи Пьер (1807). Естественная история птиц Северной Америки: содержит большое количество видов, описанных или изображенных впервые (на французском языке). Полет. 1. Париж: Крепле. п. 61.
- ^ Кабанис, Жан (1850). Музей Heineanum: Справочник орнитологической коллекции Оберамтмана Фердинанда Гейне в Гут-Сент-Бурхард фон Хальберштадт (на немецком языке). Том 1. Хальберштадт: Независимо по заказу Р. Франца. п. 48.
- ^ Куэс, Эллиотт (1878). Птицы долины Колорадо: хранилище научной и популярной информации по орнитологии Северной Америки . Вашингтон, округ Колумбия: Государственная типография.
- ^ Перейти обратно: а б Куэс, Эллиотт (1882). Контрольный список птиц Северной Америки (2-е изд.). Бостон: Эстес и Лориат. п. 42.
- ^ Тернер, Анджела (2020). дель Ойо, Джозеф; Эллиотт, Эндрю; Саргатал, Джордж; Кристи, Дэвид А.; де Хуана, Эдуардо (ред.). «Белохвостая ласточка ( Tachycineta leucorrhoa )» . Справочник птиц мира живых . Рысь Издания. дои : 10.2173/bow.whrswa1.01 . S2CID 216180193 . Проверено 14 января 2017 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м Тернер, Анджела (2010). Справочник для ласточек и ласточек мира . Лондон: A&C Black. стр. 97–100. ISBN 978-1-4081-3172-5 .
- ^ Джоблинг, Джеймс А. (2010). Словарь научных названий птиц Хелма . Лондон: Кристофер Хелм. стр. 72, 377. ISBN. 978-1-4081-2501-4 .
- ^ Серасале, Дэвид Дж.; Дор, Рой; Винклер, Дэвид В.; Ловетт, Ирби Дж. (2012). «Филогения рода Tachycineta ласточек Нового Света: данные полных митохондриальных геномов» (PDF) . Энергетический баланс у перелетных птиц: данные митохондриального генома и биологии рецепторов лептина (доктор философии). Корнелльский университет. стр. 91–122.
- ^ Уиттингем, Линда А; Сликас, Бет; Винклер, Дэвид В.; Шелдон, Фредерик Х (2002). «Филогения рода древесных ласточек Tachycineta (Aves: Hirundinidae) на основе байесовского анализа последовательностей митохондриальной ДНК». Молекулярная филогенетика и эволюция . 22 (3): 430–441. дои : 10.1006/mpev.2001.1073 . ISSN 1055-7903 . ПМИД 11884168 .
- ^ Мур, Уильям С. (1995). «Выводы о филогениях на основе вариаций мтДНК: деревья митохондриальных генов и деревья ядерных генов». Эволюция . 49 (4): 718–726. дои : 10.2307/2410325 . ISSN 0014-3820 . JSTOR 2410325 . ПМИД 28565131 .
- ^ Дор, Рой; Карлинг, Мэтью Д.; Ловетт, Ирби Дж.; Шелдон, Фредерик Х.; Винклер, Дэвид В. (2012). «Деревья видов древесных ласточек (род Tachycineta ): альтернативная филогенетическая гипотеза митохондриальному генному дереву». Молекулярная филогенетика и эволюция . 65 (1): 317–322. дои : 10.1016/j.ympev.2012.06.020 . ISSN 1055-7903 . ПМИД 22750631 .
- ^ «Идентификация древесных ласточек, Все о птицах, Орнитологическая лаборатория Корнелла» . www.allaboutbirds.org . Проверено 28 сентября 2020 г.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С Тернер, А. (2020). дель Ойо, Хосеп; Эллиотт, Эндрю; Саргатал, Джорди; Кристи, Дэвид А.; де Хуана, Эдуардо (ред.). «Древесная ласточка ( Tachycineta bicolor )» . Справочник птиц мира живых . Барселона, Испания: Lynx Edicions. дои : 10.2173/bow.treswa.01 . S2CID 216382529 . Проверено 10 декабря 2017 г.
- ^ Перейти обратно: а б Хассел, Дэвид Дж. Т. (1983). «Возраст и цвет оперения самок древесных ласточек». Журнал полевой орнитологии . 54 (3): 312–318.
- ^ Статчбери, Бриджит Дж.; Робертсон, Рэли Дж. (октябрь 1987 г.). «Сигнализация подчиненного статуса и статуса самки: две гипотезы об адаптивном значении подросткового оперения самок древесных ласточек» (PDF) . Аук . 104 (4): 717–723. дои : 10.1093/аук/104.4.717 . ISSN 0004-8038 .
- ^ Коуди, Челси Д.; Доусон, Рассел Д. (2013). «Цвет оперения полувзрослых самок древесных ласточек ( Tachycineta bicolor ) снижает агрессию сородичей в период размножения». Орнитологический журнал Уилсона . 125 (2): 348–357. дои : 10.1676/12-155.1 . ISSN 1559-4491 . S2CID 85030082 .
- ^ Перейти обратно: а б с Шарман, штат Миссури; Робертсон, Р.Дж.; Рэтклифф, LM (1994). «Вокализации древесной ласточки ( Tachycineta bicolor ) в период перед кладкой: структурный и контекстуальный анализ». Американский натуралист из Мидленда . 132 (2): 264–274. дои : 10.2307/2426582 . ISSN 0003-0031 . JSTOR 2426582 .
- ^ Перейти обратно: а б Леонард, Марти Л.; Фернандес, Николь; Браун, Глен (1997). «Родительские крики и птенцовое поведение древесных ласточек» . Аук . 114 (4): 668–672. дои : 10.2307/4089286 . ISSN 0004-8038 . JSTOR 4089286 .
- ^ Перейти обратно: а б с д и Винклер, Дэвид В. (1992). «Причины и последствия изменения защитного поведения родителей древесных ласточек». Кондор . 94 (2): 502–520. дои : 10.2307/1369222 . ISSN 0010-5422 . JSTOR 1369222 . S2CID 45389696 .
- ^ Макинтайр, Эмма; Хорн, Эндрю Г.; Леонард, Марти Л. (2014). «Реагируют ли птенцы древесных ласточек ( Tachycineta bicolor ) на сигналы тревоги родителей?» . Аук . 131 (3): 314–320. дои : 10.1642/AUK-13-235.1 . ISSN 0004-8038 . S2CID 86754716 .
- ^ Макинтайр, Эмма; Леонард, Марти Л.; Хорн, Эндрю Г. (2014). «Окружающий шум и сообщение родителей о риске нападения хищников у древесных ласточек Tachycineta bicolor ». Поведение животных . 87 : 85–89. дои : 10.1016/j.anbehav.2013.10.013 . ISSN 0003-3472 . S2CID 53173370 .
- ^ Леонард, Марти Л.; Хорн, Эндрю Г.; Освальд, Криста Н.; Макинтайр, Эмма (2015). «Влияние окружающего шума на взаимодействие родителей и потомков у древесных ласточек». Поведение животных . 109 : 1–7. дои : 10.1016/j.anbehav.2015.07.036 . ISSN 0003-3472 . S2CID 53160897 .
- ^ Найт, Саманта М.; Брэдли, Дэвид В.; Кларк, Роберт Г.; Гоу, Элизабет А.; Белиль, Марк; Берзиньш, Лиша Л.; Блейк, Триша; Бридж, Эли С.; Берк, Лорен; Доусон, Рассел Д.; Данн, Питер О.; Гарант, Дэни; Холройд, Джеффри Л.; Хассел, Дэвид Дж.Т.; Лансдорп, Ольга; Лафлин, Эндрю Дж.; Леонард, Марти Л.; Пеллетье, Фани; Шатлер, Дэйв; Зифферман, Линн; Тейлор, Кэз М.; Трефри, Хелен Э.; Влек, Кэрол М.; Влек, Дэвид; Винклер, Дэвид В.; Уиттингем, Линда А.; Норрис, Д. Райан (2018). «Построение и оценка общеконтинентальной сети перелетных певчих птиц в течение годового цикла» . Экологические монографии . 88 (3): 445–460. дои : 10.1002/ecm.1298 . hdl : 10214/11411 . ISSN 0012-9615 .
- ^ Перейти обратно: а б Винклер, Дэвид В.; Реге, Питер Х.; Аллен, Пол Э.; Каст, Трейси Л.; Сенесак, Пикси; Уоссон, Мэтью Ф.; Льямбиас, Пауло Э.; Ферретти, Валентина; Салливан, Патрик Дж. (2004). «Рассредоточение и филопатрия древесной ласточки». Кондор (Представлена рукопись). 106 (4): 768–776. дои : 10.1650/7634 . ISSN 0010-5422 . S2CID 42958338 .
- ^ Винклер, Дэвид В. (2006). «Насесты и миграции ласточек» . Эль Хорнеро . 21 (2): 85–97. дои : 10.56178/eh.v21i2.790 . hdl : 20.500.12110/hornero_v021_n02_p085 .
- ^ Джонс, Джейсон (2003). «Ласточки ( Tachycineta bicolor ): новый модельный организм?». Аук . 120 (3): 591. doi : 10.1642/0004-8038(2003)120[0591:TSTBAN]2.0.CO;2 . ISSN 0004-8038 . S2CID 36003514 .
- ^ Шатлер, Дэйв; Кларк, Роберт Г. (2003). «Причины и последствия расселения древесной ласточки ( Tachycineta bicolor ) в Саскачеване». Аук . 120 (3): 619–631. doi : 10.1642/0004-8038(2003)120[0619:CACOTS]2.0.CO;2 . ISSN 0004-8038 . S2CID 28251342 .
- ^ Мулдал, Элисон; Гиббс, Х. Лайл; Робертсон, Рэли Дж. (1985). «Предпочтительное расположение гнезда облигатной птицы, гнездящейся в дуплах, древесной ласточки». Кондор . 87 (3): 356–363. дои : 10.2307/1367216 . ISSN 0010-5422 . JSTOR 1367216 .
- ^ Ренделл, Уоллес Б.; Робертсон, Рэли Дж. (1989). «Характеристики гнезда, репродуктивный успех и наличие полостей для древесных ласточек, размножающихся в естественных полостях». Кондор . 91 (4): 875. дои : 10.2307/1368072 . ISSN 0010-5422 . JSTOR 1368072 .
- ^ Перейти обратно: а б Винклер, Дэвид В. (1993). «Использование и значение перьев в качестве подкладки гнезда у древесных ласточек ( Tachycineta bicolor )». Аук . 110 (1): 29–36.
- ^ Ломбардо, Майкл П.; Босман, Рут М.; Фаро, Кристин А.; Хауттеман, Стивен Г.; Клуиза, Тимоти С. (1995). «Влияние перьев как изоляции гнезда на инкубационное поведение и репродуктивную способность древесных ласточек ( Tachycineta bicolor )» . Аук . 112 (4): 973–981. дои : 10.2307/4089028 . ISSN 0004-8038 . JSTOR 4089028 . S2CID 55119375 .
- ^ Виндзор, Ребекка Л.; Фегели, Джессика Л.; Ардия, Дэниел Р. (2013). «Влияние размера гнезда и изоляции на тепловые свойства ласточкиных гнезд». Журнал птичьей биологии . 44 (4): 305–310. дои : 10.1111/j.1600-048X.2013.05768.x . ISSN 0908-8857 .
- ^ Холланд, Эрика Р.; Шатлер, Дэйв (2018). «Реакция оперения гнезд древесных ласточек ( Tachycineta bicolor ) на экспериментальное потепление». Журнал орнитологии . 159 (4): 991–998. дои : 10.1007/s10336-018-1568-6 . ISSN 2193-7192 . S2CID 46971093 .
- ^ Данн, ПО; Винклер, Д.В. (1999). «Изменение климата повлияло на сроки размножения древесных ласточек по всей Северной Америке» . Труды Королевского общества B: Биологические науки . 266 (1437): 2487–2490. дои : 10.1098/rspb.1999.0950 . ISSN 0962-8452 . ПМК 1690485 . ПМИД 10693819 .
- ^ Перейти обратно: а б Нукер, Жаклин К.; Данн, Питер О.; Уиттингем, Линда А. (2005). «Влияние изобилия пищи, погоды и состояния самок на размножение древесных ласточек ( Tachycineta bicolor )» . Аук . 122 (4): 1225. doi : 10.1642/0004-8038(2005)122[1225:EOFAWA]2.0.CO;2 . ISSN 0004-8038 . S2CID 4995233 .
- ^ Винклер, Дэвид В.; Аллен, Пол Э. (1996). «Сезонное уменьшение размера кладки ласточек деревьев: физиологическое ограничение или стратегическая корректировка?». Экология . 77 (3): 922–932. дои : 10.2307/2265512 . ISSN 0012-9658 . JSTOR 2265512 . S2CID 84919928 .
- ^ Данн, Питер О.; Хэннон, Сьюзен Дж. (1991). «Внутривидовая конкуренция и поддержание моногамии у древесных ласточек». Поведенческая экология . 2 (3): 258–266. дои : 10.1093/beheco/2.3.258 . ISSN 1045-2249 .
- ^ Данн, Питер О.; Хэннон, Сьюзен Дж. (1992). «Влияние изобилия пищи и родительской заботы самцов на репродуктивный успех и моногамию древесных ласточек». Аук . 109 (3): 488–499. ISSN 0004-8038 .
- ^ Перейти обратно: а б Лифьельд, Ян Т.; Данн, Питер О.; Робертсон, Рэли Дж.; Боаг, Питер Т. (1993). «Внепарное отцовство у моногамных древесных ласточек». Поведение животных . 45 (2): 213–229. дои : 10.1006/anbe.1993.1028 . ISSN 0003-3472 . S2CID 53195131 .
- ^ Кроу, Сьюзен А.; Клевен, Одмунд; Делмор, Кира Э.; Ласкемоен, Терье; Ночера, Джозеф Дж.; Лифьельд, Ян Т.; Робертсон, Рэли Дж. (2009). «Утверждение отцовства посредством частых совокуплений диких воробьиных с интенсивной конкуренцией сперматозоидов». Поведение животных . 77 (1): 183–187. дои : 10.1016/j.anbehav.2008.09.024 . ISSN 0003-3472 . S2CID 13722818 .
- ^ Кемпенаерс, Барт; Ланкто, Ричард Б.; Робертсон, Рэли Дж. (1998). «Уверенность в отцовстве и отцовские инвестиции в восточных синих птиц и древесных ласточек». Поведение животных . 55 (4): 845–860. дои : 10.1006/anbe.1997.0667 . ISSN 0003-3472 . ПМИД 9632472 . S2CID 24760456 .
- ^ Кемпенаерс, Барт; Эвердинг, Сьюзи; Бишоп, Шерил; Боаг, Питер; Робертсон, Рэли Дж. (2001). «Внепарное отцовство и репродуктивная роль самцов древесной ласточки ( Tachycineta bicolor )». Поведенческая экология и социобиология . 49 (4): 251–259. дои : 10.1007/s002650000305 . ISSN 0340-5443 . S2CID 25483760 .
- ^ Статчбери, Бриджит Дж.; Робертсон, Рэли Дж. (1985). «Плавающие популяции самок древесных ласточек». Аук . 102 (3): 651–654. дои : 10.1093/аук/102.3.651 . ISSN 0004-8038 .
- ^ Стэплтон, Мэри К.; Клевен, Одмунд; Лифьельд, Ян Т.; Робертсон, Рэли Дж. (2007). «Самки древесных ласточек ( Tachycineta bicolor ) увеличивают гетерозиготность потомства за счет внепарного спаривания». Поведенческая экология и социобиология . 61 (11): 1725–1733. дои : 10.1007/s00265-007-0404-4 . ISSN 0340-5443 . S2CID 22513713 .
- ^ Барбер, Коллин А.; Эдвардс, Мэнди Дж.; Робертсон, Рэли Дж. (2005). «Проверка гипотезы генетической совместимости с древесными ласточками Tachycineta bicolor ». Канадский журнал зоологии . 83 (7): 955–961. дои : 10.1139/z05-091 . ISSN 0008-4301 .
- ^ Биттон, Пьер-Поль; О'Брайен, Эрин Л.; Доусон, Рассел Д. (2007). «Яркость оперения и возраст предсказывают успех экстрапарного оплодотворения самцов древесных ласточек Tachycineta bicolor ». Поведение животных . 74 (6): 1777–1784. дои : 10.1016/j.anbehav.2007.03.018 . ISSN 0003-3472 . S2CID 53170228 .
- ^ Халлингер, Келли Кристен (2017). Контекстно-зависимые последствия отцовства внепарной пары у древесных ласточек ( Tachycineta bicolor ) (доктор философии). Корнеллский университет .
- ^ Белмейкер, Амос; Халлингер, Келли К.; Глинн, Ребекка А.; Османн, Марк Ф.; Винклер, Дэвид В. (2018). «Есть ли контекстно-зависимое преимущество спаривания дополнительных пар у древесных ласточек?». Аук . 135 (4): 998–1008. дои : 10.1642/AUK-18-3.1 . hdl : 10150/631782 . ISSN 0004-8038 . S2CID 91253157 .
- ^ Перейти обратно: а б с Арнквист, Йоран; Киркпатрик, Марк (2005). «Эволюция неверности у социально моногамных воробьинообразных: сила прямого и косвенного отбора на внепарное совокупительное поведение самок». Американский натуралист . 165 (С5): С26–С37. дои : 10.1086/429350 . ISSN 0003-0147 . ПМИД 15795859 . S2CID 32810854 .
- ^ Данн, Питер О.; Уиттингем, Линда А. (2006). «Затраты на поиск влияют на пространственное распределение, но не на уровень спаривания дополнительных пар у древесных ласточек». Поведенческая экология и социобиология . 61 (3): 449–454. дои : 10.1007/s00265-006-0272-3 . ISSN 0340-5443 . S2CID 4975256 .
- ^ Форстмайер, Вольфганг; Накагава, Шиничи; Гриффит, Саймон К.; Кемпенаерс, Барт (2014). «Внепарное спаривание самок: адаптация или генетическое ограничение?». Тенденции в экологии и эволюции . 29 (8): 456–464. дои : 10.1016/j.tree.2014.05.005 . ISSN 0169-5347 . ПМИД 24909948 .
- ^ Перейти обратно: а б Хаубер, Марк Э. (1 августа 2014 г.). Книга яиц: путеводитель по яйцам шестисот видов птиц мира в натуральную величину . Чикаго: Издательство Чикагского университета. п. 448. ИСБН 978-0-226-05781-1 .
- ^ Перейти обратно: а б Клотфелтер, Итан Д.; Уиттингем, Линда А.; Данн, Питер О. (2000). «Порядок яйцекладки, асинхронность вылупления и масса тела птенцов у древесных ласточек Tachycineta bicolor ». Журнал птичьей биологии . 31 (3): 329–334. дои : 10.1034/j.1600-048X.2000.310308.x . ISSN 0908-8857 .
- ^ Робертсон, Рэли Дж. (1990). «Тактика и контртактика детоубийства древесных ласточек, отобранных половым путем». Популяционная биология воробьиных птиц: комплексный подход . Серия НАТО ASI. стр. 381–390. дои : 10.1007/978-3-642-75110-3_32 . ISBN 978-3-540-51759-7 .
- ^ Уиттингем, Линда А.; Данн, Питер О. (2000). «Соотношение полов в потомстве древесных ласточек: самки в лучшем состоянии рожают больше сыновей». Молекулярная экология . 9 (8): 1123–1129. дои : 10.1046/j.1365-294x.2000.00980.x . ISSN 0962-1083 . ПМИД 10964231 . S2CID 3720108 .
- ^ Маккарти, Джон П.; Винклер, Дэвид В. (2008). «Относительная важность переменных окружающей среды в определении роста птенцов ласточки Tachycineta bicolor ». Ибис . 141 (2): 286–296. дои : 10.1111/j.1474-919X.1999.tb07551.x . ISSN 0019-1019 .
- ^ Данн, Эрика Х. (1979). «Возраст эффективной гомеотермии у птенцов древесных ласточек в зависимости от размера выводка». Бюллетень Уилсона . 91 (3): 455–457. ISSN 0043-5643 .
- ^ Де Стивен, Дайан (1980). «Размер кладки, успех размножения и выживание родителей древовидной ласточки ( Iridoprocne bicolor )». Эволюция . 34 (2): 278–291. дои : 10.1111/j.1558-5646.1980.tb04816.x . hdl : 2027.42/137562 . ISSN 0014-3820 . ПМИД 28563429 . S2CID 19454000 .
- ^ Робертсон, Рэли Дж.; Ренделл, Уоллес Б. (1990). «Сравнение экологии размножения птицы, гнездящейся во вторичных дуплах, ласточки ( Tachycineta bicolor ) в гнездовых ящиках и естественных полостях». Канадский журнал зоологии . 68 (5): 1046–1052. дои : 10.1139/z90-152 . ISSN 0008-4301 .
- ^ Лич, Сьюзен М.; Леонард, Марти Л. (1997). «Попрошайничество и опасность нападения хищников на птенцов птиц» . Поведенческая экология . 8 (6): 644–646. дои : 10.1093/beheco/8.6.644 . ISSN 1045-2249 .
- ^ Перейти обратно: а б Маккарти, Джон П.; Винклер, Дэвид В. (1999). «Экология кормодобывания и избирательность питания древесных ласточек, выкармливающих птенцов» . Кондор . 101 (2): 246–254. дои : 10.2307/1369987 . ISSN 0010-5422 . JSTOR 1369987 .
- ^ Твининг, Корнелия В.; Лоуренс, Питер; Винклер, Дэвид В.; Флекер, Александр С.; Бренна, Дж. Томас (2018). «Эффективность преобразования α-линоленовой кислоты в высоконенасыщенные жирные кислоты омега-3 у воздушных насекомоядных цыплят» . Журнал экспериментальной биологии . 221 (3): jeb165373. дои : 10.1242/jeb.165373 . ISSN 0022-0949 . ПМИД 29217628 .
- ^ Леонард, Марти Л.; Хорн, Эндрю Г. (2001). «Попрошайничество в отсутствие родителей птенцов древесных ласточек» . Поведенческая экология . 12 (4): 501–505. дои : 10.1093/beheco/12.4.501 . ISSN 1465-7279 .
- ^ Джонсон, Л. Скотт; Виммерс, Ларри Э.; Кэмпбелл, Сара; Гамильтон, Люси (2003). «Скорость роста, размер и соотношение полов последнего отложенного и последнего вылупившегося потомства древесной ласточки Tachycineta bicolor ». Журнал птичьей биологии . 34 (1): 35–43. дои : 10.1034/j.1600-048X.2003.02950.x . ISSN 0908-8857 .
- ^ Уиттингем, Линда А; Данн, Питер О; Клотфелтер, Итан Д. (2003). «Родительское распределение пищи птенцам древесных ласточек: влияние поведения птенца, пола и отцовства». Поведение животных . 65 (6): 1203–1210. дои : 10.1006/anbe.2003.2178 . ISSN 0003-3472 . S2CID 4993059 .
- ^ Леонард, ML; Хорн, Эндрю Г. (2001). «Призывы к попрошайничеству и решения родителей о кормлении древесных ласточек ( Tachycineta bicolor )». Поведенческая экология и социобиология . 49 (2–3): 170–175. дои : 10.1007/s002650000290 . ISSN 0340-5443 . S2CID 25507198 .
- ^ Перейти обратно: а б Сент-Луис, Винсент Л.; Брибарт, Лоес (1991). «Добавки кальция в рационе птенцов древесных ласточек вблизи чувствительных к кислоте озер». Кондор . 93 (2): 286–294. дои : 10.2307/1368944 . ISSN 0010-5422 . JSTOR 1368944 . S2CID 87271779 .
- ^ Влек, Кэрол М.; Влек, Дэвид; Паласиос, Мария Г. (2011). «Эволюционная экология старения: пример использования древесных ласточек Tachycineta bicolor ». Журнал орнитологии . 152 (С1): 203–211. дои : 10.1007/s10336-010-0629-2 . ISSN 0021-8375 . S2CID 25321642 .
- ^ Османн, Марк Ф.; Винклер, Дэвид В.; Влек, Кэрол М. (22 июня 2005 г.). «Более длинные теломеры связаны с более высокой выживаемостью птиц» . Письма по биологии . 1 (2): 212–214. дои : 10.1098/rsbl.2005.0301 . ISSN 1744-9561 . ПМЦ 1626238 . ПМИД 17148169 .
- ^ Белмейкер, Амос (2016). «Закономерности ковариации длины теломер, состояния, истории жизни и приспособленности у недолговечных видов птиц». Роль длины теломер в поведении и истории жизни древесной ласточки (доктор философии). Корнелльский университет.
- ^ Белмейкер, Амос; Халлингер, Келли К.; Глинн, Реббека А.; Винклер, Дэвид В.; Османн, Марк Ф. (2019). «Экологические и генетические детерминанты длины теломер цыпленка у древесных ласточек ( Tachycineta bicolor )» . Экология и эволюция . 9 (14): 8175–8186. дои : 10.1002/ece3.5386 . ISSN 2045-7758 . ПМК 6662556 . ПМИД 31380080 .
- ^ Перейти обратно: а б с д и Винклер, Д.В.; Халлингер, К.К.; Ардия, ДР; Робертсон, Р.Дж.; Статчбери, Би Джей; Чоэн, Р.Р. (2011). Пул, А.Ф., изд. «Древесная ласточка ( Tachycineta bicolor )». Птицы Северной Америки . Итака, Нью-Йорк : Корнельская лаборатория орнитологии.
- ^ Зак, Рето; Мэйо, Кейт Р. (1984). «Воздействие гамма-излучения на птенцов древесных ласточек». Экология . 65 (5): 1641–1647. дои : 10.2307/1939142 . ISSN 0012-9658 . JSTOR 1939142 .
- ^ Чепмен, LB (1955). «Исследования колонии древесных ласточек». Кольцевание птиц . 6 (2): 45–70.
- ^ Эррингтон, Польша (1932). «Пищевые привычки хищников южного Висконсина. Часть I. Совы». Кондор . 34 (4): 176–186.
- ^ Перейти обратно: а б Маасс, Наталья Мэй (2019). Воспринимаемый риск нападения хищников и реакция взрослых и птенцов древесных ласточек ( Tachycineta bicolor ) ( MS ). Университет Восточного Кентукки .
- ^ Шатлер, Дэйв; Мулли, Адель; Кларк, Роберт Дж. (2004). «Дерево глотает репродуктивные инвестиции, стресс и паразиты». Канадский журнал зоологии . 82 (3): 442–448. CiteSeerX 10.1.1.530.1736 . дои : 10.1139/z04-016 . ISSN 0008-4301 .
- ^ Перейти обратно: а б Роби, Дэниел Д.; Бринк, Карен Л.; Виттманн, Карин (1992). «Влияние паразитизма птичьей мясной мухи на птенцов восточной синей птицы и древесных ласточек» (PDF) . Бюллетень Уилсона . 104 (4): 630–643.
- ^ Дезимоун, Джоэли Дж.; Клотфелтер, Итан Д.; Блэк, Элизабет К.; Кнути, Сара А. (2018). «Избегание, толерантность и устойчивость к эктопаразитам у птенцов и взрослых древесных ласточек». Журнал птичьей биологии . 49 (2): jav–01641. дои : 10.1111/jav.01641 . ISSN 0908-8857 .
- ^ Хассельквист, Деннис; Уоссон, Мэтью Ф.; Винклер, Дэвид В. (2001). «Гуморальная иммунокомпетентность коррелирует с датой откладки яиц и отражает рабочую нагрузку самок древесных ласточек» . Поведенческая экология . 12 (1): 93–97. doi : 10.1093/oxfordjournals.beheco.a000384 . ISSN 1465-7279 .
- ^ Ардия, Дэниел Р. (2005). «Индивидуальные качества опосредуют компромисс между репродуктивными усилиями и иммунной функцией древесных ласточек» . Журнал экологии животных . 74 (3): 517–524. дои : 10.1111/j.1365-2656.2005.00950.x . ISSN 0021-8790 .
- ^ Ардия, Дэниел Р. (2005). «Древесные ласточки по-разному распределяют иммунную функцию и репродуктивные усилия в зависимости от своего ареала». Экология . 86 (8): 2040–2046. дои : 10.1890/04-1619 . ISSN 0012-9658 . S2CID 33354318 .
- ^ Паласиос, Мария Дж; Кунник, Джоан Э; Винклер, Дэвид В.; Влек, Кэрол М (2007). «Иммуностарение некоторых, но не всех иммунных компонентов у свободноживущих позвоночных, древесной ласточки» . Труды Королевского общества B: Биологические науки . 274 (1612): 951–957. дои : 10.1098/рспб.2006.0192 . ISSN 0962-8452 . ПМК 2141670 . ПМИД 17251097 .
- ^ Паласиос, Мария Г.; Винклер, Дэвид В.; Класинг, Кирк К.; Хассельквист, Деннис; Влек, Кэрол М. (2011). «Последствия старения иммунной системы в природе: исследование затрат на иммуностарение у свободноживущих древесных ласточек» . Экология (Представлена рукопись). 92 (4): 952–966. дои : 10.1890/10-0662.1 . ISSN 0012-9658 . ПМИД 21661557 .
- ^ «Охраняемые виды Закона о перелетных птицах (список 10.13)» . Служба рыболовства и дикой природы США. 2013 . Проверено 4 июня 2018 г.
- ^ «Птицы, охраняемые Законом о Конвенции о перелетных птицах» . Правительство Канады. 2017. Архивировано из оригинала 20 мая 2019 года . Проверено 21 июля 2018 г.
- ^ Вибе, Карен Л. (2016). «Межвидовая конкуренция за гнезда: прежнее владение превосходит потенциал владения ресурсами для горной синей птицы, конкурирующей с древесной ласточкой» . Аук . 133 (3): 512–519. дои : 10.1642/AUK-16-25.1 . ISSN 0004-8038 .
- ^ Финч, Дебора М. (1990). «Влияние хищников и вмешательства конкурентов на успех гнездования крапивников и древесных ласточек». Кондор . 92 (3): 674–687. дои : 10.2307/1368686 . ISSN 0010-5422 . JSTOR 1368686 .
- ^ Смитс, Юдит Э.; Вэйланд, Марк Э.; Миллер, Майкл Дж.; Либер, Карстен; Трюдо, Сюзанна (2000). «Репродуктивные, иммунные и физиологические показатели у древесных ласточек на участках добычи нефтеносных песков». Экологическая токсикология и химия . 19 (12): 2951–2960. дои : 10.1002/etc.5620191216 . ISSN 0730-7268 . S2CID 85573498 .
- ^ Гентес, Мари-Лин; Уолднер, Шерил; Папп, Жужанна; Смитс, Юдит Э.Г. (2006). «Влияние соединений хвостохранилищ нефтеносных песков и суровых погодных условий на уровень смертности, рост и усилия по детоксикации птенцов древесных ласточек ( Tachycineta bicolor )». Загрязнение окружающей среды . 142 (1): 24–33. дои : 10.1016/j.envpol.2005.09.013 . ISSN 0269-7491 . ПМИД 16297515 .
- ^ Годвин, Кристин М.; Барклай, Роберт М.Р.; Смитс, Юдит Э.Г. (2019). «Успех гнездования древесной ласточки ( Tachycineta bicolor ) и рост птенцов возле месторождений нефтеносных песков на северо-востоке Альберты, Канада» . Канадский журнал зоологии . 97 (6): 547–557. дои : 10.1139/cjz-2018-0247 . ISSN 0008-4301 .
- ^ Халлингер, Келли К.; Кристол, Дэниел А. (2011). «Роль погоды в воздействии ртути на репродуктивный успех древесных ласточек». Экотоксикология . 20 (6): 1368–1377. дои : 10.1007/s10646-011-0694-1 . ISSN 0963-9292 . ПМИД 21553259 . S2CID 42492417 .
- ^ Винклер, Дэвид В.; Луо, Майлз К.; Рахимбердиев, Эльдар (сентябрь 2013 г.). «Влияние температуры на запасы пищи и смертность птенцов древесных ласточек (Tachycineta bicolor)» . Экология . 173 (1): 129–138. Бибкод : 2013Oecol.173..129W . дои : 10.1007/s00442-013-2605-z . ISSN 0029-8549 . ПМЦ 3751296 . ПМИД 23468236 .
- ^ Шипли, Дж. Райан; Твининг, Корнелия В.; Тафф, Конор К.; Витоусек, Марен Н.; Флэк, Андреа; Винклер, Дэвид В. (28 сентября 2020 г.). «Птицы, приближающиеся к яйцеклетке из-за потепления весны, сталкиваются с большим риском смертности птенцов» . Труды Национальной академии наук . 117 (41): 25590–25594. Бибкод : 2020PNAS..11725590S . дои : 10.1073/pnas.2009864117 . ISSN 0027-8424 . ПМЦ 7568286 . ПМИД 32989166 . S2CID 222159036 .
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|