Дерево ласточка
| Дерево ласточка | |
|---|---|

| |
| Взрослый в Ямайском заливе, заповедник дикой природы в Квинсе, Нью -Йорк | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Сорт: | Птица |
| Заказ: | Прохожий |
| Семья: | Hirundinidae |
| Род: | Тахицинета |
| Разновидность: | Т. Биколор
|
| Биномиальное название | |
| Tachycineta Bicolor ( Старый модный , 1808)
| |
| |
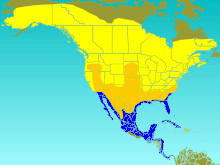
| Диапазон T. bicolor Разведение летнего посетителя Миграционный посетитель Зимний посетитель
| |
| Синонимы | |
|
Hirundo Bicolor Vieillot, 1808 | |

( Ласточка дерева Tachycineta Bicolor ) - это мигрирующая птица семьи Hirundinidae . Найденная в Америке , ласточка дерева была впервые описана в 1807 году французским орнитологом Луи Вейилотом в роли Хирундо Биколор . С тех пор он был перенесен на свой нынешний род Tachycineta , в рамках которого обсуждается его филогенетическое размещение. Ласточка дерева имеет глянцевые голубо-зеленые верхние части , за исключением черных крыльев и хвоста и белых нижних частей . Счет черный, глаза темно -коричневые, а ноги и ноги бледно -коричневые. Женщина, как правило, тусая, чем мужчина, а у самой первой год у самки в основном коричневые верхние части, с некоторыми голубыми перьями. У несовершеннолетних есть коричневые верхние части и серо-коричневые грудь. Ласточка дерева порождает в США и Канаде. Он зима вдоль южного побережья США на юг, вдоль побережья Мексиканского залива, до Панамы и северо -западного побережья Южной Америки и в Вест -Индии .
Дерево глотает гнездятся либо в изолированных парах, либо в свободных группах, как в естественных, так и в искусственных полостях. Разведение может начаться уже в начале мая, хотя эта дата происходит ранее из -за изменения климата , и это может закончиться уже в июле. Эта птица, как правило, является социально моногамной (хотя около 8% мужчин являются полигинскими ), с высоким уровнем отцовства вверху . Это может принести пользу мужчине, но, поскольку женщина контролирует совокупление, отсутствие разрешения на то, как это поведение приносит пользу женщинам, делает высокий уровень загадки от отцовства. Самка инкубирует сцепление от двух до восьми (но обычно от четырех до семи) чистых белых яиц в течение примерно 14-15 дней. Циклы немного асинхронно вылупляются, позволяя самке расставлять приоритеты, какие цыплят кормит во времена нехватки пищи. Обычно они усажены примерно от 18 до 22 дней после вылупления. Ласточка дерева иногда считается образцовым организмом из -за большого количества исследований, проведенных на нем.
Аэрофотоялочный насекомые , дерево глотает корму как в одиночестве, так и в группах, есть в основном насекомых , в дополнение к моллюскам , паукам и фруктам. Приходы, как взрослые, в основном едят насекомых, питаемые им обоими полов. Эта ласточка уязвима для паразитов, но, когда на птенцах это мало ущерб. Эффект заболевания может стать сильнее по мере того, как ласточка дерева становится старше, так как некоторые части иммунной системы снижаются с возрастом. приобретенный иммунитет, опосредованный Т -клетками Например, , уменьшается с возрастом, тогда как как врожденный, так и приобретенный гуморальный иммунитет не делают. Из -за своего большого диапазона и стабильной популяции ласточка дерева считается наименее обеспокоенной Международным союзом сохранения природы . В США он защищен Законом о договоре о миграционных птицах 1918 года , а в Канаде Законом о конвенции о перелетных птицах . Эта ласточка негативно влияет на человеческую деятельность, такую как очистка лесов; Окинимые озера могут заставлять ласточку для размножения, чтобы пройти на большие расстояния, чтобы найти богатые кальцием пищевые продукты, которые питаются его цыплятами.
Таксономия и этимология
[ редактировать ]Ласточка дерева была описана как Биколор Хирундо Луи Пьером Вийлотом в его естественной истории птиц Северной Америки , опубликованной в 1807 году. [ 2 ] Затем он был помещен в его нынешний род Tachycineta , когда Джин Кабанис основал его в 1850 году. [ 3 ] В 1878 году Эллиотт Коус предположил, что ласточка дерева, по крайней мере, будет помещена в свой собственный подрод , иридопрокни , на основе его оперения вместе с белой ласточкой , ласточкой из чили , белой ласточкой и мангроув. глотать . [ 4 ] К 1882 году он обновил это до полного рода. [ 5 ] Некоторые авторы продолжали использовать эту классификацию с добавлением ласточки Tumbes ; Однако генетические данные подтверждают существование одного рода, Tachycineta . [ 6 ] Ласточка дерева также называется белоснежной ласточкой для ее белых нижних частей. [ 7 ]
Общепринятое название рода-от древнегреческого тахикинетоса , «движущийся быстро», а конкретная двукратная биколора является латинской и означает «двухцветный». [ 8 ] Другое название рода, иридопрокни , происходит от греческой радужной оболочки , что означает Радугу, и Прокне , фигура, которая предположительно превратилась в ласточку. [ 5 ]
То, как именно ласточка дерева связана с другими членами Tachycineta , неразрешено. В исследованиях, основанных на митохондриальной ДНК , она была размещена базально (то есть это был первый ответвление в дереве вида) в рамках североамериканской карибской клады, состоящей из фиолетолето-зеленой ласточки , золотой ласточки и бахамы . [ 9 ] [ 10 ] Хотя митохондриальная ДНК выступает в качестве лучшего показателя эволюционных изменений, поскольку она развивается быстро, анализы, основанные на ней, могут пострадать, потому что она только наследуется от матери, что делает ее хуже, чем ядерная ДНК из нескольких локусов , представляющих филогения всей группы. [ 11 ] Исследование, основанное на такой ядерной ДНК, поместило ласточку дерева в наиболее базальное положение в тахицинете в целом (как родственная группа для остальной части рода). [ 12 ]
Описание
[ редактировать ]
Ласточка дерева имеет длину от 12 до 14 см (4,7 и 5,5 дюйма) и вес приблизительно от 17 до 25,5 г (от 0,60 до 0,90 унции). Размах крыльев колеблется от 30 до 35 см (от 12 до 14 дюймов). [ 13 ] У мужчины в основном глянцевые голубо-зеленые верхние части , крылья и хвост являются черноватыми. Недоверные [ 14 ] и пятно щеки белые, [ 7 ] Хотя нижние чехлы серо-коричневые. [ 14 ] Счет черный, глаза темно -коричневые, а ноги и ноги бледно -коричневые. [ 7 ] Женщина тупая по цвету, чем у мужчины, а иногда у коричневого лба. [ 14 ] Второй год [ Примечание 1 ] У женщины также есть коричневые верхние части, с переменным количеством синих перьев; Некоторые женщины третьего курса также сохраняют часть этого оперения субадульта. [ 15 ] Согласно исследованию 1987 года, это, вероятно, позволяет младшей женщине исследовать места гнезд, поскольку резидентский мужчина обычно менее агрессивен для субадультской женщины. [ 16 ] Исследование 2013 года показало, что резидентская женщина была менее агрессивной по отношению к женским моделям второго курса, когда они были представлены отдельно от более старых моделей. Почему самка в конечном итоге заменяет свое оперение Subadult, неизвестна; Это может позволить мужчинам оценивать качество женщин, так как пары сочетаются в эфире на основе яркости оперения. [ 17 ] Ласточку для ювенильных деревьев можно различить его коричневыми верхним частями и серо-коричневой грудью. [ 14 ]
Голос
[ редактировать ]

Песня Tree Wallow состоит из трех частей: чирп, скули и бульки. Эти разделы могут быть повторены или опущены, и все могут стоять в одиночестве. Первый, как вызов чирпов (иногда делится на контактный вызов и вызов запроса), сделан женщиной во время совокупления [ 18 ] и у обоих полов стимулировать птенцов просить [ 19 ] или (в некоторых популяциях), когда их приятель уходит или входит в полость гнезда. Наколот, как правило, состоящий из сдвига вниз по частоте, за которым следует сдвиг вверх, может быть дано в одиночку, как вызов тревоги, [ 18 ] иногда делается в ответ на определенные хищники. [ 20 ] Булька, как и когда она появляется в конце песни, обычно произносится дважды. Это, вероятно, вовлечено в парную связь. Звонок в болтовне используется для рекламы сайтов гнезд (причина, по которой он также известен как «рекламный звонок на сайте гнездо»), а также предоставляется вторжению конспецификов. Короткий высокий вызов подчинения иногда произносят после агрессивного столкновения с другой ласточкой дерева. Будучи физически ограниченным или больно, может быть дано вызов бедствия. Мужчина часто произносит агрессию тикания (или разрывы) во время совокупления, и оба пола используют его в конце моббинговых погружений. Вызов сигнализации дается в ответ на хищников и других злоумышленников, [ 18 ] и может служить для того, чтобы побудить более старые птенцы приседать и прекратить просить, когда находится хищник. [ 21 ]
Общение между родителями и потомками может быть нарушено из-за человеческого шума. Например, исследование 2014 года показало, что выводки, для которых играл белый шум, с меньшей вероятностью приседают или перестали попросить в ответ на звонки с сигнализацией. Родители не изменяли свои призывы, чтобы компенсировать, вероятно, увеличивая риск хищничества. [ 22 ] Шум также может нарушить, реагируют ли родители на попрошайничество, но это может быть сбалансировано с помощью громких звонков, которые приписывают приходы. Однако увеличение усилий по умолчанию может быть неэффективным или дорогостоящим для более громкого уровня шума. [ 23 ]
Распределение и среда обитания
[ редактировать ]Ласточка дерева порождает в Северной Америке. Его диапазон простирается до северной центральной Аляски и до линии деревьев в Канаде. Он найден на юге, как Теннесси в восточной части своего диапазона, Калифорнии и Нью -Мексико на Западе, и Канзас в центре. Иногда он порождает дальше на юг в США, [ 7 ] А бродяги иногда встречаются в Арктике , северной части Тихого океана , Гренландии и Европы. [ 14 ] Зимний хребет от Калифорнии и юго -западной Аризоны на западе и юго -восточной Вирджинии на востоке юга вдоль побережья Персидского залива до Вест -Индии , Панамы и северо -западного побережья Южной Америки. [ 7 ] Во время мигрирования эта ласточка часто использует остановку участки, тратит в среднем 57 дней в этих областях осенью. Чтобы добраться до своего зимнего диапазона, он часто использует один из трех пролетов: западный пролет, к западу от Скалистых гор ; Центральный пролет, между Скалистыми горами и Великими озерами , простирающимися на юг в восточную Мексику; и восточный пролет, с Востока Великих озер. [ 24 ] Когда ласточка возвращается в гнездо, она обычно не меняет места размножения. [ 25 ]
Среда обитания этой птицы находится в основном в открытых и лесистых районах, особенно вблизи воды. [ 7 ] Он разжигает каждую ночь в течение непредудингового сезона, предпочитая отдыхать в тростниковой или тростниковой кроватях над водой, но также встречается на земле, на деревьях и проводах. Участки, как правило, составляют от 100 до 150 километров (от 62 до 93 миль) друг от друга. [ 26 ]
Поведение
[ редактировать ]
Из -за большого количества исследований по ласточке деревьев и того, как оно охотно размножается в ящиках гнезд, биолог Джейсон Джонс рекомендовал его считать модельным организмом . [ 27 ] Хотя это агрессивно в течение сезона размножения, эта ласточка общительна вне ее, образуя стада, иногда насчитывая тысячи птиц. [ 7 ]
Размножение
[ редактировать ]
Дерево глотает гнездятся в структурах с ранее существовавшими отверстиями, как естественными, так и искусственными. Когда -то они были обнаружены только в лесных регионах, но здание гнездовых ящиков позволило этой птице расширяться в открытые места обитания. [ 7 ] Эта ласточка обычно гнездится в области, которую она разводила годом ранее; Только около 14% женщин и 4% мужчин рассеиваются, чтобы размножаться на новом участке в год. Большинство не уходят далеко, обычно размножаясь на участках менее 10 километров (6,2 мили) от их первоначальных оснований. На рассеивание влияет успех размножения; Из взрослых женщин, которые не опускают птенца, около 28% рассеиваются по сравнению с 5% успешных заводчиков. [ 25 ] Натальная рассеяние (когда птица не возвращается на место, в которой она родилась для размножения), распространена на ласточке деревьев и встречается чаще, чем рассеяние размножения. [ 28 ] Он гнездится как в свободных группах, так и в изолированных парах. При гнездовании в свободных группах гнезда обычно расположены на расстоянии не менее 10-15 метров (от 33 до 49 футов) друг от друга, [ 14 ] И те, которые находятся ближе на расстоянии, обычно находятся дальше с точки зрения даты укладки. [ 29 ] В природных полостях дерево гнездится около 27 метров (89 футов), кроме соседа. В среднем отверстие гнезда в этих ситуациях составляет 3,4 метра (11 футов) выше уровня земли, хотя около 45% из них составляют менее 2 метров (6,6 фута) над землей. Более высокие полости, вероятно, пользуются предпочтением, потому что они снижают хищничество, в то время как нижние гнездовые отверстия могут быть выбраны, чтобы избежать конкуренции. Ширина входа часто составляет от 4 до 5 сантиметров (1,6 и 2,0 дюйма), тогда как высота входа более изменчивы: исследование 1989 года обнаружило, что отверстия в диапазоне от 3,5 до 26 сантиметра (от 1,4 до 10,2 дюйма). Объем полости обычно ниже 1000 см 3 (61 с вн.). [ 30 ] Найдя подходящее место для гнезда, мужские мужские окуняются рядом с ним и часто звонит. Недостаток участков может вызвать драки между птицами, иногда приводит к смерти. Эта ласточка обычно защищает область вокруг гнезда с радиусом около 4,6 метра (15 футов), [ 7 ] а также дополнительные гнезда внутри этой территории , [ 14 ] блокируя вход в гнездо и преследуя злоумышленников. [ 7 ] Сама чашка гнезда сделана из травы, моха, сосновых игл и водных растений, собранных в основном самкой, [ 14 ] и выстлана перьями, собранными в основном мужчинами в боях. [ 31 ] Перья могут функционировать для изоляции гнезда, уменьшая время инкубации [ 32 ] и, вероятно, предотвращение гипотермии у цыплят. В дополнение к более быстрому росту для цыплят, [ 31 ] Яйца прохладно медленнее в гнездах с перьями, чем без. [ 33 ] Тем не менее, исследование, опубликованное в 2018 году, не обнаружило значительной корреляции между количеством перьев в гнездах, которые были искусственно нагреты по сравнению с теми, которые не были. Кроме того, он обнаружил, что гнезда в Сент -Денисе, Саскачеван использовал значительно меньше перьев, чем в долине Аннаполис , несмотря на то, что первые были дальше на север. Тем не менее, температура в Новой Шотландии (где находится Аннаполис -Вэлли), как правило, ниже, чем в Саскачеване, возможно, объясняя неожиданный результат. [ 34 ]

Во время ухаживания мужское дерево ласточки нападают на неизвестную женщину. Это может быть стимулировано через полете с флетином крыла женщиной, что может быть приглашением к суду. Затем мужчина может принять вертикальную позу, с поднятым и слегка распространенным хвостом, а крылья щелкнули и слегка опущены. Это побуждает женщину попытаться приземлиться на спину мужчины, но он летит, чтобы предотвратить это; это повторяется. Ухаживая за женщиной, мужчина летит на его выбранное место гнезда, которое женщина осматривает. Во время совокупления мужчина парит над женщиной, а затем крепко ее, когда звонит. Затем он вступает в контакт с клоакаль с женщиной, держа ее перья в шее в своем счете и стоя на ее слегка вытянутых крыльях. Копуляция происходит несколько раз. [ 7 ]
Яйца откладываются с начала мая до середины июня (хотя это происходит раньше из-за изменения климата [ 35 ] ) и птенцы усажены между серединой июля до июля. [ 14 ] Широта положительно коррелирует с датой укладки, [ 14 ] в то время как возраст женского пола и длина крыла (более длинные крылья позволяют более эффективно питаться [ 36 ] ) отрицательно коррелируют. [ 37 ] Ласточка дерева, вероятно, является заводчиком доходов , поскольку оно порождает, основываясь на численности пищи и температуры в течение сезона укладки. [ 36 ] Этот вид, как правило, является социально моногамным , но до 8% размножающихся мужчин полигин . [ 14 ] Полигиния под влиянием территории: мужчины, имеющие территории с гнездовыми ящиками, не менее 5 метров (16 футов) друг от друга, чаще являются полигинскими. [ 38 ] Предполагается, что это многоженство зависит от условий в течение сезона укладки: лучшие условия, такие как обилие пищи, позволяют женщинам в полигинском языке, которые не получают помощь в поиске пищи, чтобы отложить больше яиц. [ 39 ]

Ласточка дерева имеет высокие показатели отцовства в экстра-паре, от 38% до 69% птенцов являются продуктом лишнего отцовства, и от 50% до 87% выводков, содержащих по крайней мере одно птенцы, который был результатом линейной пары. совокупление. [ 14 ] Одним из факторов, который может способствовать этому, является то, что женщины контролируют совокупление, делая охранники отцовства неэффективными. [ 40 ] Это может быть смягчено более частыми копами незадолго до откладывания яиц, согласно исследованию 2009 года, в котором было обнаружено, что попытки сжимы в паре достигли пика за три-один день до того, как было отложено первое яйцо, и что более успешные попытки в течение этого периода увеличили долю внутри. -Па -пайр молодых мужчин имел. Этот последний вывод противоречит исследованию 1993 и 1994 года. [ 41 ] Вместобы отцовство не меняет уровень родительской помощи, который мужчина вносит в ласточку дерева. [ 42 ] Значительное количество отцов вне пары может быть поплавками (те, которые присутствуют на местах размножения, которые, по-видимому, не размножаются). Исследование 2001 года показало, что из 35 прихожан в экстра-паре 25 были обусловлены местными жителями, три жителями близлежащих участков и семь поплавчиков мужского пола. На ласточке дерева плавание, таким образом, помогает мужчинам в хорошем состоянии производить больше цыплят, позволяя мужчинам в плохом состоянии быть успешными, инвестируя в родительскую помощь. [ 43 ] Существует также значительная популяция женских поплавок; Исследование 1985 года подсчитано, что от 23% до 27% женщин были поплавки, из которых от 47% до 79% составляли подразделения. [ 44 ]
Почему женщины участвуют в дополнительной паре, и то, как они выбирают дополнительные пары, является спорным. Одна теория, называемая гипотезой генетической совместимости, утверждает, что повышенная приспособленность потомства является результатом повышенной гетерозиготности , и, следовательно, то, что женские деревья глотают, предпочли бы спариваться с мужчинами, которые менее генетически схожи с ними. Женщины также могут выбирать сперму после совокупления, чтобы обеспечить совместимый партнер. В поддержку этой теории, исследование 2007 года показало, что потомство Extrapair было более гетерозиготным, чем потомство внутри пары. [ 45 ] Тем не менее, статья 2005 года обнаружила небольшую негативную корреляцию между генетическим сходством пары и доли внедорожного молодого в их гнезде. [ 46 ] Теория хороших генов гласит, что женщины выбирают мужчин из пары на основе качества своих генов. Это объяснило бы, почему некоторые ласточки дерева не имеют молодых людей, тогда как другие делают. [ 40 ] Тем не менее, большинство исследований не обнаружили фенотипических различий между мужчинами вне пары и внутри пары (хотя исследование 2007 года показало, что пожилые мужчины с более ярким оперением были с большей вероятностью спариваться за пределами парной связи). [ 47 ] Кроме того, согласно тезисе 2017 года, потомство Extrapair не более склонна присоединиться к своему натальному населению, чем у потомства в паре. Другая теория предполагает, что отцовство в экстра-паре зависит от контекста, а молодые пары превосходят в пределах пары в определенных ситуациях и неэффективны в других средах. Например, диссертация в 2017 году обнаружила, что Extrapair Young был большим, тяжелее и более длительным крылом, чем внутри пары, когда оба были подвергнуты воздействию хищных креплений, в то время как внутри пары молодые были тяжелее, чем молодые, когда они были Показаны не-преседторующие крепления. Этот тезис также обнаружил, что внутри-пары молодые превзошли, вновь работали молодыми, с точки зрения жизни в жизни, когда они были подняты в менее переменных средах, что позволяет предположить, что у потомства в экстра-паре меньше пластичности развития, чем у потомства внутри пары. [ 48 ] Исследование в 2018 году слабо подтвердило эту зависимую от контекста гипотеза, обнаружив, что потомство вне пары с большей вероятностью залетает, чем у потомков в паре в экспериментально увеличенных расчетках; Тем не менее, ни длина теломер (коррелят выживаемости и репродуктивного успеха), ни размер 12 дней после вылупления не были значительно отличались среди этих молодых, и никаких существенных различий между двумя типами не было обнаружено в негарных выводках. [ 49 ]
Исследования, пытающиеся доказать адаптируемость отцовского отцовства для женщин, подверглись критике за отсутствие положительного эффекта, которое повысило бы приспособленность потомства по сравнению с потенциальной стоимостью снижения приспособленности для женщин, [ 50 ] такие как увеличение хищничества от поиска товарищей. [ 51 ] Таким образом, теории, основанные на неадаптивности отцовства для женщин, были постулированы. Эти теории основаны на генетическом ограничении , где поддерживается аллель, приводящий к неадаптивному поведению, потому что он также способствует полезному фенотипу. Теория межсексуальной антагонистической плейотропии говорит, что сильный отбор для отцовства вне пары у мужчин (как видно из этой птицы [ 50 ] ) переопределяет слабый отбор против лишнего отцовства у женщин. Тем временем гипотеза внутрисексуальной антагонистической плейотропии утверждает, что присутствует в восторге от отцовства, потому что гены, регулирующие ее, оказывают плейотропное воздействие на аспекты женской пригодности, [ 52 ] как в паре. [ 50 ]

Ласточка дерева накладывает сцепление от двух до восьми, хотя обычно от четырех до семи, [ 14 ] чистый белый и полупрозрачный при укладке, яйца, которые измеряют около 19 на 14 мм (0,75 на 0,55 дюйма). [ 53 ] Эти яйца инкубируются самками, [ 14 ] Обычно после укладывания яйца второго до конца, [ 54 ] в течение от 11 до 20 дней, [ 53 ] Хотя большинство вылупляется через 14-15 дней. Около 88% гнезд производят по крайней мере одно вмешательство, но это может быть снижено из -за плохой погоды и более молодой женщины. [ 14 ] Яйца обычно вылупляются в том порядке, в котором они были уложены. Они также вылупляются слегка асинхронно, в среднем 28 часов между тем, когда появляется первое и последнее птенд. Это может привести к весовой иерархии, где более ранние цыпочки весят больше (особенно в начале гнездового периода), чем те, которые вылуплены позже, что позволяет самке расставлять приоритеты, какой цыплят придаст пищу во время нехватки пищи. Это, вероятно, оказывает наибольшее влияние в начале гнездового периода, так как через 12 дней после вылупления обычно нет существенных различий в весах. [ 54 ] Инфункциональная личность цыплят и яиц иногда возникает, когда мужчина заменяется другим мужчиной. Инфункциональный лист обычно не происходит, когда сцепление не завершено, так как замена мужчин имеют возможность оплодотворить хотя бы одно яйцо. Когда мужчина прибывает во время инкубации, он иногда совершает детоубийство, но в других случаях принимает яйца, так как есть шанс, что некоторые яйца были связаны с заменой мужчины. Если замена мужчины прибывает после вылупления цыплят, обычно совершается детоубийство, хотя женщина иногда предотвращает это. [ 55 ]
Гнезда, производимые женщинами лучшего состояния, часто имеют соотношения полов, искаженные по отношению к высококачественным мужчинам. Исследование 2000 года предположило, что это связано с тем, что мужчины имеют более вариабельный репродуктивный успех, и поэтому высококачественный мужчина производит больше потомства, чем женщина с аналогичным качеством. [ 56 ]
На рост и выживание ласточек для жидкости влияют их окружающая среда. Как у молодых, так и в старших приходе (в возрасте от двух до четырех дней и между девятью до одиннадцати дней, соответственно) рост положительно влияет на более высокую максимальную температуру, особенно в первом. Более поздняя дата вылупления негативно влияет на рост, особенно для молодых птенцов. Пожилые птенцы растут несколько быстрее, когда насекомые обильны. Рост у более молодых птенцов увеличивается с возрастом, в то время как в старых птенцах он уменьшается, когда они становятся старше. [ 57 ] Молодые ласточки для деревьев способны терморегуляции как минимум 75% столь же эффективно, как взрослый в среднем возрасте 9,5 дней, когда он вышел из гнезда, и от четырех до восьми дней, когда он находится в гнезде (в зависимости от размера вывода). [ 58 ] Прихожие усажены примерно через 18-22 дней, при этом около 80% бегая успех. Как и успех вылупления, на это негативно влияет неблагоприятная погода и молодая женщина. [ 14 ] Цыплены могут быть охотятся змеями [ 59 ] и еноты. [ 60 ] Это хищничество может быть усугублена, попрошайчивая звонки. [ 61 ]
Кормление
[ редактировать ]
Дерево глотают кормы до 50 метров (160 футов) над землей отдельно или в группах. Его полет - смесь хлопья и скольжения. В течение сезона размножения это в основном в 4 километрах (2,5 мили) от места гнезда. Однако, когда он питается для птенцов, он обычно достигает 200 метров (660 футов) от гнезда, [ 14 ] В основном оставаться в поле зрения и корм на высоте до 12 метров (39 футов). [ 62 ] А также оказался в полете, [ 7 ] насекомых иногда берут с земли, воды, растительности и вертикальных поверхностей. [ 14 ]
Ласточка деревьев ест в основном насекомых, с некоторыми моллюсками, пауками и фруктами. В Северной Америке мухи составляют около 40% диеты, дополненные жуками и муравьями. В противном случае диета составляет около 90% мух. [ 14 ] Занимаемые насекомые представляют собой смесь водных и наземных организмов; Первые являются важным источником омега-3, очень ненасыщенных жирных кислот . Это связано с тем, что, хотя ласточка дерева может преобразовать предшественник α-линоленовую кислоту в высоко ненасыщенные жирные кислоты, такие как докозагексаеновая кислота , она не может сделать это в необходимых количествах. [ 63 ] Семена и ягода в основном из рода Myrica , который в основном взят во всех четырех сезонах северного полушария, кроме лета. Также было обнаружено, что ракообразные были важны в зимней рационе в исследовании на Лонг -Айленде, штат Нью -Йорк . [ 14 ]

Оба пола питаются птенцами (хотя самец кормит цыплят меньше, чем самки), в результате чего около 10-20 кормлений в час. [ 14 ] Родители часто используют призыв Чирп, чтобы стимулировать птенцов, чтобы попросить. Это используется чаще с более молодыми цыплятами, так как они просят меньше, чем пожилые птенцы, когда родитель прибывает с едой, но не звонит. [ 19 ] Вероятность попрошайничества в отсутствие родителей также увеличивается с возрастом. [ 64 ] Порядок вывода влияет на то, сколько питается цыпочка; Последние птенцы (в тех случаях, когда вылупление является асинхронным), вероятно, кормятся меньше, чем вылупленные ранее. [ 65 ] Прихоживание ближе к входу в гнездо также скорее кормят, как и те, кто сначала просит [ 66 ] и чаще. Общий показатель, при котором питается выводка, также увеличивается с большим количеством попрошайничества. [ 67 ] Сама диета состоит в основном из насекомых, тех, кто находится в приказе Diptera , Hemiptera и Odonata, составляющих большую часть диеты. [ 14 ] Эти насекомые в основном имеют размер до 10 мм (0,39 дюйма), но иногда длины до 60 мм (2,4 дюйма). [ 62 ] В гнездах возле озер подкижены людьми, кальциевые добавки, в первую очередь рыбные кости, экзоскелеты раков , раковины моллюсков и раковины птичьих яиц, труднее найти. Это заставляет взрослое дерево глотать проходить дальше, чем обычно - иногда до 650 м (2130 футов) от гнезда - чтобы получить эти добавки кальция. [ 68 ]
Выживание
[ редактировать ]Ласточка дерева имеет среднюю продолжительность жизни 2,7 года [ 69 ] и максимум 12 лет. Около 79% людей не переживают свой первый год, и те, которые сталкиваются с годовым уровнем смертности от 40% до 60%. [ 14 ] Большинство смертей, вероятно, являются результатом холодной погоды, что снижает доступность насекомых, что приводит к голоду. [ 7 ] Срок службы связана с длиной теломер: исследование 2005 года, в котором использовались показатели возврата (в место размножения предыдущего года) в качестве доверенности для выживания показали, что у тех, у кого самые длинные теломеры в один год, прогнозируемая продолжительность жизни 3,5 года. до 1,2 года для тех, у кого самые короткие теломеры. [ 70 ] Независимо от того, вызывают ли короткие теломеры снижение физической подготовки или просто их индикатор неизвестен. Несмотря на это, тезис 2016 года обнаружил, что показатели состояния были положительно коррелированы с длиной теломер. У мужчин также обычно были более длинные теломеры, чем у женщин, как и птицы меньшего крыла. Люди с более короткими теломер могут компенсировать потенциальные потери в пригодности путем увеличения репродуктивных усилий, тогда как люди с более длинными теломерами могут уменьшить свои инвестиции, о чем свидетельствует меньшая доля женщин -цыплят с более длинными теломерами. [ 71 ] Длина теломер очень наследуется и особенно зависит от длины матери. [ 72 ]
Хищничество
[ редактировать ]Ласточка дерева подвержена широкому диапазону хищников. Яйца, птенцы и взрослые в гнезде падают жертвами чернокожих змей крысы , американских ворон , американских пустрели , общих гракелей , северных блесков , бурундуков , деэрмис , домашних кошек , ласков , [ 73 ] Американские черные медведя , [ 74 ] и еноты . [ 75 ] Во время полета или расположенного, хищники на ласточку деревья включают американские пустельры, черные сороки , [ 73 ] Зарегистрированные Совы , [ 76 ] Великие рогатые совы , Мерлинс , Перегрин Соколы и ястребы с резким . В то время как уклончивой полет является обычным ответом на хищников в свободных ласточках, поведение моббинга распространено вокруг гнезда, [ 73 ] и направлен не только на хищников, но и на конкурентов сайта Nest, которые могут быть напуганы этим. [ 20 ] Такое поведение включает в себя ласточку, роясь и погружаясь в сторону (но на самом деле не поражение) злоумышленника [ 73 ] Примерно от 5 до 20 м (от 16 до 66 футов) над землей, обычно давая мягкие тикающие звонки в конце и приближаются примерно в 0,5 до 2 м (от 1,6 до 6,6 фута) от хищника. [ 20 ] Похоже, что это изменяет интенсивность своих атак в зависимости от того, к чему приближается хищник; [ 73 ] Исследование 1992 года показало, что хорьки вызвали более энергичную защиту, чем змеи черной крысы, [ 20 ] И тезис 2019 года также обнаружил, что модели черной крысиной змеи были погружались в модели по меньшей мере и восточные модели бурундуков больше всего. [ 77 ] Предполагается, что змея вызвало более слабый ответ, потому что защитное поведение может быть менее эффективным [ 20 ] и более опасно выступать против этого. [ 77 ]
Паразиты
[ редактировать ]
Ласточка дерева уязвима для различных паразитов, таких как трипаносома для паразита крови . Он также восприимчив к Idius Flea Edius и Peather Mites Pteronyssoides Tyrrelli , Trougeessartia и (вероятно) гемиальжа . Это также, вероятно, поражен вшами родов Брюелия и Мирсидеи . Существует корреляция между количеством блох на птице и количеством молодых, о которых она заботится. Предполагается, что эта связь возникает в результате улучшенного микроклимата для блох из -за большей сцепления. [ 78 ] Приходы также страдают от паразитов, таких как флоты рода Protocalliphora , [ 79 ] что приводит к потере крови птенцами. [ 80 ] Эти паразиты, однако, встречаются в большинстве гнезд и, по -видимому, не оказывают большого влияния на птенцов. Исследование, опубликованное в 1992 году, показало, что последствия паразитизма легли объясняют только около 5,5% от вариации в гнездовой массе. [ 79 ]
Иммунология
[ редактировать ]В племенной ласточке для женского дерева гуморальная иммунокомпетентность (HIC) обратно коррелирует с датой укладки. Это означает, что в среднем птица, которая откладывает свои яйца раньше, имеет более сильный антибиотический ответ на антиген , чем птица, которая лежит позже. Дерево ласточка, которая подвергается инвалидности с помощью срезания крыла, обычно имеет более низкий HIC. Эти отношения могут быть истолкованы как подтверждение вывода о том, что женщина, которая закладывает ранее, приобретает более высокий HIC, но авторы исследования обнаружили, что корреляции считают это маловероятным из -за более холодных температур в начале сезона размножения. Вместо этого они думали, что HIC может быть мерой качества, и что женщина с более высоким качеством может лежать раньше. Авторы также постулировали, что это индикатор рабочей нагрузки, как показано нижним HIC птиц с ограниченными возможностями. [ 81 ]
Женщины с более высоким качеством ласточки (как измеряются датой укладки) способны поддерживать свои репродуктивные усилия, направляя ресурсы для борьбы с иммунной проблемой. Болоты с более низким качеством менее способны это сделать; Исследование, проведенное в 2005 году в Итаке, штат Нью-Йорк, показало, что женщины с поздним слоем с искусственно увеличенным выводком, хотя способность поддерживать качество потомства, имели более низкие ответы на иммунный вызов, чем те, которые были более высокого качества или не имели увеличенного выводка. [ 82 ] Решает ли женщина определять приоритеты качества потомства или иммунокомпетентности, вероятно, связано с вероятностями выживания; Исследование 2005 года обнаружило, что женщины с увеличенным выводком в Араске, где выживаемость ниже, имели более слабые иммунные реакции, но продолжали устойчивые репродуктивные усилия, тогда как в Теннесси, с более высокими показателями выживаемости, имели более высокий ответ, но более низкий качественный потомство. [ 83 ]
На ласточке дерева некоторые компоненты иммунной системы ухудшаются с возрастом. Например, приобретенный иммунитет, опосредованный Т -клетками , снижается с возрастом на женской ласточке. Но возраст женщины не влияет как на приобретенный, так и врожденный гуморальный иммунитет ; Отсутствие ухудшения в первых контрастах с исследованиями на сараях ласточек и мухолочках с воротниками . [ 84 ] Из -за этого иммунозистенного (снижение иммунной функции с возрастом) пожилые женщины, инфицированные заболеванием, обычно посещают их гнездо меньше, что приводит к ростам медленнее. Они также могут похудеть из -за инфекции. [ 85 ]
Статус
[ редактировать ]Международное союз по сохранению природы считается наименее обеспокоенной международным союзом сохранения природы . Это связано с большим диапазоном птиц около 834 000 км 2 (322 000 кв. МИ), [ 1 ] и его стабильное население, оцениваемое около 20 000 000 человек. [ 14 ] Он защищен в США Законом о договоре о миграционных птицах 1918 года , [ 86 ] и в Канаде Законом о конвенции о перелетных птицах . [ 87 ] В некоторых частях США диапазон этой ласточки расширился на юг, вероятно, из -за изменений в землепользовании, реинтродукции бобров и гнезд, установленных для синих птиц . На ластость дерева негативно влияет очистка лесов и сокращение болот, последняя уменьшает среду обитания, доступную для зимы. Эта ласточка должна бороться за гнездовые участки с общим звездочком , домашним воробей (оба представлены в Северную Америку ), [ 14 ] синие птицы, [ 88 ] и дом Вран (который также уничтожает гнезда, не занимая их). [ 89 ] Подкисление озер может заставить эту ласточку пройти относительно большие расстояния, чтобы найти богатые кальцием предметы, и может привести к тому, что цыплят едят пластик. [ 68 ] Другие химические вещества, такие как пестициды и другие загрязняющие вещества, могут стать высоко сконцентрированными в яйцах, а ПХБ связаны с отказом от сцепления пары. [ 14 ] Загрязнение на местах нефтяных песков может негативно влиять на ласточки деревьев путем увеличения присутствия токсинов, измеряемой активностью этоксирезоруфин -ометилазы (детоксикационное фермент) у поддержанных. Обычно это мало влияет на уступка и бежание, [ 90 ] Хотя экстремальная погода может выявить последствия: исследование 2006 года показало, что птенцы из водно -болотных угодий, наиболее загрязненных материалом для обработки нефтяных песков, более чем в 10 раз чаще умирали, чем у контрольного участка в периоды синхронизированных холодных температур и сильных осадков по сравнению с Отсутствие различий в смертности между группами, когда погода была менее экстремальной. [ 91 ] Однако статья 2019 года показала, что увеличение осадков вызвало аналогичное снижение вылупления и успеха в птенцах для птенцов как вблизи, так и далеко от мест нефтяных песков. [ 92 ] В другом исследовании птицы, подвергшиеся воздействию ртути, в среднем, в среднем, на одну меньше цыпочки, чем те, что нет, эффект, усиленный теплой погодой. [ 93 ] Кроме того, холодные погодные явления могут быстро уменьшить доступность добычи воздушных насекомых, [ 94 ] А в некоторых популяциях с развитым репродукцией может привести к снижению выживаемости потомства. [ 95 ]
Ссылки
[ редактировать ]Примечания
[ редактировать ]Цитаты
[ редактировать ]- ^ Подпрыгнуть до: а беременный BirdLife International (2016). « Tachycineta Bicolor » . МСОП красный список угрожаемых видов . 2016 : E.T22712057A94316797. doi : 10.2305/iucn.uk.2016-3.rlts.t22712057a94316797.en . Получено 12 ноября 2021 года .
- ^ Vieillot, Луи Пьер (1807). Естественная история птиц Северной Америки: содержит большое количество видов, описанных или фигурированных впервые (на французском языке). Полет. 1. Париж: Краплет. п. 61.
- ^ Кабанис, Джин (1850). Музей Heineanum: Список орнитологической коллекции Оберамтмана Фердинанда Хейне на Gut St. Burchard перед Halberstatdt (на немецком языке). Том. 1. Хальберштадт: независимо от пополнения услуг Р. Франца. п. 48
- ^ Coues, Elliott (1878). Птицы долины Колорадо: хранилище научной и популярной информации, касающейся североамериканской орнитологии . Вашингтон, округ Колумбия: Правительственная типография.
- ^ Подпрыгнуть до: а беременный Coues, Elliott (1882). Проверенный список североамериканских птиц (2 изд.). Бостон: Эстес и Лауриат. п. 42
- ^ Тернер, Анжела (2020). Дель Хойо, Джозеф; Эллиот, Эндрю; Саругал, Горди; Кристиан, Дэвид А.; Дина, Эдуардо (ред.). Tachycineta leucorrhoa)"Полем Птицы птиц Lynx Edition. doi : 10.2173/ ow.whswa1 S2CID S2CID 14 2017января
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж k л м Тернер, Анжела (2010). Справочник по ласточкам и мартинскому миру . Лондон: A & C Black. С. 97–100. ISBN 978-1-4081-3172-5 .
- ^ Jobling, Джеймс А (2010). Хелм Словарь научных имен птиц . Лондон: Кристофер Хелм. с. 72, 377. ISBN 978-1-4081-2501-4 .
- ^ Cerasale, David J.; DOR, ROI; Винклер, Дэвид В.; Ловетт, Ирби Дж. (2012). «Филогения рода тахицинеты глобалов Нового Света: понимание полных митохондриальных геномов» (PDF) . Энергетический баланс у перелетных птиц: понимание митохондриальных геномов и биологии рецептора лептина (PhD). Корнелльский университет. С. 91–122.
- ^ Уиттингем, Линда А; Сликас, Бет; Винклер, Дэвид У; Шелдон, Фредерик Х (2002). «Филогения рода ласточки дерева, Tachycineta (Aves: Hirundinidae), байесовским анализом последовательностей митохондриальной ДНК». Молекулярная филогенетика и эволюция . 22 (3): 430–441. doi : 10.1006/mpev.2001.1073 . ISSN 1055-7903 . PMID 11884168 .
- ^ Мур, Уильям С. (1995). «Вывод филогений из вариации мтДНК: деревья митохондриальных генов по сравнению с деревьями ядерных генов». Эволюция 49 (4): 718–726. doi : 10.2307/2410325 . ISSN 0014-3820 . JSTOR 2410325 . PMID 28565131 .
- ^ DOR, ROI; Карлинг, Мэтью Д.; Ловетт, Ирби Дж.; Шелдон, Фредерик Х.; Винклер, Дэвид В. (2012). «Видовые деревья для деревьев ласточки (род Tachycineta ): альтернативная филогенетическая гипотеза митохондриальному генному дереву». Молекулярная филогенетика и эволюция . 65 (1): 317–322. doi : 10.1016/j.ympev.2012.06.020 . ISSN 1055-7903 . PMID 22750631 .
- ^ «Идентификация ласточки дерева, все о птицах, Корнелл -лаборатория орнитологии» . www.allaboutbirds.org . Получено 28 сентября 2020 года .
- ^ Подпрыгнуть до: а беременный в дюймовый и фон глин час я Дж k л м не а п Q. ведущий с Т в v В х и С Тернер, А. (2020). Дель Хойо, Джозеп; Эллиот, Эндрю; Саргатал, Джорди; Кристи, Давид А.; Де Хуана, Эдуардо (ред.). «Дерево ласточки ( Tachycineta Bicolor )» . Справочник по птицам мира живым . Барселона, Испания: Линкс Эдицины. doi : 10.2173/bow.treswa.01 . S2CID 216382529 . Получено 10 декабря 2017 года .
- ^ Подпрыгнуть до: а беременный Hussell, David JT (1983). «Возраст и цвет оперения у женских деревьев глотает». Журнал поля орнитологии . 54 (3): 312–318.
- ^ Stutchbury, Bridget J.; Робертсон, Роли Дж. (Октябрь 1987 г.). «Сигнализация подчиненного и женского статуса: две гипотезы для адаптивной значимости оперения субадульта у женских ласточек» (PDF) . Аук . 104 (4): 717–723. doi : 10.1093/auk/104.4.717 . ISSN 0004-8038 .
- ^ Коади, Челси Д.; Доусон, Рассел Д. (2013). «Осветный цвет оперения в салоне женских деревьев ( Tachycineta Bicolor ) уменьшает конспецифическую агрессию в течение сезона размножения». Wilson Journal of Ornithology . 125 (2): 348–357. doi : 10.1676/12-155.1 . ISSN 1559-4491 . S2CID 85030082 .
- ^ Подпрыгнуть до: а беременный в Шарман, мой; Робертсон, RJ; Рэтклифф, Л.М. (1994). «Вокализации ласточки дерева ( Tachycineta Bicolor ) в течение предварительного периода: структурный и контекстуальный анализ». Американский Мидленд натуралист . 132 (2): 264–274. doi : 10.2307/2426582 . ISSN 0003-0031 . JSTOR 2426582 .
- ^ Подпрыгнуть до: а беременный Леонард, Марти Л.; Фернандес, Николь; Браун, Глен (1997). «Родительские звонки и поведение в птенцах в деревьях ласточек» . Аук . 114 (4): 668–672. doi : 10.2307/4089286 . ISSN 0004-8038 . JSTOR 4089286 .
- ^ Подпрыгнуть до: а беременный в дюймовый и Винклер, Дэвид В. (1992). «Причины и последствия вариации в поведении родителей по пролавкам деревьев». Кондор . 94 (2): 502–520. doi : 10.2307/1369222 . ISSN 0010-5422 . JSTOR 1369222 . S2CID 45389696 .
- ^ Макинтайр, Эмма; Хорн, Эндрю Дж.; Леонард, Марти Л. (2014). «Ответ ли ласточки для птенцов ( Tachycineta Bicolor ) отвечает на звонки по родительской тревоге?» Полем Аук . 131 (3): 314–320. doi : 10.1642/auk-13-235.1 . ISSN 0004-8038 . S2CID 86754716 .
- ^ Макинтайр, Эмма; Леонард, Марти Л.; Хорн, Эндрю Г. (2014). «Окружающий шум и родительское общение риска хищничества в деревьях, тахицинета биколор ». Поведение животных . 87 : 85–89. doi : 10.1016/j.anbehav.2013.10.013 . ISSN 0003-3472 . S2CID 53173370 .
- ^ Леонард, Марти Л.; Хорн, Эндрю Дж.; Освальд, Криста Н.; Макинтайр, Эмма (2015). «Влияние окружающего шума на взаимодействие родителей с пружином на ласточках деревьев». Поведение животных . 109 : 1–7. doi : 10.1016/j.anbehav.2015.07.036 . ISSN 0003-3472 . S2CID 53160897 .
- ^ Рыцарь, Саманта М.; Брэдли, Дэвид В.; Кларк, Роберт Дж.; Гоу, Элизабет А.; Bélisle, Marc; Berzins, Lisha L.; Блейк, Триша; Мост, Эли С.; Берк, Лорен; Доусон, Рассел Д.; Данн, Питер О.; Гарант, Дэни; Холройд, Джеффри Л.; Хусселл, Дэвид Дж. Лансдорп, Ольга; Laughlin, Andrew J.; Леонард, Марти Л.; Пеллетье, Фан; Шеплер, Дэйв; Сифферман, Линн; Тейлор, Каз М.; Trefry, Helen E.; Влек, Кэрол М.; Влек, Дэвид; Винклер, Дэвид В.; Уиттингем, Линда А.; Норрис, Д. Райан (2018). «Создание и оценка сети миграционных певцов в целом по всему континенту в годовом цикле» . Экологические монографии . 88 (3): 445–460. doi : 10.1002/ecm.1298 . HDL : 10214/11411 . ISSN 0012-9615 .
- ^ Подпрыгнуть до: а беременный Винклер, Дэвид В.; Wrege, Peter H.; Аллен, Пол Э.; Каст, Трейси Л.; Senesac, Pixie; Уоссон, Мэтью Ф.; Llambías, Paulo E.; Ферретти, Валентина; Салливан, Патрик Дж. (2004). «Разведение рассеяния и филопатрии в ласточке дерева». Кондор (представленная рукопись). 106 (4): 768–776. doi : 10.1650/7634 . ISSN 0010-5422 . S2CID 42958338 .
- ^ Винклер, Дэвид В. (2006). «Разрушители и миграции ласточек» . El Hornero . 21 (2): 85–97. doi : 10.56178/eh.v21i2.790 . HDL : 20.500.12110/hornero_v021_n02_p085 .
- ^ Джонс, Джейсон (2003). «Дерево ласточки ( Tachycineta Bicolor ): новый модельный организм?». Аук . 120 (3): 591. doi : 10.1642/0004-8038 (2003) 120 [0591: tstban] 2.0.co; 2 . ISSN 0004-8038 . S2CID 36003514 .
- ^ Шеплер, Дэйв; Кларк, Роберт Дж. (2003). «Причины и последствия рассеивания по ласточке деревьев ( Tachycineta Bicolor ) в Саскачеване». Аук . 120 (3): 619–631. doi : 10.1642/0004-8038 (2003) 120 [0619: Cacots] 2.0.co; 2 . ISSN 0004-8038 . S2CID 28251342 .
- ^ Малдал, Элисон; Гиббс, Х. Лисл; Робертсон, Роли Дж. (1985). «Предпочтительное расстояние между гнездами птицы с облигацией полости, ласточка». Кондор . 87 (3): 356–363. doi : 10.2307/1367216 . ISSN 0010-5422 . JSTOR 1367216 .
- ^ Ренделл, Уоллес Б.; Робертсон, Роли Дж. (1989). «Характеристики на сайте гнездо, репродуктивный успех и доступность полости для лесов, размножающихся в естественных полостях». Кондор . 91 (4): 875. doi : 10.2307/1368072 . ISSN 0010-5422 . JSTOR 1368072 .
- ^ Подпрыгнуть до: а беременный Винклер, Дэвид В. (1993). «Использование и важность перьев в качестве гнездовой подкладки в ласточках деревьев ( Tachycineta Bicolor )». Аук . 110 (1): 29–36.
- ^ Ломбардо, Майкл П.; Босман, Рут М.; Фаро, Кристина А.; Houtteman, Stephen G.; Kluisza, Timothy S. (1995). «Влияние перьев в качестве изоляции гнезда на инкубационное поведение и репродуктивные характеристики ласточек деревьев ( Tachycineta Bicolor )» . Аук . 112 (4): 973–981. doi : 10.2307/4089028 . ISSN 0004-8038 . JSTOR 4089028 . S2CID 55119375 .
- ^ Виндзор, Ребекка Л.; Fegely, Jessica L.; Ардия, Даниэль Р. (2013). «Влияние размера гнезда и изоляции на тепловые свойства гнезд ласточки». Журнал птичьей биологии . 44 (4): 305–310. doi : 10.1111/j.1600-048x.2013.05768.x . ISSN 0908-8857 .
- ^ Голландия, Эрика Р.; Шатлер, Дэйв (2018). «Гнездо перенащиеся реакции по ласточкам деревьев ( Tachycineta Bicolor ) на экспериментальное потепление». Журнал орнитологии . 159 (4): 991–998. doi : 10.1007/S10336-018-1568-6 . ISSN 2193-7192 . S2CID 46971093 .
- ^ Данн, По; Винклер, DW (1999). «Изменение климата повлияло на дату размножения деревьев по всей Северной Америке» . Труды Королевского общества B: Биологические науки . 266 (1437): 2487–2490. doi : 10.1098/rspb.1999.0950 . ISSN 0962-8452 . PMC 1690485 . PMID 10693819 .
- ^ Подпрыгнуть до: а беременный NOOKER, Жаклин К.; Данн, Питер О.; Уиттингем, Линда А. (2005). «Влияние численности пищи, погоды и состояния женского пола на размножение на ласточках деревьев ( Tachycineta Bicolor )» . Аук . 122 (4): 1225. DOI : 10.1642/0004-8038 (2005) 122 [1225: eofawa] 2.0.co; 2 . ISSN 0004-8038 . S2CID 4995233 .
- ^ Винклер, Дэвид В.; Аллен, Пол Э. (1996). «Сезонное снижение размера сцепления деревьев: физиологическое ограничение или стратегическая корректировка?». Экология . 77 (3): 922–932. doi : 10.2307/22655512 . ISSN 0012-9658 . JSTOR 2265512 . S2CID 84919928 .
- ^ Данн, Питер О.; Хэннон, Сьюзен Дж. (1991). «Внутривидочная конкуренция и поддержание моногамии в деревьях ласточек». Поведенческая экология . 2 (3): 258–266. doi : 10.1093/beheco/2.3.258 . ISSN 1045-2249 .
- ^ Данн, Питер О.; Хэннон, Сьюзен Дж. (1992). «Влияние численности пищи и ухода за родителями мужчин на репродуктивный успех и моногамию в ласточках деревьев». Аук . 109 (3): 488–499. ISSN 0004-8038 .
- ^ Подпрыгнуть до: а беременный Lifjeld, Jan T.; Данн, Питер О.; Робертсон, Роли Дж.; Boag, Peter T. (1993). «Внешнее отцовство в моногамных деревьях глотает». Поведение животных . 45 (2): 213–229. doi : 10.1006/anbe.1993.1028 . ISSN 0003-3472 . S2CID 53195131 .
- ^ Кроу, Сьюзен А.; Kleven, Oddmund; Delmore, Kira E.; Laskemoen, Terje; Nocera, Joseph J.; Lifjeld, Jan T.; Робертсон, Роли Дж. (2009). «Обеспечение отцовства посредством частых копа в дикой прохожности с интенсивной конкуренцией сперматозоидов». Поведение животных . 77 (1): 183–187. doi : 10.1016/j.anbehav.2008.09.024 . ISSN 0003-3472 . S2CID 13722818 .
- ^ Kempenaers, Bart; Lanctot, Richard B.; Робертсон, Роли Дж. (1998). «Уверенность отцовства и отцовских инвестиций в восточные голубые птицы и деревьев». Поведение животных . 55 (4): 845–860. doi : 10.1006/anbe.1997.0667 . ISSN 0003-3472 . PMID 9632472 . S2CID 24760456 .
- ^ Kempenaers, Bart; Эвердинг, Сьюзи; Епископ, Шерил; Боаг, Петр; Робертсон, Роли Дж. (2001). «Восторское отцовство и репродуктивная роль мужских поплавок в ласточке дерева ( Tachycineta Bicolor )». Поведенческая экология и социобиология . 49 (4): 251–259. doi : 10.1007/s0026550000305 . ISSN 0340-5443 . S2CID 25483760 .
- ^ Stutchbury, Bridget J.; Робертсон, Роли Дж. (1985). «Плавающие популяции женского дерева глотают». Аук . 102 (3): 651–654. doi : 10.1093/auk/102.3.651 . ISSN 0004-8038 .
- ^ Стэплтон, Мэри К.; Kleven, Oddmund; Lifjeld, Jan T.; Робертсон, Роли Дж. (2007). «Женское дерево глотает ( Tachycineta Bicolor ) увеличивает гетерозиготность потомства за счет экстрапаитного спаривания». Поведенческая экология и социобиология . 61 (11): 1725–1733. doi : 10.1007/s00265-007-0404-4 . ISSN 0340-5443 . S2CID 22513713 .
- ^ Барбер, Коллин А.; Эдвардс, Мэнди Дж.; Робертсон, Роли Дж. (2005). «Испытание гипотезы генетической совместимости с деревьями ласточек, Tachycineta Bicolor ». Канадский журнал зоологии . 83 (7): 955–961. doi : 10.1139/Z05-091 . ISSN 0008-4301 .
- ^ Биттон, Пьер-Поль; О'Брайен, Эрин Л.; Доусон, Рассел Д. (2007). «Яркость оперения и возраст прогнозируют успех оплодотворения экстрапаиров мужских деревьев, биколор тахицинеты ». Поведение животных . 74 (6): 1777–1784. doi : 10.1016/j.anbehav.2007.03.018 . ISSN 0003-3472 . S2CID 53170228 .
- ^ Халлингер, Келли Кристен (2017). Контекстно-зависимые последствия для подготовки к отцовству в экстра-паре на ласточках деревьев ( Tachycineta Bicolor ) (PhD). Корнелльский университет .
- ^ Belmaker, Amos; Халлингер, Келли К.; Глинн, Ребекка А.; Haussmann, Mark F.; Винклер, Дэвид В. (2018). «Существует ли контекстно-зависимое преимущество спаривания вне пары в ласточках деревьев?». Аук . 135 (4): 998–1008. doi : 10.1642/AUK-18-3.1 . HDL : 10150/631782 . ISSN 0004-8038 . S2CID 91253157 .
- ^ Подпрыгнуть до: а беременный в Arnqvist, Göran; Киркпатрик, Марк (2005). «Эволюция неверности в социально моногамных пробирках: сила прямого и косвенного отбора при поведении экстрапаиров у женщин». Американский натуралист . 165 (S5): S26 - S37. doi : 10.1086/429350 . ISSN 0003-0147 . PMID 15795859 . S2CID 32810854 .
- ^ Данн, Питер О.; Уиттингем, Линда А. (2006). «Затраты поиска влияют на пространственное распределение, но не на уровень, а также спаривание в экстра-паре в ласточках деревьев». Поведенческая экология и социобиология . 61 (3): 449–454. doi : 10.1007/s00265-006-0272-3 . ISSN 0340-5443 . S2CID 4975256 .
- ^ Forstmeier, Wolfgang; Накагава, Шиничи; Гриффит, Саймон С.; Kempenaers, Bart (2014). «Женское сопряжение в экстра-паре: адаптация или генетическое ограничение?». Тенденции в экологии и эволюции . 29 (8): 456–464. doi : 10.1016/j.tree.2014.05.005 . ISSN 0169-5347 . PMID 24909948 .
- ^ Подпрыгнуть до: а беременный Хаубер, Марк Э. (1 августа 2014 г.). Книга яиц: руководство по размеру в натуральную величину по яйцам шестисот видов птиц мира . Чикаго: Университет Чикагской Прессы. п. 448. ISBN 978-0-226-05781-1 .
- ^ Подпрыгнуть до: а беременный Clotfelter, Ethan D.; Уиттингем, Линда А.; Данн, Питер О. (2000). «Порядок укладывания, вылупление асинхронности и масса тела в притяжении в деревьях проглатывает Tachycineta Bicolor ». Журнал птичьей биологии . 31 (3): 329–334. doi : 10.1034/j.1600-048x.2000.310308.x . ISSN 0908-8857 .
- ^ Робертсон, Роли Дж. (1990). «Тактика и контр-таблица сексуально отобранного детоубийства в ласточках деревьев». Популяционная биология пассинских птиц: интегрированный подход . Серия НАТО ASI. С. 381–390. doi : 10.1007/978-3-642-75110-3_32 . ISBN 978-3-540-51759-7 .
- ^ Уиттингем, Линда А.; Данн, Питер О. (2000). «Строительные соотношения потомства на деревьях ласточек: женщины в лучшем состоянии производят больше сыновей». Молекулярная экология . 9 (8): 1123–1129. doi : 10.1046/j.1365-294x.2000.00980.x . ISSN 0962-1083 . PMID 10964231 . S2CID 3720108 .
- ^ Маккарти, Джон П.; Винклер, Дэвид В. (2008). «Относительная важность переменных окружающей среды в определении роста гнездового дерева проглатывает биколор тахицинеты ». Ибис . 141 (2): 286–296. doi : 10.1111/j.1474-919x.1999.tb07551.x . ISSN 0019-1019 .
- ^ Данн, Эрика Х. (1979). «Возраст эффективной гомеотермии в птенцах прощела в зависимости от размера выводка». Бюллетень Уилсона . 91 (3): 455–457. ISSN 0043-5643 .
- ^ Де Стивен, Дайан (1980). «Размер сцепления, успех размножения и выживание родителей на ласточке дерева ( биколор иридопрокнов )». Эволюция 34 (2): 278–291. doi : 10.1111/j.1558-5646.1980.tb04816.x . HDL : 2027.42/137562 . ISSN 0014-3820 . PMID 28563429 . S2CID 19454000 .
- ^ Робертсон, Роли Дж.; Ренделл, Уоллес Б. (1990). «Сравнение экологии размножения вторичной полости, гнездовая птица, ласточка дерева ( биколор тахицинета ), в гнездовых ящиках и природных полостях». Канадский журнал зоологии . 68 (5): 1046–1052. doi : 10.1139/Z90-152 . ISSN 0008-4301 .
- ^ Пиянка, Сьюзен М.; Леонард, Марти Л. (1997). «Стремление и риск хищничества у брюцких птиц» . Поведенческая экология . 8 (6): 644–646. doi : 10.1093/beheco/8.6.644 . ISSN 1045-2249 .
- ^ Подпрыгнуть до: а беременный Маккарти, Джон П.; Винклер, Дэвид В. (1999). «Экология и селективность пищи деревьев, питающих птенцов» . Кондор . 101 (2): 246–254. doi : 10.2307/1369987 . ISSN 0010-5422 . JSTOR 1369987 .
- ^ Twining, Cornelia W.; Лоуренс, Петр; Винклер, Дэвид В.; Flecker, Alexander S.; Бренна, Дж. Томас (2018). «Эффективность конверсии α-линоленовой кислоты в омега-3 высоко ненасыщенные жирные кислоты у аэрофотояжных цыплят» . Журнал экспериментальной биологии . 221 (3): JEB165373. doi : 10.1242/jeb.165373 . ISSN 0022-0949 . PMID 29217628 .
- ^ Леонард, Марти Л.; Хорн, Эндрю Г. (2001). «Смотреть в отсутствие родителей, поглощающих дерево» . Поведенческая экология . 12 (4): 501–505. doi : 10.1093/beheco/12.4.501 . ISSN 1465-7279 .
- ^ Джонсон, Л. Скотт; Wimmers, Larry E.; Кэмпбелл, Сара; Гамильтон, Люси (2003). размер и полов Скорость роста , « Журнал птичьей биологии . 34 (1): 35–43. doi : 10.1034/j.1600-048x.2003.02950.x . ISSN 0908-8857 .
- ^ Уиттингем, Линда А; Данн, Питер О; Clotfelter, Ethan D (2003). «Распределение родительской пищи для птенцов по ладошам: влияние гнездового поведения, пола и отцовства». Поведение животных . 65 (6): 1203–1210. doi : 10.1006/anbe.2003.2178 . ISSN 0003-3472 . S2CID 4993059 .
- ^ Леонард, ML; Хорн, Эндрю Г. (2001). «Промогание звонков и решения о кормлении родителей в деревьях проглатывания ( Tachycineta Bicolor )». Поведенческая экология и социобиология . 49 (2–3): 170–175. doi : 10.1007/s0026550000290 . ISSN 0340-5443 . S2CID 25507198 .
- ^ Подпрыгнуть до: а беременный Сент -Луис, Винсент Л.; Breabaart, Loes (1991). «Добавки кальция в рационе гнездового дерева глотают вблизи кислотных озер». Кондор . 93 (2): 286–294. doi : 10.2307/1368944 . ISSN 0010-5422 . JSTOR 1368944 . S2CID 87271779 .
- ^ Влек, Кэрол М.; Влек, Дэвид; Паласиос, Мария Г. (2011). «Эволюционная экология старения: тематическое исследование с использованием ласточек деревьев, тахицинета биколор ». Журнал орнитологии . 152 (S1): 203–211. doi : 10.1007/s10336-010-0629-2 . ISSN 0021-8375 . S2CID 25321642 .
- ^ Haussmann, Mark F.; Винклер, Дэвид В.; Влек, Кэрол М. (22 июня 2005 г.). «Более длинные теломеры, связанные с более высокой выживаемостью у птиц» . Биологические письма . 1 (2): 212–214. doi : 10.1098/rsbl.2005.0301 . ISSN 1744-9561 . PMC 1626238 . PMID 17148169 .
- ^ Belmaker, Amos (2016). «Модели ко-вариации длины теломер, состояния, истории жизни и пригодности у недолговечных видов птиц». Роль длины теломер в поведении по ласточке деревьев и истории жизни (докторская степень). Корнелльский университет.
- ^ Belmaker, Amos; Халлингер, Келли К.; Глинн, Ребека А.; Винклер, Дэвид В.; Haussmann, Mark F. (2019). «Экологические и генетические детерминанты длины теломеры цыплят на ласточках деревьев ( Tachycineta Bicolor )» . Экология и эволюция . 9 (14): 8175–8186. doi : 10.1002/ECE3.5386 . ISSN 2045-7758 . PMC 6662556 . PMID 31380080 .
- ^ Подпрыгнуть до: а беременный в дюймовый и Винклер, DW; Халлингер, KK; Ардия, доктор; Робертсон, RJ; Stutchbury, BJ; Chohen, RR (2011). Пул, AF, ed. «Дерево ласточки ( Tachycineta Bicolor )». Птицы Северной Америки . Итака, Нью -Йорк : Корнелл -лаборатория орнитологии.
- ^ Зак, Рето; Mayoh, Keith R. (1984). «Гамма -радиационное воздействие на петлишние деревья прощеивает». Экология . 65 (5): 1641–1647. doi : 10.2307/1939142 . ISSN 0012-9658 . JSTOR 1939142 .
- ^ Чепмен, LB (1955). «Исследования колонии по ласточке деревьев». Птиц . 6 (2): 45–70.
- ^ Errington, PL (1932). «Пищевые привычки южного Висконсинского хищника. Часть I. Совы». Кондор . 34 (4): 176–186.
- ^ Подпрыгнуть до: а беременный Маас, Наталья Мэй (2019). Воспринимаемый риск хищничества и ответы взрослых и птенцов ласточек ( Tachycineta Bicolor ) ( MS ). Восточный университет Кентукки .
- ^ Шеплер, Дэйв; Малли, Адель; Кларк, Роберт Дж. (2004). «Дерево ласточки репродуктивные инвестиции, стресс и паразиты». Канадский журнал зоологии . 82 (3): 442–448. Citeseerx 10.1.1.530.1736 . doi : 10.1139/Z04-016 . ISSN 0008-4301 .
- ^ Подпрыгнуть до: а беременный Роби, Даниэль Д.; Бринк, Карен Л.; Виттманн, Карин (1992). «Влияние паразитизма птиц на восточную синюю птицу и прихожания по ласточке деревьев» (PDF) . Бюллетень Уилсона . 104 (4): 630–643.
- ^ Desimone, Joely G.; Clotfelter, Ethan D.; Черный, Элизабет С.; Кнути, Сара А. (2018). «Избегание, толерантность и сопротивление эктопаразитам в приходе и взрослых деревьях». Журнал птичьей биологии . 49 (2): JAV - 01641. doi : 10.1111/jav.01641 . ISSN 0908-8857 .
- ^ Хассельквист, Деннис; Уоссон, Мэтью Ф.; Винклер, Дэвид В. (2001). «Гуморальная иммунокомпетентность коррелирует с датой яйцеклетой и отражает рабочую нагрузку на женских лесах» . Поведенческая экология . 12 (1): 93–97. doi : 10.1093/oxfordjournals.beheco.a000384 . ISSN 1465-7279 .
- ^ Ардия, Даниэль Р. (2005). «Индивидуальное качество опосредует компромиссы между репродуктивными усилиями и иммунной функцией в ласточках деревьев» . Журнал экологии животных . 74 (3): 517–524. doi : 10.1111/j.1365-2656.2005.00950.x . ISSN 0021-8790 .
- ^ Ардия, Даниэль Р. (2005). «Деревные ласточки обменяют иммунную функцию и репродуктивные усилия по -разному в их диапазоне». Экология . 86 (8): 2040–2046. doi : 10.1890/04-1619 . ISSN 0012-9658 . S2CID 33354318 .
- ^ Паласиос, Мария Г; Кунник, Джоан Е; Винклер, Дэвид У; Влек, Кэрол М. (2007). «Иммунозвуковое возражение в некоторых, но не на всех иммунных компонентах в свободно жившем позвоночном, дереве глотает» . Труды Королевского общества B: Биологические науки . 274 (1612): 951–957. doi : 10.1098/rspb.2006.0192 . ISSN 0962-8452 . PMC 2141670 . PMID 17251097 .
- ^ Паласиос, Мария Г.; Винклер, Дэвид В.; Класинг, Кирк С.; Хассельквист, Деннис; Влек, Кэрол М. (2011). «Последствия старения иммунной системы в природе: исследование затрат на иммунозвучное разрыв в свободно жидких деревьях» . Экология (представленная рукопись). 92 (4): 952–966. doi : 10.1890/10-0662.1 . ISSN 0012-9658 . PMID 21661557 .
- ^ «Закон о мигрирующих договорах о птицах, защищенных видах (10.13)» . Американская рыба и дикая природа. 2013 . Получено 4 июня 2018 года .
- ^ «Птицы защищены в соответствии с Законом о конвенции о перелетных птицах» . Правительство Канады. 2017. Архивировано из оригинала 20 мая 2019 года . Получено 21 июля 2018 года .
- ^ Вибе, Карен Л. (2016). «Межвидовая конкуренция за гнезда: предшествующее владение превосходит потенциал ресурсов для гору -синей птицы, конкурирующей с ласточкой деревьев» . Аук . 133 (3): 512–519. doi : 10.1642/auk-16-25.1 . ISSN 0004-8038 .
- ^ Финч, Дебора М. (1990). «Влияние хищничества и вмешательства конкурентов на успех гнездования домашних руль и деревьев». Кондор . 92 (3): 674–687. doi : 10.2307/1368686 . ISSN 0010-5422 . JSTOR 1368686 .
- ^ Smits, Judit E.; Уэйленд, Марк Э.; Миллер, Майкл Дж.; Либер, Карстен; Трюдо, Сюзанна (2000). «Репродуктивные, иммунные и физиологические конечные точки в ласточках деревьев на мелиорированных шахтных площадках нефтяных песков». Экологическая токсикология и химия . 19 (12): 2951–2960. doi : 10.1002/и т. Д. 5620191216 . ISSN 0730-7268 . S2CID 85573498 .
- ^ Gentes, Marie-Line; Уолднер, Шерил; Папп, Зсузсанна; Smits, Judit EG (2006). «Влияние хвостов нефтяных песков и суровых погодных условий на показатели смертности, усилия по росту и детоксикации на ласточках для жидких деревьев ( Tachycineta Bicolor )». Загрязнение окружающей среды . 142 (1): 24–33. doi : 10.1016/j.envpol.2005.09.013 . ISSN 0269-7491 . PMID 16297515 .
- ^ Годвин, Кристина М.; Барклай, Роберт М.Р.; Smits, Judit EG (2019). «Дерево ласточки ( Tachycineta Bicolor ) Успех гнездо и рост гнездования возле добычи нефтяных песков в северо -восточной Альберте, Канада» . Канадский журнал зоологии . 97 (6): 547–557. doi : 10.1139/cjz-2018-0247 . ISSN 0008-4301 .
- ^ Халлингер, Келли К.; Кристол, Даниэль А. (2011). «Роль погоды в опосредовании влияния воздействия ртути на репродуктивный успех на ласточках деревьев». Экотоксикология . 20 (6): 1368–1377. doi : 10.1007/s10646-011-0694-1 . ISSN 0963-9292 . PMID 21553259 . S2CID 42492417 .
- ^ Винклер, Дэвид В.; Ло, Майлз К.; Ракхимбердиев, Эльдар (сентябрь 2013 г.). «Влияние температуры на пищевые продукты и смертность Chick в деревьях ласточек (Tachycineta Bicolor)» . Oecologia . 173 (1): 129–138. BIBCODE : 2013OECOL.173..129W . doi : 10.1007/s00442-013-2605-z . ISSN 0029-8549 . PMC 3751296 . PMID 23468236 .
- ^ Шипли, Дж. Райан; Twining, Cornelia W.; Тафф, Конор С.; VitOUSEK, Maren N.; Флэк, Андреа; Винклер, Дэвид В. (28 сентября 2020 г.). «Птицы, продвигающиеся даты, с потеплением источников сталкиваются с большим риском смертности в цыпочках» . Труды Национальной академии наук . 117 (41): 25590–25594. Bibcode : 2020pnas..11725590S . doi : 10.1073/pnas.2009864117 . ISSN 0027-8424 . PMC 7568286 . PMID 32989166 . S2CID 222159036 .
Внешние ссылки
[ редактировать ]| Базы данных управления авторитетом : национальный |
|---|