Парацератерий
| Парацератерий Временной диапазон: олигоцен , | |
|---|---|
 | |
| Установленный P. transouralicum скелет , Московский Палеонтологический музей ; это наиболее полностью известный скелет, но череп представляет собой слепок экземпляра из Американского музея естественной истории. [1] | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Периссодактилия |
| Семья: | † Парацератерииды |
| Род: | † Парацератерий Форстер-Купер , 1911 год. |
| Типовой вид | |
| † Aceratherium bugtiense Пилигрим , 1908 год. | |
| Разновидность | |
| |
| Синонимы | |
Синонимия рода Синонимия видов | |
Paraceratherium — вымерший род безрогих носорогов, принадлежащих к семейству Paraceratheriidae . Это одно из крупнейших наземных млекопитающих , когда-либо существовавших и жившее с эпохи раннего до позднего олигоцена (34–23 миллиона лет назад). Первые окаменелости были обнаружены на территории нынешнего Пакистана, а останки были найдены по всей Евразии между Китаем и Балканами . Paraceratherium означает «около безрогого зверя» в отношении Aceratherium , рода, к которому первоначально был отнесен типовой вид P. bugtiense .
Точный размер Paraceratherium неизвестен из-за неполноты окаменелостей . Высота в плечах составляла около 4,8 метра (15,7 фута), а длина около 7,4 метра (24,3 фута). По оценкам, его вес составлял от 15 до 20 тонн (от 33 000 до 44 000 фунтов). Длинная шея поддерживала череп длиной около 1,3 метра (4,3 фута). У него были большие, похожие на бивни резцы и носовой разрез, который позволяет предположить, что у него была цепкая верхняя губа или хоботок (туловище). Ноги были длинными и похожими на столбы. Образ жизни Paraceratherium , возможно, был похож на образ жизни современных крупных млекопитающих, таких как слоны и современные носороги. Из-за своего размера у него было мало хищников и длительный период беременности . Это был браузер , питавшийся в основном листьями, мягкими растениями и кустарниками. Он обитал в самых разных средах обитания: от засушливых пустынь с редкими деревьями до субтропических лесов. Причины исчезновения животного неизвестны, но были предложены различные факторы.
Таксономия . рода и видов внутри него имеет долгую и сложную историю Другие роды олигоценовых индрикотериев, такие как Baluchitherium , Indricotherium и Pristinotherium , были названы, но полных экземпляров не существует, что затрудняет сравнение и классификацию. Большинство современных ученых считают эти роды младшими синонимами Paraceratherium ; и, как полагают, содержат следующие виды P. bugtiense , P. transouralicum , P. huangheense и P. linxiaense . Наиболее изучен вид P. transouralicum , поэтому на его основе основано большинство реконструкций рода. Различия между P. bugtiense и P. transouralicum могут быть связаны с половым диморфизмом , который делает их одним и тем же видом.
Таксономия
[ редактировать ]Таксономическая советские история Paraceratherium сложна из-за фрагментарного характера известных окаменелостей, а также из-за того, что западные, и китайские ученые работали изолированно друг от друга на протяжении большей части 20-го века и публиковали исследования в основном на своих языках. [1] Ученые из разных уголков мира пытались сравнить свои находки, чтобы получить более полное представление об этих животных, но им помешали политика и войны. [2] Противоположные таксономические тенденции « объединения и расщепления » также усугубили проблему. [3] Неточные геологические датировки ранее заставляли ученых полагать, что различные геологические образования , которые, как теперь известно, являются одновременными, имели разный возраст. Многие роды были названы на основе тонких различий в характеристиках коренных зубов — особенностях, которые варьируются в пределах популяций других таксонов носорогов — и поэтому не принимаются большинством ученых для различения видов. [4]

Ранние открытия индрикотериев были сделаны благодаря различным колониальным связям с Азией. [2] Первые известные окаменелости индрикотеров были собраны в Белуджистане (на территории современного Пакистана) в 1846 году солдатом по имени Викари, но в то время эти фрагменты было невозможно идентифицировать. [5] Первые окаменелости, ныне признанные Paraceratherium, были обнаружены британским геологом Гаем Эллкоком Пилигримом в Белуджистане в 1907–1908 годах. Его материал состоял из верхней челюсти, нижних зубов и задней части челюсти. Окаменелости были собраны в формации Читарвата в Дера Бугти , где ранее исследовал Пилигрим. В 1908 году он использовал окаменелости в качестве основы для нового вида вымершего рода носорогов Aceratherium ; А. бутьенсе . К тому времени Aceratherium был таксоном мусорной корзины ; он включал несколько неродственных видов безрогих носорогов, многие из которых с тех пор были перенесены в другие роды. [1] [6] Ископаемые резцы , которые Пилигрим ранее отнес к несвязанному роду Bugtitherium, позже выяснилось, что они принадлежат к новому виду. [7]
В 1910 году в Дера Бугти во время экспедиции британского палеонтолога Клайва Форстера-Купера были обнаружены еще частичные окаменелости . На основании этих останков Фостер-Купер перенес A. bugtiense в новый род Paraceratherium , что означает «около безрогого зверя», применительно к Aceratherium . [1] [8] Его обоснованием для этой реклассификации послужили явно повернутые вниз нижние бивни этого вида. [7] В 1913 году Форстер-Купер назвал новый род и вид Thaumastotherium («чудесный зверь») osborni , основываясь на более крупных окаменелостях из тех же раскопок (некоторые из которых он ранее предполагал принадлежать самцам P. bugtiense ), но переименовал род Baluchitherium позже в том же году, потому что прежнее название было озабочено , поскольку оно уже использовалось для полужесткокрылых насекомых. [9] [10] [8] Окаменелости Baluchitherium были настолько фрагментарны, что Фостер-Купер смог идентифицировать его лишь как разновидность непарнокопытного копытного , однако он упомянул о возможности путаницы с Paraceratherium . [11] Американский палеонтолог Генри Фэйрфилд Осборн , в честь которого был назван B. osborni , предположил, что это мог быть титанотерий . [2]
Позже экспедиция Российской академии наук обнаружила окаменелости в аральской формации возле Аральского моря в Казахстане; это был самый полный из известных скелетов индрикотеров, но у него не было черепа. Он установлен в Московском Палеонтологическом музее . В 1916 году на основе этих останков Алексей Алексеевич Борисяк выделил род Indricotherium, названный по имени мифологического чудовища — « Индрикского зверя ». Он не присвоил видовое название I. asiaticum до 1923 года, но русский палеонтолог Мария Павлова уже назвала его I. transouralicum в 1922 году. [1] [12] Также в 1923 году Бориссяк создал подсемейство Indricotheriinae, включив в него различные родственные формы, известные к тому времени. [13]
В 1922 году американский исследователь Рой Чепмен Эндрюс возглавил хорошо задокументированную экспедицию в Китай и Монголию , спонсируемую Американским музеем естественной истории . В образованиях монгольской пустыни Гоби были обнаружены различные останки индрикотера , в том числе ноги экземпляра, стоявшего в вертикальном положении, что указывает на то, что он умер, находясь в ловушке в зыбучих песках , а также очень полный череп. Эти останки стали основой Baluchitherium grangeri , названного Осборном в 1923 году. [14] [15]
новый вид, P. huangheense В 2017 году китайский палеонтолог Юн-Сян Ли и его коллеги назвали , на основе элементов челюстей из формации Ханьцзяцзин в провинции Ганьсу в Китае; название относится к близлежащей реке Хуанхэ . [16] В 2021 году китайский палеонтолог Тао Дэн и его коллега описали новый вид P. linxiaense на основе полного черепа с связанной нижней челюстью и атласной костью из формации Цзяоцигоу бассейна Линься (к которому относится название) на северо-западе Китая. [17] Для различных останков индрикотериев было придумано множество других названий видов и родов, в основном основанных на различиях в размере, форме морды и расположении передних зубов. Окаменелости, относящиеся к Paraceratherium, продолжают обнаруживаться по всей Евразии, но политическая ситуация в Пакистане стала слишком нестабильной, чтобы проводить там дальнейшие раскопки. [5]
Виды и синонимы
[ редактировать ]
В 1922 году Форстер-Купер назвал новый вид Metamynodon bugtiensis на основе неба и других фрагментов Дера Бугти, которые, как считалось, принадлежали гигантскому представителю этого рода. Сейчас считается, что эти окаменелости принадлежали аберрантному экземпляру Paraceratherium bugtiense, у которого отсутствовал коренной зуб M3. [18] [19] В 1936 году американские палеонтологи Уолтер Грейнджер и Уильям К. Грегори Форстера-Купера, предположили, что Baluchitherium osborni вероятно, был младшим синонимом (недопустимым названием для того же таксона) Paraceratherium bugtiense , поскольку эти экземпляры были собраны в одном и том же месте и, возможно, были часть одного и того же морфологически изменчивого вида. [20] Американский палеонтолог Уильям Диллер Мэтью и сам Форстер-Купер выразили аналогичные сомнения несколькими годами ранее. Хотя оно уже было объявлено младшим синонимом, название рода Baluchitherium Осборна оставалось популярным в различных средствах массовой информации из-за шумихи вокруг B. grangeri . [3] [21]
В 1989 году американские палеонтологи Спенсер Г. Лукас и Джей К. Собус опубликовали пересмотренную версию таксонов индрикотериев, которой впоследствии последовали западные ученые. Они пришли к выводу, что Paraceratherium , как старейшее название, был единственным действительным родом индрикотериев из олигоцена и содержал четыре действительных вида: P. bugtiense , P. transouralicum (первоначально в Indricotherium ), P. prohorovi (первоначально в Aralotherium ) и P. . orgosensis (первоначально из Dzungariotherium ). Они считали большинство других названий младшими синонимами этих таксонов или сомнительными названиями , основанными на останках, слишком фрагментарных, чтобы их можно было правильно идентифицировать. Анализируя предполагаемые различия между названными родами и видами, Лукас и Собус обнаружили, что они, скорее всего, представляют собой вариации внутри популяций и что большинство особенностей между экземплярами неотличимы, как было отмечено в 1930-х годах. Тот факт, что единственный череп, отнесенный к P. transouralicum или Indricotherium, был куполообразным, в то время как другие были плоскими сверху, приписывают половой диморфизм ; возможно, что окаменелости P. bugtiense представляют самку, а P. transouralicum представляют собой самца того же вида. [3] [22] [23]


По мнению Лукаса и Собуса, типовой вид P. bugtiense из позднего олигоцена Пакистана включал младшие синонимы, такие как B. osborni и P. zhajremensis . P. transouralicum из позднего олигоцена Казахстана, Монголии и северного Китая включала B. grangeri и I. minus . Согласно этой схеме, P. orgosensis из среднего и позднего олигоцена северо-западного Китая включала D. turfanensis и P. Lipidus . [22] В 2013 году американский палеонтолог Дональд Протеро предположил, что P. orgosensis может быть достаточно отличным, чтобы гарантировать его первоначальное название рода Dzungariotherium , хотя его точное положение требует оценки. P. prohorovi из позднего олигоцена Казахстана может оказаться слишком неполным для выяснения его положения по отношению к другим видам; то же самое относится к предлагаемым видам, таким как I. intermedium и P. Tienshanensis , а также к роду Benaratherium . [3] [22] Хотя название рода Indricotherium теперь является младшим синонимом Paraceratherium , название подсемейства Indricotheriinae все еще используется, поскольку синонимия названий родов не влияет на названия таксонов более высокого уровня, которые произошли от них. Поэтому членов подсемейства до сих пор часто называют индрикотериями. [24]
В отличие от редакции Лукаса и Собуса, статья китайского палеонтолога Цзе Йе и его коллег в 2003 году предположила, что Indricotherium и Dzungariotherium были действительными родами, и что P. prohorovi не принадлежал к Paraceratherium . Они также признали ценность таких видов, как P. Lipidus , P. Tienshanensis и P. sui . [25] В статье Дэна и его коллег 2004 года также были выделены три отдельных рода. [26] Некоторые западные авторы аналогичным образом использовали имена, которые в противном случае считались недействительными после редакции 1989 года, но без подробного анализа и обоснования. [3] Дэн и его коллеги признали шесть видов Paraceratherium в 2021 году, включая некоторые, которые ранее были объявлены синонимами, P. grangeri , P. asiaticum и P. lepidum , сохраняя при этом Indricotherium и Baluchitherium как синонимы этого рода. [17] [3]
Эволюция
[ редактировать ]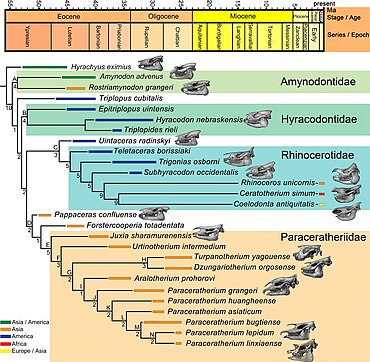
Надсемейство — около Rhinocerotoidea , в которое входят современные носороги, можно проследить до раннего эоцена 50 миллионов лет назад — с ранними предшественниками, такими как Hyrachyus . Rhinocerotoidea включает три семейства; Amynodontidae , Rhinocerotidae («настоящие носороги») и Hyracodontidae . Разнообразие внутри группы носорогов в доисторические времена было намного больше; они варьировались от размеров собаки до размеров Paraceratherium . Существовали длинноногие, бегающие формы, приспособленные для бега, и приземистые, полуводные формы. У большинства видов не было рогов. Окаменелости носорогов идентифицируются как таковые главным образом по характеристикам их зубов, которые, скорее всего, сохранились. Верхние коренные зубы большинства носорогов имеют пи на коронке -образный (π) рисунок, а каждый нижний коренной зуб имеет парную L-образную форму. Различные особенности черепа также используются для идентификации ископаемых носорогов. [27]
Подсемейство Indricotheriinae, к которому принадлежит Paraceratherium , было впервые классифицировано как часть семейства Hyracodontidae американским палеонтологом Леонардом Б. Радински в 1966 году. Ранее они считались подсемейством Rhinocerotidea или даже полным семейством Indricotheriidae. [28] В кладистическом исследовании тапироморфов в 1999 году американский палеонтолог Люк Холбрук обнаружил, что индрикотерии находятся за пределами клады гиракодонтид , и написал, что они не могут быть монофилетической (естественной) группой. [29] Схема Радинского сегодня является распространенной гипотезой. В семействе гиракодонтов имеются длинноногие представители, приспособленные к бегу, такие как гиракодон , и отличавшиеся характеристиками резцов. Индрикотерии отличаются от других гиракодонтов более крупными размерами и своеобразным строением морды, резцов и клыков. Самый ранний известный индрикотерий - это Forstercooperia размером с собаку из среднего и позднего эоцена западной части Северной Америки и Азии. размером с корову Джуксия известна со среднего эоцена; к позднему эоцену род Urtinotherium Азии почти достиг размеров Paraceratherium . [22] [27] Сам парацератерий жил в Евразии в период олигоцена , 23–34 миллиона лет назад. [30] Этот род отличается от других индрикотериев своим большим размером, носовым вырезом, который поддерживал бы мускулистую морду, и обращенными вниз предчелюстными костями . [22] Он также потерял вторые и третьи нижние резцы, нижние клыки и нижние первые премоляры . [27]
соответствует Приведенная ниже кладограмма анализу Indricotheriinae, проведенному Лукасом и Собусом в 1989 году, и показывает ближайших родственников Paraceratherium : [22]
| Гиракодонтиды |
| ||||||||||||||||||||||||
Лукас и его коллеги пришли к аналогичным выводам в предыдущем анализе Forstercooperia в 1981 году , в котором они все еще сохраняли Paraceratherium и Indricotherium как отдельные роды. [31] В 2016 году китайские исследователи Хайбинг Ван и его коллеги использовали название Paraceratheriidae для семейства и Paraceratheriine для подсемейства и поместили их за пределы Hyracodontidae. [32] Дэн и его коллеги подтвердили предыдущие исследования своим анализом 2021 года, предполагая, что Juxia произошла из клады, состоящей из Forstercooperia и Pappaceras, 40 миллионов лет назад, в результате чего образовавшаяся популяция превратилась в Urtinotherium в позднем эоцене и Paraceratherium в олигоцене. Эти исследователи не обнаружили, что Hyracodontidae образуют естественную группу, и обнаружили, что Paraceratheriidae ближе к Rhinocerotidae, в отличие от предыдущих исследований. [17]
Описание
[ редактировать ]
Парацератерий — одно из крупнейших известных наземных млекопитающих , когда-либо существовавших, но его точный размер неясен из-за отсутствия полных экземпляров. [4] Общая длина его тела была оценена Грейнджер и Грегори в 1936 году в 8,7 м (28,5 футов) спереди назад и в 7,4 м (24,3 фута) палеонтологом Верой Громовой в 1959 году. [33] но прежняя оценка теперь считается преувеличенной. Вес Paraceratherium был аналогичен весу некоторых вымерших хоботных , при этом самый большой известный полный скелет принадлежал степному мамонту ( Mammuthus trogontherii ). [34] [35] Несмотря на примерно такую же массу, Paraceratherium мог быть выше любого хоботного. [4] Высота его плеча была оценена Грейнджером и Грегори в 5,25 м (17,2 фута) и в 4,8 м (15,7 фута) палеонтологом Грегори С. Полом в 1997 году. [36] В 2013 году палеонтологи Майкл П. Тейлор и Мэтью Дж. Ведель оценили длину шеи от 2 до 2,5 м (от 6,6 до 8,2 футов). [37]

Первоначальные оценки в 30 тонн (66 000 фунтов) теперь считаются преувеличенными; максимум он мог находиться в диапазоне от 15 до 20 тонн (от 33 000 до 44 000 фунтов) и в среднем всего лишь 11 тонн (24 000 фунтов). Расчеты в основном основывались на окаменелостях P. transouralicum , поскольку этот вид известен по наиболее полным останкам. [4] Оценки были основаны на измерениях черепа, зубов и костей конечностей, но известные костные элементы представлены людьми разных размеров, поэтому все реконструкции скелета представляют собой сложную экстраполяцию, что приводит к нескольким диапазонам веса. [34] [38]
Нет никаких указаний на цвет и текстуру кожи животного, поскольку не известны отпечатки кожи или мумии. Большинство реставраций показывают, что кожа существа толстая, складчатая, серая и безволосая, как у современных носорогов. Поскольку волосы сохраняют тепло тела, современные крупные млекопитающие, такие как слоны и носороги, в основном лишены шерсти. Протеро предположил, что, вопреки большинству изображений, у Paraceratherium были большие слоновьи уши, которые он использовал для терморегуляции . Уши слонов увеличивают площадь поверхности тела и наполнены кровеносными сосудами, что облегчает рассеивание избыточного тепла. По мнению Протеро, это было верно и для Paraceratherium , о чем свидетельствуют крепкие кости вокруг ушных отверстий. [4] Палеонтологи Пьер-Оливье Антуан и Даррен Нэйш выразили скептицизм по отношению к этой идее. [39] [40]
Из-за фрагментарности известных окаменелостей Paraceratherium скелет животного с момента его открытия реконструировался несколькими различными способами. [41] В 1923 году Мэтью руководил художником, который нарисовал реконструкцию скелета на основе еще менее полных экземпляров P. transouralicum, известных к тому времени, используя в качестве ориентира пропорции современного носорога. [42] Результат оказался слишком приземистым и компактным, и позже в том же году Осборн нарисовал более тонкую версию. Некоторые более поздние реставрации сделали животное слишком стройным, не принимая во внимание основной скелет. [4] В 1959 году Громова опубликовала более полную реконструкцию скелета, основанную на скелете P. transouralicum из аральской свиты, но в ней также отсутствовало несколько шейных позвонков. [33]
Череп
[ редактировать ]Самые большие черепа Paraceratherium имеют длину около 1,3 метра (4,3 фута), от 33 до 38 сантиметров (от 13 до 15 дюймов) в задней части черепа и ширину 61 сантиметр (24 дюйма) по скуловым дугам . У парацератерия был длинный гладкий лоб без шероховатой части, служащей местом крепления рогов других носорогов. Кости над носовой областью длинные, носовой разрез заходит далеко в череп. Это указывает на то, что Paraceratherium имел цепкую верхнюю губу, похожую на таковую у черного носорога и индийского носорога , или короткий хобот (хобот), как у тапиров . [4] Отличительной особенностью было то, что носовой разрез был отведен к премолярам P2-P3. [17]
Задняя часть черепа была низкой и узкой, без больших лямбдоидных гребней наверху и вдоль сагиттального гребня , которые обычно встречаются у рогатых и клыкастых животных, которым нужны сильные мышцы, чтобы толкаться и сражаться. У него также была глубокая ямка для прикрепления выйных связок, которые автоматически удерживают череп. Затылочный мыщелок был очень широким, и у Paraceratherium, по-видимому, были большие и сильные мышцы шеи, что позволяло ему сильно наклонять голову вниз при поиске пищи на ветках. [4] Верхний профиль черепа дугообразный, что является отличительной особенностью рода. [17] У одного черепа P. transouralicum лоб куполообразный, тогда как у других лоб плоский, возможно, из-за полового диморфизма. [22] мозга Эндокаст показывает P. transouralicum , что его длина составляет всего 8 процентов от длины черепа, тогда как мозг индийского носорога составляет 17,7 процента от длины его черепа. [20]
Виды Paraceratherium в основном различимы по характеристикам черепа. P. bugtiense имел такие особенности, как относительно тонкие верхние и предчелюстные кости, неглубокие крыши черепа , сосцевидно-парокципитальные отростки, которые были относительно тонкими и располагались назад на черепе, ламбдоидный гребень, который меньше простирался назад, и затылочный мыщелок с горизонтальной ориентацией. который он разделил с Dzungariotherium . P. transouralicum имел крепкие верхние и предчелюстные кости, перевернутые скулы, куполообразные лобные кости , толстые сосцевидно-затылочные отростки, выступающий назад ламбдоидный гребень и затылочные мыщелки с вертикальной ориентацией. [3] P. huangheense отличался от P. bugtiense только анатомией задней части челюсти, а также более крупными размерами. [16] P. linxiaense отличался от других видов тем, что носовая вырезка была более глубокой, с расположением дна выше середины коренного зуба М2, пропорционально более высоким затылочным мыщелком по сравнению с высотой затылочной поверхности, короткими мордочными костями и диастемой перед щечными зубами. , высокая скуловая дуга с выступающим задним концом и меньший верхний резец I1. [17]
В отличие от зубов большинства примитивных носорогов, передние зубы Paraceratherium были уменьшены до одной пары резцов на каждой челюсти, которые были большими и коническими и были описаны как бивни. Верхние резцы были направлены вниз; нижние были короче и направлены вперед. Среди известных носорогов такое расположение уникально для Paraceratherium и родственного ему Urtinotherium . Резцы могли быть крупнее у самцов. Клыки , которые в противном случае находились за резцами, были утеряны. Резцы были отделены от ряда щечных зубов большой диастемой (промежутком). [4] Эта особенность встречается у млекопитающих, у которых резцы и щечные зубы имеют разную специализацию. [27] Верхние моляры, за исключением третьего верхнего моляра, который имел V-образную форму, имели пи-образную форму (π) и уменьшенный метастиль . Премоляры лишь частично сформировали рисунок «пи». Каждый коренной зуб был размером с человеческий кулак; среди млекопитающих их превосходили по размеру только хоботные, хотя они были небольшими по сравнению с размером черепа. Нижние щечные зубы имели Г-образную форму, что характерно для носорогов. [4]
Посткраниальный скелет
[ редактировать ]Полный набор позвонков и ребер Paraceratherium пока не обнаружен, а хвост совершенно неизвестен. Атлант и осевые позвонки шеи были шире, чем у большинства современных носорогов, и в них было место для сильных связок и мышц, необходимых для удержания большой головы. Остальные позвонки также были очень широкими и имели большие зигапофизы , в которых было много места для мышц, сухожилий, связок и нервов, поддерживающих голову, шею и позвоночник. Нервные отростки были длинными и образовывали длинный «горб» вдоль спины, к которому прикреплялись мышцы шеи и выйные связки, удерживающие череп. Ребра были похожи на ребра современных носорогов, но грудная клетка выглядела меньше по сравнению с длинными ногами и крупным телом, поскольку современные носороги сравнительно короткие конечности. Последний позвонок нижней части спины сросся с крестцом — особенность, обнаруженная у продвинутых носорогов. [4] Как и динозавры-зауроподы , Paraceratherium имели плевроцелеподобные отверстия (полые части костей) в предкрестцовых позвонках, что, вероятно, способствовало облегчению скелета. [43]
Конечности были большими и крепкими, чтобы выдерживать большой вес животного, и в некотором смысле были похожи и сходились с конечностями слонов и динозавров-зауроподов с их аналогичным гравипортальным (тяжелым и медленным) телосложением. В отличие от таких животных, которые имеют тенденцию удлинять кости верхних конечностей, одновременно укорачивая, сращивая и сжимая кости нижних конечностей, кистей и стоп, у Paraceratherium были короткие кости верхних конечностей и длинные кости кистей и стоп — за исключением дискообразных фаланг — аналогично бегущим носорогам, от которых они произошли. Некоторые кости стопы имели длину почти 50 сантиметров (20 дюймов). Бедренные кости обычно имели длину 1,5 м (4,9 фута), и этот размер превосходил кости лишь некоторых слонов и динозавров. Бедренные кости были столбообразными, намного толще и крепче, чем у других носорогов, а три вертела по бокам были значительно уменьшены, поскольку эта прочность уменьшала их важность. Конечности удерживались в колоннообразном положении, а не согнутыми, как у более мелких животных, что уменьшало потребность в крупных мышцах конечностей. [4] Передние конечности имели по три пальца. [44]
Палеобиология
[ редактировать ]В 1988 году зоолог Роберт М. Александер предположил, что перегрев мог быть серьезной проблемой для Paraceratherium из-за его размера. [45] По мнению Протеро, лучшими живыми аналогами Paraceratherium могут быть крупные млекопитающие, такие как слоны, носороги и гиппопотамы. Чтобы улучшить терморегуляцию, эти животные охлаждаются в течение дня, отдыхая в тени или валяясь в воде и грязи. Они также добывают пищу и передвигаются в основном ночью. Из-за своих больших размеров парацератерии не смогли бы быстро бегать и передвигаться, но могли бы преодолевать большие расстояния, что было бы необходимо в условиях нехватки пищи. Следовательно, они могли иметь большие ареалы обитания и мигрировать. [4] Протеро предполагает, что таким большим животным, как индрикотерии, потребуются очень большие ареалы обитания или территории площадью не менее 1000 квадратных километров (250 000 акров) и что из-за нехватки ресурсов в Азии было бы мало места для многих популяций или множества почти идентичных видов и родов. Этот принцип называется конкурентным исключением ; его используют для объяснения того, как черный носорог (обозреватель) и белый носорог (пастбище) занимают разные ниши в одних и тех же районах Африки. [3]
Большинство наземных хищников в их среде обитания были не крупнее современного волка и не представляли угрозы для Paraceratherium . [30] Взрослые особи были бы слишком большими для нападения наземных хищников, но молодые особи были бы уязвимы. Следы укусов на костях из пластов Бугти указывают на то, что даже на взрослых особей могли охотиться крокодилы Astorgosurus длиной от 10 до 11 метров (от 33 до 36 футов) . у слонов, период беременности Paraceratherium Как и мог быть длительным, а отдельные особи могли иметь долгую продолжительность жизни. [4] Парацератерии , возможно, жили небольшими стадами, состоявшими, возможно, из самок и их телят, которых они защищали от хищников. [30] Было высказано предположение, что 20 тонн (44 000 фунтов) могут быть максимально возможным весом для наземных млекопитающих, и Paraceratherium был близок к этому пределу. [46] Причины, по которым млекопитающие не могут достичь гораздо большего размера, чем динозавры-зауроподы, неизвестны. Причина может быть экологической, а не биомеханической и, возможно, связана со стратегиями воспроизводства. [34] Движение, звук и другое поведение, наблюдаемое в документальных компьютерных фильмах, таких как «Прогулки со зверями» , полностью предположительны. [4]
Диета и кормление
[ редактировать ]
Простые зубы с низкой коронкой указывают на то, что Paraceratherium был браузером, питавшимся относительно мягкими листьями и кустарниками. Позже носороги стали травоядными и имели зубы с высокой коронкой, поскольку в их рационе содержался песок, который быстро изнашивал зубы. Исследования мезоизноса на зубах Paraceratherium подтверждают, что эти существа питались мягкими листьями; исследования микроодежды еще не проводились. [4] Изотопный анализ показывает, что Paraceratherium питается в основном растениями C3 , которые в основном состоят из листьев. [47] [48] Подобно своим непарнокопытным родственникам, лошадям, тапирам и другим носорогам, Paraceratherium мог ферментировать заднюю кишку ; он будет получать относительно мало питательных веществ из своей пищи, и ему придется потреблять большие объемы, чтобы выжить. Как и у других крупных травоядных, у Paraceratherium был большой пищеварительный тракт. [4]
Грейнджер и Грегори утверждали, что большие резцы использовались для защиты или для рыхления кустов путем перемещения шеи вниз, действуя тем самым как кирки и рычаги. [20] Тапиры используют хоботок, чтобы обхватывать ветки, сдирая кору передними зубами; эта способность была бы полезна для Парацератерия . Некоторые российские авторы предположили, что бивни, вероятно, использовались для ломания веток, снятия коры и сгибания высоких ветвей, и что, поскольку виды из раннего олигоцена имели более крупные бивни, чем более поздние, они, вероятно, питались больше корой, чем листьями. Поскольку теперь известно, что рассматриваемые виды были одновременными, а различия в бивнях теперь считаются половым диморфизмом, последняя идея сегодня не принимается. [4] Стада Paraceratherium , возможно, мигрировали, постоянно добывая пищу с высоких деревьев, до которых более мелкие млекопитающие не могли добраться. [30] Осборн предположил, что его способ добывания пищи был похож на способ добывания пищи у жирных жирафов и окапи , а не у современных носорогов, чьи головы тянутся близко к земле. [42]
Распространение и среда обитания
[ редактировать ]
Останки, относящиеся к Paraceratherium, были обнаружены в формациях раннего и позднего олигоцена (34–23 миллиона лет назад) по всей Евразии, в современном Китае, Монголии, Индии, Пакистане, Казахстане, Грузии, Турции, Румынии, Болгарии и на Балканах . [5] Их распространение можно соотнести с палеогеографическим развитием Альпийско-Гималайского горного пояса. Диапазон находок Paraceratherium предполагает, что они населяли сплошную территорию с одинаковой средой обитания по всей ней, но это противоречит палеогеографическим картам, которые показывают, что на этой территории были различные морские барьеры, поэтому, несмотря на это, этому роду удалось широко распространиться. [49] В фауну, сосуществовавшую с Paraceratherium, входили другие носороги, парнокопытные , грызуны, амфициониды , куньи , гиенодонты , нимравиды и кошачьи . [30]
Среда обитания Paraceratherium , по-видимому, различалась в зависимости от типа геологических образований, в которых он был обнаружен. [30] Формация Хсанда-Гол в Монголии представляет собой засушливый пустынный бассейн, и считается, что в этой среде было мало высоких деревьев и ограниченный кустарниковый покров, поскольку фауна состояла в основном из животных, которые питались с верхушек деревьев или близко к земле. [50] Исследование ископаемой пыльцы показало, что большая часть Китая представляет собой древесно-кустарниковую местность с такими растениями, как солончак , мормонский чай ( Эфедра ) и селитровый куст ( Нитрария ), приспособленные к засушливой среде. Деревья были редкими и концентрировались вблизи грунтовых вод. [51] В тех частях Китая, где обитали Paraceratherium , были высохшие озера и обильные песчаные дюны , а наиболее распространенными окаменелостями растений являются листья адаптированной к пустыне Палибинии . Деревья в Монголии и Китае включали березу, вяз, дуб и другие лиственные деревья, а в Сибири и Казахстане также были ореховые деревья. [30] Дера Бугти в Пакистане имел сухой лес от умеренного до субтропического. [47]

Дэн и его коллеги высказали предположения о палеобиогеографии Paraceratherium P. на основе своего филогенетического анализа в 2021 году. Они обнаружили, что bugtiense был единственным видом этого рода, представленным в олигоцене западного Пакистана, в то время как этот род был очень разнообразен на Монгольском плато , северо-западе Пакистана. Китай и Казахстан на север до Тибетского нагорья . Они предположили, что P. asiaticum распространился на запад, в Казахстан, в раннем олигоцене из ареала предков Монголии, где жил самый примитивный представитель рода, P. grangeri , и его потомки, возможно, продолжили путь в Южную Азию как P. bugtiense , расселившись. через Тибетский регион. P. lepidum существовал в Синьцзяне и Казахстане, а P. linxiaense - в Линься в позднем олигоцене, и вполне возможно, что эти родственные виды смогли P. bugtiense мигрировать обратно на север, в Среднюю Азию, в это время, когда эта территория стала тропической. (в раннем олигоцене было засушливо). Это означает, что Тибетский регион еще не был высокогорным плато, которое могло бы служить барьером, и поэтому крупные животные могли свободно передвигаться вдоль восточного побережья Тибета. Море Тетис и низменности в этом районе, некоторые из которых, возможно, в то время находились на высоте менее 2000 м (6600 футов) над уровнем моря. [17]
Вымирание
[ редактировать ]
Причины вымирания Paraceratherium и его родственников, просуществовавших около 11 миллионов лет, неизвестны, но маловероятно, что это была одна причина. Теории включают в себя то, что их большой размер был связан с ныне устаревшей концепцией неадаптивной эволюции , изменением климата , изменением растительности и низкой скоростью воспроизводства . Протеро и зоолог Павел В. Путшков сочли эти причины маловероятными, поскольку этим животным удавалось выжить, несмотря на эти проблемы, в течение миллионов лет в суровых условиях окружающей среды, и они были ненамного крупнее самых крупных хоботных, как вымерших, так и современных. , который столкнулся с аналогичными проблемами. [30] [52]
Вместо этого Путшков и Анджей Х. Кульчицкий предположили в 1995 и 2001 годах, что вторжение хоботных гомфотер из Африки в позднем олигоцене (между 28 и 23 миллионами лет назад), возможно, значительно изменило среду обитания, в которую они вошли, как это делают сегодня африканские слоны . Это привело бы к ограничению продовольствия для Paraceratherium , и по мере того, как их численность сокращалась, они стали бы более уязвимыми для других угроз. [52] Протеро отметил, что гомфотерии, как известно, обычно не сосуществовали с парацератериями, и нет известных случаев одновременного существования парацератеров и крупных дейнотериев , которые были бы их наиболее вероятными конкурентами. Предупреждая, что истинная причина их вымирания никогда не будет известна наверняка, Протеро счел более чем совпадением то, что парацератеры исчезли как раз тогда, когда крупные хищники и другие крупные травоядные животные проникли в Азию в раннем миоцене (между 23 и 16 миллионами лет назад). ). [30]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и Протеро, 2013. стр. 17–34
- ^ Jump up to: а б с Маниас, К. (2014). «Создание белухитерия и индрикотерия : имперские и международные сети в палеонтологии начала двадцатого века» (PDF) . Журнал истории биологии (Представлена рукопись). 48 (2): 237–78. дои : 10.1007/s10739-014-9395-y . ПМИД 25537636 . S2CID 207150574 .
- ^ Jump up to: а б с д и ж г час Протеро, 2013. стр. 67–86
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р Протеро, 2013. стр. 87–106
- ^ Jump up to: а б с Протеро, 2013. стр. 35–52
- ^ Пилигрим, GE (1910). «Уведомления о новых родах и видах млекопитающих третичного периода Индии». Отчеты Геологической службы Индии . 40 (1): 63–71.
- ^ Jump up to: а б Купер, CF (1924). «О черепе и зубных рядах Paraceratherium bugtiense : род аберрантных носорогов из нижнемиоценовых отложений Дера Бугти» . Философские труды Королевского общества B: Биологические науки . 212 (391–401): 369–394. Бибкод : 1924RSPTB.212..369F . дои : 10.1098/rstb.1924.0009 .
- ^ Jump up to: а б Форстер-Купер, К. (1911). «LXXVIII. — Paraceratherium bugtiense , новый род Rhinocerotidae с холмов Бугти в Белуджистане. — Предварительное уведомление» . Анналы и журнал естественной истории . Серия 8. 8 (48): 711–716. дои : 10.1080/00222931108693085 . Архивировано из оригинала 20 ноября 2015 года . Проверено 28 августа 2015 г.
- ^ Форстер-Купер, К. (1913). «XLIV. — Thaumastotherium osborni , новый род непарнокопытных из верхнеолигоценовых отложений холмов Бугти в Белуджистане. — Предварительное уведомление» . Анналы и журнал естественной истории . Серия 8. 12 (70): 376–381. дои : 10.1080/00222931308693412 . Архивировано из оригинала 7 октября 2015 года . Проверено 28 августа 2015 г.
- ^ Форстер-Купер, К. (1913). «Исправление родового названия» . Анналы и журнал естественной истории . Ряд 8. 12 (71): 504. doi : 10.1080/00222931308693431 . Архивировано из оригинала 12 августа 2018 года . Проверено 13 января 2018 г.
- ^ Форстер-Купер, К. (1923). « Baluchitherium osborni (? син. Indricotherium turgaicum , Borrissyak)» . Философские труды Лондонского королевского общества . 212 (391–401): 35–66. дои : 10.1098/rstb.1924.0002 . JSTOR 92060 .
- ^ Павлова, М. (1922). « Indricotherium transouralicum n. sp. из Тургайского района». Вестник Московского общества естествоиспытателей, геологический отдел (на французском языке). 31 : 95–116.
- ^ Бориссяк, А.А. (1924). «О подсемействе Indricotheriinae Борисс. = Baluchitheriinae Osb». Центральный журнал минералогии, геологии и палеонтологии (на немецком языке). 18 :571-575.
- ^ Протеро, 2013. стр. 1–16
- ^ Осборн, HF (1923). «Вымерший гигантский носорог Baluchitherium Западной и Центральной Азии» . Естественная история . 3. 23 : 208–228. Архивировано из оригинала 9 марта 2016 года . Проверено 8 сентября 2014 г.
- ^ Jump up to: а б Юн-Сян, Л.; Юн-Сян, З.; Джи, Л.; Чжи-Чао, Л.; Кун, X. (2017). «Новые окаменелости парацератеров (Perissodactyla, Mammalia) из раннего олигоцена бассейна Ланьчжоу, провинция Ганьсу, Китай» . Позвоночные Палазиатские . 55 (4): 367–381. doi : 10.19615/j.cnki.1000-3118.170922 . Архивировано из оригинала 12 октября 2017 года . Проверено 11 октября 2017 г.
- ^ Jump up to: а б с д и ж г Дэн, Т.; Лу, Х.; Ван, С.; Флинн, LJ; Сан, Д.; Он, В.; Чен, С. (2021). «Гигантский носорог олигоцена дает представление об Paraceratherium эволюции » . Коммуникационная биология . 4 (1): 639. дои : 10.1038/s42003-021-02170-6 . ПМЦ 8211792 . ПМИД 34140631 .
- ^ Форстер-Купер, К. (1922). «LXXIV.— Metamynodon bugtiensis , sp. n., из отложений Дера Бугти в Белуджистане. — предварительное уведомление» . Анналы и журнал естественной истории . 9 (53): 617–620. дои : 10.1080/00222932208632717 .
- ^ Тиссье, Дж.; Беккер, Д.; Кодря, В.; Костёр, Л.; Фаркаш, К.; Соломон, А.; Венцель, М.; Маридет, О.; Смит, Т. (2018). «Новые данные об Amynodontidae (Mammalia, Perissodactyla) из Восточной Европы: филогенетические и палеобиогеографические последствия перехода эоцена в олигоцен» . ПЛОС ОДИН . 13 (4): e0193774. Бибкод : 2018PLoSO..1393774T . дои : 10.1371/journal.pone.0193774 . ПМК 5905962 . ПМИД 29668673 .
- ^ Jump up to: а б с Грейнджер, В.; Грегори, WK (1936). «Дальнейшие заметки о гигантском вымершем носорогове Baluchitherium из олигоцена Монголии». Бюллетень Американского музея естественной истории . 72 : 1–73. HDL : 2246/363 .
- ^ Форстер-Купер, К. (1934). «Вымершие носороги Белуджистана» . Философские труды Королевского общества B: Биологические науки . 223 (494–508): 569–616. Бибкод : 1934RSPTB.223..569F . дои : 10.1098/rstb.1934.0013 .
- ^ Jump up to: а б с д и ж г Лукас, СГ; Собус, Дж. К. (1989), «Систематика индрикотериев» , в Протеро, ДР; Шох, Р.М. (ред.), Эволюция непарнокопытных , Нью-Йорк, Нью-Йорк и Оксфорд, Англия: Oxford University Press , стр. 358–378, ISBN. 978-0-19-506039-3 , OCLC 19268080
- ^ Чжан-Сян, Ц. (1973). «Новый род гигантских носорогов из олигоцена Джунгарии, Синкан» (PDF) . Позвоночные Палазиатские . 11 (на китайском и английском языках). 2 : 182–191. Архивировано (PDF) из оригинала 22 октября 2014 г. Проверено 22 октября 2014 г.
- ^ Антуан, ПО; Караденизли, Л.; Сарач, GE; Сен, С. (2008). «Гигантский носорог (Mammalia, Perissodactyla) из позднего олигоцена северо-центральной Анатолии (Турция)» . Зоологический журнал Линнеевского общества . 152 (3): 581–592. дои : 10.1111/j.1096-3642.2007.00366.x .
- ^ Йе, Й.; Дж., Мэн; Ю, В.В. (2003). «Обнаружение парацератерия в северной части Джунгарской котловины Синьцзяна» (PDF) . Vertebrata PalAsiatica (на китайском и английском языках). 41 : 220–229. Архивировано (PDF) из оригинала 3 марта 2016 года . Проверено 22 октября 2014 г.
- ^ Куи, З.-Х.; Ван, Б.-Ю.; Дэн, Т. (2004). «Indricotheres (Perissodactyla, Mammalia) из олигоцена в бассейне Линься, Ганьсу, Китай» (PDF) . Vertebrata PalAsiatica (на китайском и английском языках). 42 (3): 177–192. Архивировано из оригинала (PDF) 22 октября 2014 года . Проверено 22 октября 2014 г.
- ^ Jump up to: а б с д Протеро, 2013. стр. 53–66
- ^ Радинский, Л.Б. (1966). «Семейства Rhinocerotoidea (Mammalia, Perissodactyla)». Журнал маммологии . 47 (4): 631–639. дои : 10.2307/1377893 . JSTOR 1377893 .
- ^ Холбрук, Л. (1999). «Филогения и классификация тапироморфных непарнокопытных (Mammalia)» . Кладистика . 15 (3): 331–350. дои : 10.1006/клад.1999.0107 . ПМИД 34902952 .
- ^ Jump up to: а б с д и ж г час я Протеро, 2013. стр. 107–121
- ^ Лукас, СГ; Шох, РМ; Мэннинг, Э. (1981). «Систематика Forstercooperia , гиракодонтида среднего и позднего эоцена (Perissodactyla: Rhinocerotoidea) из Азии и западной части Северной Америки». Журнал палеонтологии . 55 (4): 826–841. JSTOR 1304430 .
- ^ Ван, Х.; Бай, Б.; Мэн, Дж.; Ван, Ю. (2016). «Самый ранний из известных носорогов проливает новый свет на происхождение гигантских носорогов и филогению ранних носорогов» . Научные отчеты . 6 (1): 39607. Бибкод : 2016NatSR...639607W . дои : 10.1038/srep39607 . ПМК 5175171 . ПМИД 28000789 .
- ^ Jump up to: а б Gromova, V. L. (1959). "Gigantskie nosorogi". Trudy Paleontology Institut Akademii Nauk SSSR (in Russian). 71 : 154–156.
- ^ Jump up to: а б с Фортелиус, М.; Каппельман, Дж. (1993). «Самое большое наземное млекопитающее, которое когда-либо можно было себе представить». Зоологический журнал Линнеевского общества . 108 : 85–101. дои : 10.1111/j.1096-3642.1993.tb02560.x .
- ^ Ларраменди, А. (2016). «Высота плеч, масса тела и форма хоботков» (PDF) . Acta Palaeontologica Polonica . 61 . дои : 10.4202/app.00136.2014 . S2CID 2092950 . Архивировано (PDF) из оригинала 24 августа 2016 г.
- ^ Пол, GS (1997). «Модели динозавров: хорошие, плохие и их использование для оценки массы динозавров» (PDF) . Международные слушания Динофеста : 129–142. Архивировано (PDF) из оригинала 4 марта 2016 года . Проверено 3 января 2015 г.
- ^ Тейлор, член парламента; Ведель, MJ (2013). «Почему у зауроподов были длинные шеи и почему у жирафов шеи короткие» . ПерДж . 1 : е36. дои : 10.7717/peerj.36 . ПМЦ 3628838 . ПМИД 23638372 .
- ^ Цубамото, Т. (2012). «Оценка массы тела по таранной кости у млекопитающих» . Acta Palaeontologica Polonica : 259–265. дои : 10.4202/app.2011.0067 . S2CID 54686160 .
- ^ Антуан, ПО (2014). «В те дни на земле жили великаны (Рецензия на книгу)» . Палеовертебрата . 38 : 1–3. дои : 10.18563/pv.38.1.e4 . S2CID 203264606 . Архивировано из оригинала 12 ноября 2014 года . Проверено 20 октября 2014 г.
- ^ Нэйш, Д. (29 июня 2013 г.). Ван Гроува «Книжная полка зоопарка Тет: «Неоперенная птица» Бодио , «Вечность орлов» » Уиттона , «Птерозавры , «Морские чудовища» Ван Дузера на картах Средневековья и Возрождения !» . blogs.scientificamerican.com/tetrapod-zoology . Архивировано из оригинала 6 октября 2014 года . Проверено 5 октября 2014 г.
- ^ Грейнджер, В.; Грегори, WK (1935). «Пересмотренная реставрация скелета Baluchitherium , гигантского ископаемого носорога Центральной Азии». Новитаты Американского музея (787): 1–3. hdl : 2246/2123 .
- ^ Jump up to: а б Осборн, HF (1923). « Baluchitherium grangeri , гигантский безрогий носорог из Монголии». Новитаты Американского музея (78): 1–15. HDL : 2246/3262 .
- ^ Сандер, премьер-министр; Кристиан, А.; Клаусс, М.; Фехнер, Р.; Ну и дела, Коннектикут; Грибелер, Э.М.; Гунга, ХК; Хаммель, младший; Мэллисон, Х.; Перри, Сан-Франциско; Пройшофт, Х.; Раухут, OWM; Ремес, К.; Тюткен, Т.; Уингз, О.; Витцель, У. (2011). «Биология динозавров-зауроподов: Эволюция гигантизма» . Биологические обзоры . 86 (1): 117–155. дои : 10.1111/j.1469-185X.2010.00137.x . ПМК 3045712 . ПМИД 21251189 .
- ^ Антуан, ПО; Ибрагим Шах, SM; Чима, АйЮ; Крючком, JY; Франчески, Д.Д.; Мариво, Л.; Метаис, Г.Г.; Добро пожаловать, Дж.Л. (2004). «Новые останки белухитера Paraceratherium bugtiense из позднего/позднего олигоцена холмов Бугти, Белуджистан, Пакистан». Журнал азиатских наук о Земле . 24 (1): 71–77. Бибкод : 2004JAESc..24...71A . дои : 10.1016/j.jaes.2003.09.005 .
- ^ Александр, РМ (1998). «Гиганты на все времена: самые большие животные и их проблемы» . Палеонтология . 41 (6): 1231–1245. Архивировано из оригинала 27 сентября 2016 года . Проверено 1 марта 2016 г.
- ^ Клаусс, М.; Фрей, Р.; Кифер, Б.; Лехнер-Долл, М.; Лелейн, В.; Польстер, К.; Рёсснер, GE; Штрайх, WJ (2003). «Максимально достижимый размер тела травоядных млекопитающих: морфофизиологические ограничения передней кишки и адаптации ферментеров задней кишки» (PDF) . Экология (Представлена рукопись). 136 (1): 14–27. Бибкод : 2003Oecol.136...14C . дои : 10.1007/s00442-003-1254-z . ПМИД 12712314 . S2CID 206989975 . Архивировано (PDF) из оригинала 8 июня 2019 года . Проверено 6 августа 2018 г.
- ^ Jump up to: а б Мартин, К.; Бенталеб, И.; Антуан, П. -О. (2011). «Стабильные изотопы зубов пакистанских млекопитающих демонстрируют палеоклиматические и палеоэкологические изменения с раннего олигоцена». Палеогеография, Палеоклиматология, Палеоэкология . 311 (1–2): 19–29. Бибкод : 2011PPP...311...19M . дои : 10.1016/j.palaeo.2011.07.010 .
- ^ Ван, Ю.; Дэн, Т. (2005). «25 моих изотопных записей палеодиеты и изменений окружающей среды по ископаемым млекопитающим и палеопочвам северо-восточной окраины Тибетского нагорья». Письма о Земле и планетологии . 236 (1–2): 322–338. Бибкод : 2005E&PSL.236..322W . дои : 10.1016/j.epsl.2005.05.006 .
- ^ Сен, С.; Антуан, ПО; Вароль, Б.; Айилдиз, Т.; Созери, К. (2011). «Гигантский носорог Paraceratherium и другие позвоночные из олигоценовых и среднемиоценовых отложений бассейна Кагызман-Тузлука, Восточная Турция». Naturwissenschaften . 98 (5): 407–423. Бибкод : 2011NW.....98..407S . дои : 10.1007/s00114-011-0786-z . ПМИД 21465174 . S2CID 19968371 .
- ^ Мэн, Дж.; Маккенна, MC (1998). «Фаунистические обороты палеогеновых млекопитающих Монгольского нагорья». Природа . 394 (6691): 364–367. Бибкод : 1998Natur.394..364M . дои : 10.1038/28603 . S2CID 204998953 .
- ^ Леопольд, Е.Б.; Лю, Г.; Клэй-Пул, С. (1992), «Растительность с низкой биомассой в олигоцене?», в Протеро, Д.Р .; Берггрен, Вашингтон (ред.), Эоцен-олигоценовая климатическая и биотическая эволюция , Принстон: Princeton University Press, стр. 399–420, ISBN. 978-0-691-02542-1
- ^ Jump up to: а б Путшков, П.В. (2001). « Хоботок-агент» некоторых третичных вымираний мегафауны». Terra Degli Elefanti Congresso Internazionale: Мир слонов : 133–136.
Библиография
[ редактировать ]- Протеро, Д. (2013). Гиганты-носороги: палеобиология индрикотериев . Индиана: Издательство Университета Индианы. ISBN 978-0-253-00819-0 .
- Парацератерииды
- Олигоценовые носороги
- Аквитанское вымирание родов
- Доисторические плацентарные роды
- Вымершие животные Пакистана
- Первые появления рода Rupelian
- Ископаемые таксоны описаны в 1911 году.
- Таксоны, названные Клайвом Форстером-Купером
- Олигоценовые млекопитающие Азии
- Олигоценовые млекопитающие Европы