Хромиста
| Хромиста | |
|---|---|
 | |
| Хромиста коллаж | |
| Научная классификация (устаревший) | |
| Домен: | Эукариоты |
| Клэйд : | потогонные средства |
| Королевство: | Хромиста Кавалер-Смит 1981 г. |
| Группы включены | |
| Кладистически включенные, но традиционно исключенные таксоны | |
Chromista - предложенный, но полифилетический вариант. [1] [2] [3] биологическое царство , выделенное из Chromalveolata , состоящее из одноклеточных и многоклеточных эукариотических видов, которые имеют схожие особенности фотосинтетических органелл ( пластид ). [4] К нему относятся все эукариоты, пластиды которых содержат хлорофилл С и окружены четырьмя мембранами. Если предок уже обладал хлоропластами, полученными в результате эндосимбиоза из красных водорослей , все нефотосинтезирующие хромисты вторично утратили способность к фотосинтезу. Ее члены могли возникнуть независимо как отдельные эволюционные группы от последнего общего предка эукариот. [1]
Chromista как таксон был создан британским биологом Томасом Кавалье-Смитом в 1981 году для различения страменопилей , гаптофитов и криптофитов . [5] По мнению Кавальера-Смита, первоначально царство состояло в основном из фотосинтезирующих эукариотов ( водорослей ), но позднее он ввёл множество гетеротрофов ( простейших в предложенную группу ). По состоянию на 2018 год королевство было почти таким же разнообразным, как королевства Plantae и Animalia, и состояло из восьми типов. Известные представители включают морские водоросли, ожог , динофлагелляты , парамеции , мозговой паразит токсоплазму . и малярийный паразит плазмодий картофельный [6]
Однако гипотеза Кавальер-Смита о монофилии хромистов была отвергнута другими исследователями, которые считают более вероятным, что некоторые хромисты приобрели свои пластиды путем включения другого хромиста, а не унаследовали их от общего предка. Считается, что это происходило неоднократно, так что красные пластиды распространялись от одной группы к другой. Пластиды не только не характеризуют своих хозяев как принадлежащих к одной кладе, но и имеют историю, отличную от истории своих разрозненных хозяев. Они, по-видимому, произошли от Rhodophytina и перешли к Cryptophytina , а от них к Ochrophyta и Haptophyta , а затем от последних к Myzozoa . [3]
Биология
[ редактировать ]
Члены Chromista — это одноклеточные и многоклеточные эукариоты, обладающие в основном одной или обеими характеристиками: [5]
- пластиды, которые содержат хлорофилл c и лежат внутри дополнительной (перипластидной) мембраны в просвете шероховатой эндоплазматической сети (обычно внутри перинуклеарной цистерны);
- реснички с трехраздельными или двураздельными жесткими трубчатыми волосками.
В царство входят разнообразные организмы от водорослей до малярийных паразитов ( плазмодий ). [7] Молекулярные данные показывают, что пластиды хромистов произошли от красных водорослей в результате вторичного симбиогенеза в одном событии. [8] Напротив, растения приобрели пластиды от цианобактерий посредством первичного симбиогенеза . [9] Эти пластиды теперь заключены в две дополнительные клеточные мембраны, образуя четырехмембранную оболочку, в результате чего они приобрели множество других мембранных белков для транспортировки молекул в органеллы и из них. Предполагается, что разнообразие хромистов возникло в результате дегенерации, потери или замены пластид в некоторых линиях. [10] Дополнительный симбиогенез зеленых водорослей обеспечил сохранение генов у некоторых представителей (например, у гетероконтов), [11] и бактериальный хлорофилл (на что указывает наличие гена рибосомального белка L36, rpl36 ) у гаптофитов и криптофитов. [12]
История и группы
[ редактировать ]Ниже показаны некоторые примеры классификации участвующих групп, которые имеют перекрывающееся, но неидентичное членство. [13] [14]
Chromophycées (Шадефо, 1950)
[ редактировать ]Хромофицеи (Шадефо, 1950), [15] переименован в Chromophycota (Chadefaud, 1960), [16] включали современные Ochrophyta (автотрофные Stramenopiles ), Haptophyta (входили в состав Chrysophyceae до Christensen, 1962), Cryptophyta , Dinophyta , Euglenophyceae и Choanoflagellida (входили в состав Chrysophyceae до Hibberd, 1975).
Chromophyta (Кристенсен 1962, 1989)
[ редактировать ]Chromophyta (Christensen 1962, 2008), определяемая как водоросли с хлорофиллом c , включала современные Ochrophyta (автотрофные Stramenopiles ), Haptophyta , Cryptophyta , Dinophyta и Choanoflagellida . Euglenophyceae состав были переведены в Chlorophyta .
Хромофита (Буррелли, 1968)
[ редактировать ]В состав Chromophyta (Bourrelly, 1968) вошли современные Ochrophyta (автотрофные Stramenopiles ), Haptophyta и Choanoflagellida . Cryptophyceae (= Dinophyta и Dinophyceae были частью Pyrrophyta ).
Хромиста (Кавальер-Смит, 1981)
[ редактировать ]Название Chromista было впервые введено Кавальер-Смитом в 1981 году; [5] более ранние названия Chromophyta, Chromobiota и Chromobionta соответствуют примерно одной и той же группе. Было описано, что он состоит из трех разных групп: [17] В него входят все простейшие, пластиды которых содержат хлорофилл c . [1]
- Гетероконты или страменопилы: бурые водоросли , диатомеи , водная плесень и т. д.
- Гаптофиты
- Криптомонады
В 1994 году Кавальер-Смит и его коллеги указали, что Chromista, вероятно, представляет собой полифилетическую группу, члены которой возникли независимо, имея не более чем происхождение от общего предка всех эукариот: [1]
Четыре типа, содержащие хромофитные водоросли (с хлорофиллом c, т. е. Cryptista, Heterokonta, Haptophyta, Dinozoa), имеют отдаленное родство друг с другом и с Chlorarachniophyta на наших деревьях. Однако все фотосинтетические таксоны в каждом из этих четырех типов существенно расходятся друг от друга после излучения самих четырех типов. Это подтверждает мнение о том, что общий предок этих четырех типов не был фотосинтезирующим и что хлоропласты были имплантированы отдельно в каждый из них гораздо позже. Эта вероятная полифилия хромофитных водорослей, если она подтвердится, сделает желательным рассматривать Cryptista, Heterokonta и Haptophyta как отдельные царства, а не группировать их в одно царство Chromista. [1]
В 2009 году Кавалер-Смит объяснил причину создания нового королевства, сказав: [7]
Я установил Chromista как царство, отличное от Plantae и Protozoa, на основании доказательств того, что хромистные хлоропласты были приобретены вторично в результате порабощения красной водоросли, которая сама является членом царства Plantae, и их уникальной топологии мембран. [7]
С тех пор Chromista в разное время определялась по-разному. В 2010 году Кавальер-Смит реорганизовал Chromista, включив в нее супергруппу SAR (названную в честь входящих в нее групп Stramenopiles , Alveolata и Rhizaria ) и Hacrobia ( Haptista и Cryptista ). [7]
Патрон и др . (2004) считали наличие уникального класса фермента FBA (фруктозо-1,6-биофосфат-альдолазы), не похожего на тот, который обнаружен у растений, свидетельством монофилии хромистов. [18] Фаст и др . (2001) подтвердили единое происхождение пластид мизозоа (динофлагеллята + апикомплексан), гетероконта и криптофита на основании сравнения генов GAPDH (глицеральдегид-3-фосфатдегидрогеназы). [19] Harper & Keeling (2003) описали гомологи гаптофитов и сочли их еще одним свидетельством единого эндосимбиотического события с участием предка всех хромистов. [20]
Chromalveolata (Adl et al ., 2005)
[ редактировать ]В состав Chromalveolata вошли Stramenopiles , Haptophyta , Cryptophyta и Alveolata . [21] Однако в 2008 году было установлено, что группа не является монофилетической. [22] [23] и более поздние исследования подтвердили это. [24] [25]
Классификация
[ редактировать ]Кавальер-Смит и др . 2015 год
[ редактировать ]В 2015 году Кавальер-Смит и его коллеги создали новую группировку всех организмов более высокого уровня как пересмотр модели семи королевств. В нем они разделили королевство Хромиста на 2 подцарства и 11 типов, а именно: [26]
| Хромиста | |
| 2015 |
Кавалер-Смит 2018
[ редактировать ]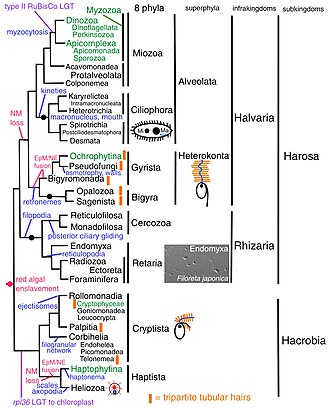
Кавалье-Смит провел новый анализ Chromista в 2018 году, в котором он классифицировал всех хромистов на 8 типов (Gyrista соответствует вышеуказанному типу Ochrophyta и Pseudofungi, Cryptista соответствует вышеуказанному типу Cryptista и «NN», Haptista соответствует вышеуказанному типу Haptophyta). и Гелиозоа): [6]
| Хромиста | |
| 2018 |
Полифилия и серийный эндосимбиоз
[ редактировать ]Молекулярным деревьям было трудно разрешить отношения между различными группами. Все три могут иметь общего предка с альвеолятами (см. хромальвеолаты ), но есть свидетельства того, что гаптофиты и криптомонады не принадлежат вместе с гетероконтами или кладой SAR, но могут быть связаны с Archaeplastida . [4] [27] Криптиста может быть сестрой или частью Археопластиды. [24] хотя это могло быть артефактом, связанным с приобретением криптомонадами генов красных водорослей. [3]
В филогении эукариот 2020 года говорится, что «гипотеза хромальвеолата не получила широкого признания» (отмечая Cavalier-Smith et al 2018). [28] как исключение), объясняя, что в «большинстве филогенетических анализов» линии хозяина не кажутся тесно связанными. [2] [29] Более того, ни одна из групп TSAR, Cryptista и Haptista, ранее входивших в состав Chromalveolata, по-видимому, «вероятно, не была наследственно обусловлена красными вторичными пластидами». Это связано с тем, что многие нефотосинтезирующие организмы относятся к группам с хлорофиллом с , а также с возможностью того, что криптофиты более тесно связаны с растениями. [2]
Альтернативой монофилии является серийный эндосимбиоз , означающий, что «хромисты» получили свои пластиды друг от друга, а не унаследовали их от одного общего предка. Таким образом, филогения характерных пластид, которые, как принято считать, имеют общее происхождение у родофитов, отличается от филогении клеток-хозяев. [3] В 2021 году Юрген Штрассерт и его коллеги смоделировали сроки предполагаемого распространения красных пластид, придя к выводу, что «гипотезы серийного эндосимбиоза хронологически возможны, поскольку стволовые линии всех групп, содержащих красные пластиды, перекрываются во времени» во время мезопротерозоя и Неопротерозойская эра. Они предполагают, что пластиды передавались между группами следующим образом: [3]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и Кавалер-Смит, Томас; Олсопп, Монтана; Чао, Э.Э. (ноябрь 1994 г.). «Химерная загадка: нуклеоморфы и хромисты монофилетичны или полифилетичны?» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (24): 11368–11372. Бибкод : 1994PNAS...9111368C . дои : 10.1073/pnas.91.24.11368 . ПМЦ 45232 . ПМИД 7972066 .
- ^ Перейти обратно: а б с Бурки, Фабьен; Роджер, Эндрю Дж.; Браун, Мэтью В.; Симпсон, Аластер ГБ (2020). «Новое дерево эукариотов» . Тенденции в экологии и эволюции . 35 (1). Эльзевир: 43–55. дои : 10.1016/j.tree.2019.08.008 . ISSN 0169-5347 . ПМИД 31606140 . S2CID 204545629 .
- ^ Перейти обратно: а б с д и Штрассерт, Юрген Ф.Х.; Ирисарри, Икер; Уильямс, Том А.; Бурки, Фабьен (25 марта 2021 г.). «Молекулярная временная шкала эволюции эукариот с последствиями для происхождения пластид, полученных из красных водорослей» . Природные коммуникации . 12 (1): 1879. Бибкод : 2021NatCo..12.1879S . дои : 10.1038/s41467-021-22044-z . ISSN 2041-1723 . ПМЦ 7994803 . ПМИД 33767194 .
- ^ Перейти обратно: а б Парфри, Лаура Вегенер; Барберо, Эрика; Лассер, Элиз; и др. (декабрь 2006 г.). «Оценка поддержки текущей классификации эукариотического разнообразия» . ПЛОС Генетика . 2 (12): е220. дои : 10.1371/journal.pgen.0020220 . ПМЦ 1713255 . ПМИД 17194223 .
- ^ Перейти обратно: а б с Кавальер-Смит, Томас (1981). «Царства эукариот: семь или девять?». Биосистемы . 14 (3–4): 461–81. дои : 10.1016/0303-2647(81)90050-2 . ПМИД 7337818 .
- ^ Перейти обратно: а б Кавальер-Смит, Томас (2018). «Королевство Chromista и его восемь типов: новый синтез, подчеркивающий нацеливание на перипластидные белки, эволюцию цитоскелета и перипластид, а также древние расхождения» . Протоплазма . 255 (1): 297–357. дои : 10.1007/s00709-017-1147-3 . ПМЦ 5756292 . ПМИД 28875267 .
- ^ Перейти обратно: а б с д Кавалер-Смит, Томас (2009). «Царства Protozoa и Chromista и эозойный корень эукариотического дерева» . Письма по биологии . 6 (3): 342–345. дои : 10.1098/rsbl.2009.0948 . ПМК 2880060 . ПМИД 20031978 .
- ^ Килинг, Патрик Дж. (2009). «Хромальвеолаты и эволюция пластид путем вторичного эндосимбиоза». Журнал эукариотической микробиологии . 56 (1): 1–8. дои : 10.1111/j.1550-7408.2008.00371.x . ПМИД 19335769 . S2CID 34259721 .
- ^ Понсе-Толедо, Рафаэль И.; Дешам, Филипп; Лопес-Гарсия, «Очищение»; Живанович, Иван; Бензерара, Карим; Морейра, Дэвид (2017). «Ранневетвящиеся пресноводные цианобактерии у происхождения пластид» . Современная биология . 27 (3): 386–391. дои : 10.1016/j.cub.2016.11.056 . ПМК 5650054 . ПМИД 28132810 .
- ^ Килинг, Патрик Дж. (2010). «Эндсимбиотическое происхождение, диверсификация и судьба пластид» . Философские труды Королевского общества B: Биологические науки . 365 (1541): 729–748. дои : 10.1098/rstb.2009.0103 . ПМК 2817223 . ПМИД 20124341 .
- ^ Морозов А.А.; Галачанц, Юрий П. (2019). «Гены диатомовых водорослей, происходящие из красных и зеленых водорослей: значение для моделей вторичного эндосимбиоза». Морская геномика . 45 : 72–78. Бибкод : 2019Март..45...72М . дои : 10.1016/j.margen.2019.02.003 . ПМИД 30792089 . S2CID 73458340 .
- ^ Райс, Дэнни В.; Палмер, Джеффри Д. (2006). «Исключительный горизонтальный перенос генов в пластидах: замена гена отдаленным бактериальным паралогом и свидетельство того, что гаптофитные и криптофитные пластиды являются сестрами» . БМК Биология . 4 (1): 31. дои : 10.1186/1741-7007-4-31 . ПМК 1570145 . ПМИД 16956407 .
- ^ де Ревье, Бруно (2006). Биология и филогения водорослей . Порту-Алегри: Редакция Artmed. стр. 156–157. ISBN 9788536315102 .
- ^ Блэквелл, Уилл Х. (2009). «Возвращение к Хромисте: дилемма перекрытия предполагаемых королевств и попытка применения ботанического номенклатурного кода» (PDF) . Фитология . 91 (2): 191–225.
- ^ Шадефо, Мариус (1950). «Плавающие клетки морских водорослей ветви Chromophyceae». Еженедельные отчеты сессий Академии наук (на французском языке). 231 : 788–790.
- ^ Шадефо, Мариус (1960). «Бессосудистые растения (криптогамия)». В Шадефо, Мариус; Эмбергер, Л. (ред.). Трактат по систематической ботанике . Париж: Том I.
- ^ Чурос, М.; Рогозин И.Б.; Кунин, Евгений В. (май 2008 г.). «Гены, богатые интронами у альвеолятных предков, выявленные с помощью гибкого подхода максимального правдоподобия» . Молекулярная биология и эволюция . 25 (5): 903–911. дои : 10.1093/molbev/msn039 . ПМИД 18296415 .
- ^ Покровитель Никола Дж.; Роджерс, Мэтью Б.; Килинг, Патрик Дж. (2004). «Замена гена фруктозо-1,6-бисфосфатальдолазы подтверждает гипотезу о единственном фотосинтетическом предке хромальвеолатов» . Эукариотическая клетка . 3 (5): 1169–1175. doi : 10.1128/EC.3.5.1169-1175.2004 . ISSN 1535-9778 . ПМК 522617 . ПМИД 15470245 .
- ^ Фаст, Наоми М.; Киссинджер, Джессика С.; Роос, Дэвид С.; Килинг, Патрик Дж. (01 марта 2001 г.). «Ядерно-кодируемые гены, нацеленные на пластиды, предполагают единое общее происхождение пластид апикомплексана и динофлагеллят» . Молекулярная биология и эволюция . 18 (3): 418–426. doi : 10.1093/oxfordjournals.molbev.a003818 . ISSN 1537-1719 . ПМИД 11230543 .
- ^ Харпер, Джей Ти (27 июня 2003 г.). «Кодируемая в ядре глицеральдегид-3-фосфатдегидрогеназа (GAPDH), нацеленная на пластиды, указывает на единое происхождение хромальвеолатных пластид» . Молекулярная биология и эволюция . 20 (10): 1730–1735. дои : 10.1093/molbev/msg195 . ISSN 0737-4038 . ПМИД 12885964 .
- ^ Адл, Сина М.; и др. (2005). «Новая классификация эукариот более высокого уровня с акцентом на таксономию протистов» . Журнал эукариотической микробиологии . 52 (5): 399–451. дои : 10.1111/j.1550-7408.2005.00053.x . ПМИД 16248873 . S2CID 8060916 .
- ^ Бурки, Фабьен; Шалчиан-Тебризи, Камран и Павловский, Январь (2008). «Филогеномика открывает новую «мегагруппу», включающую большинство фотосинтезирующих эукариот» . Письма по биологии . 4 (4): 366–369. дои : 10.1098/rsbl.2008.0224 . ПМК 2610160 . ПМИД 18522922 .
- ^ Ким, Э.; Грэм, Луизиана (июль 2008 г.). Редфилд, Розмари Джин (ред.). «Анализ EEF2 бросает вызов монофилии Archaeplastida и Chromalveolata» . ПЛОС ОДИН . 3 (7): e2621. Бибкод : 2008PLoSO...3.2621K . дои : 10.1371/journal.pone.0002621 . ПМК 2440802 . ПМИД 18612431 .
- ^ Перейти обратно: а б Бурки, Фабьен; Каплан, Майя; Тихоненков Денис Владимирович; и др. (январь 2016 г.). «Распутывание ранней диверсификации эукариот: филогеномное исследование эволюционного происхождения Centrohelida, Haptophyta и Cryptista» . Слушания. Биологические науки . 283 (1823): 20152802. doi : 10.1098/rspb.2015.2802 . ПМК 4795036 . ПМИД 26817772 .
- ^ Бурки, Ф.; Окамото, Н.; Помберт, Дж. Ф.; Килинг, ПиДжей (2012). «Эволюционная история гаптофитов и криптофитов: филогеномные доказательства раздельного происхождения» . Труды Королевского общества . 279 (1736): 2246–2254. дои : 10.1098/rspb.2011.2301 . ПМК 3321700 . ПМИД 22298847 .
- ^ Руджеро, Майкл А.; Гордон, Деннис П.; Оррелл, Томас М.; и др. (2015). «Классификация всех живых организмов более высокого уровня» . ПЛОС ОДИН . 10 (4): e0119248. Бибкод : 2015PLoSO..1019248R . дои : 10.1371/journal.pone.0119248 . ПМЦ 4418965 . ПМИД 25923521 .
- ^ Бурки, Фабьен; Шалчиан-Тебризи, Кямран; Минге, Марианна; и др. (август 2007 г.). «Филогеномика меняет супергруппы эукариот» . ПЛОС ОДИН 2 (8): е790. Бибкод : 2007PLoSO...2..790B . дои : 10.1371/journal.pone.0000790 . ЧВК 1949142 . ПМИД 17726520 .
- ^ Кавальер-Смит, Томас ; Чао, Эма Э.; Льюис, Родри (17 апреля 2018 г.). «Мультигенная филогения и клеточная эволюция хромистского инфрацарства Rhizaria: контрастная клеточная организация сестринских типов Cercozoa и Retaria» . Протоплазма . 255 (5): 1517–1574. дои : 10.1007/s00709-018-1241-1 . ISSN 0033-183X . ПМК 6133090 . ПМИД 29666938 .
- ^ Бурки, Ф. (2017). «Запутанная эволюция эукариот со сложными пластидами». Ин Хиракава, Ю. (ред.). Достижения в ботанических исследованиях . Том. 84. Академическая пресса. стр. 1–30.
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|