Анатомия гиппокампа


Анатомия гиппокампа физические аспекты и свойства гиппокампа , нервной структуры в медиальной височной доле мозга описывает . Он имеет характерную изогнутую форму, которую сравнивают с морским коньком из греческой мифологии и бараньими рогами Амона из египетской мифологии . Эта общая схема применима ко всему спектру видов млекопитающих , от ежа до человека, хотя детали различаются. Например, у крысы два гиппокампа похожи на пару бананов, соединенных стеблями. В мозгу приматов , включая человека, часть гиппокампа у основания височной доли намного шире, чем часть наверху. Из-за трехмерной кривизны этой конструкции обычно можно увидеть двумерные сечения, подобные показанным. Нейровизуализационные снимки могут показывать различные формы, в зависимости от угла и местоположения разреза.
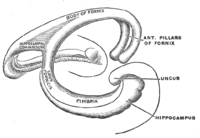
Топологически поверхность полушария головного мозга можно рассматривать как сферу с углублением в месте прикрепления к среднему мозгу. Структуры, выстилающие край отверстия, в совокупности составляют так называемую систему (лат лимбическую . border ), причем гиппокамп выстилает задний край этого отверстия. Эти лимбические структуры включают гиппокамп, поясную извилину , обонятельную кору и миндалевидное тело . Пол Маклин однажды в рамках своей теории тройственного мозга предположил , что лимбические структуры составляют нервную основу эмоций . Хотя большинство нейробиологов больше не верят в концепцию единой «лимбической системы», эти регионы тесно взаимосвязаны и взаимодействуют друг с другом. [ нужна ссылка ]
Базовая схема гиппокампа
[ редактировать ]
Начиная с зубчатой извилины и двигаясь внутрь вдоль S-образной кривой гиппокампа, необходимо пройти черезряд узких зон. Первая из них, зубчатая извилина (ЗГ), фактически представляет собой отдельнуюСтруктура — плотно упакованный слой мелких гранулярных клеток, обернутый вокруг конца собственно гиппокампа , образующий в одних поперечных сечениях заостренный клин, в других — полукруг. Следующийследует ряд областей Cornu Ammonis : сначала СА4 (который лежит в основе зубчатой извилины), затем СА3 , затемочень маленькая зона называется CA2 , затем CA1 . Все области СА заполнены плотно упакованными пирамидальными клетками, подобными тем, которые обнаружены в неокортексе . После CA1 идет область, называемая субикулюмом . После этого идет пара нечетких областей, называемых пресубикулум и парасубикулум, а затемпереход в собственно кору (преимущественно энторинальную область коры). Большинство анатомовиспользуйте термин «собственно гиппокамп» для обозначения четырех полей СА и формирования гиппокампа. для обозначения собственно гиппокампа плюс зубчатой извилины и субикулюма. [1]
Основные сигнальные пути проходят через гиппокамп и объединяются, образуя петлю. Большая часть внешней информации поступает из прилегающей энторинальной коры через аксоны так называемого перфорантного пути . Эти аксоны возникают из слоя 2 энторинальной коры (EC) и заканчиваются в зубчатой извилине и CA3. Существует также отдельный путь от слоя 3 EC непосредственно к CA1, часто называемый темпороаммонным или TA-CA1 путем. Гранулярные клетки DG посылают свои аксоны (называемые «мшистыми волокнами») к CA3. Пирамидные клетки СА3 посылают свои аксоны в СА1. Пирамидные клетки СА1 посылают свои аксоны в субикулюм и глубокие слои ЭК. Субикулярные нейроны направляют свои аксоны преимущественно в ЭК. Перфорантный путь от зубчатой извилины до CA3-CA1 Пер Андерсен назвал трисинаптической цепью , которая отметила, что из гиппокампа можно вырезать тонкие срезы перпендикулярно его длинной оси таким образом, чтобы сохранить все эти связи. Это наблюдение легло в основу его пластинчатой гипотезы. , который предположил, что гиппокамп можно рассматривать как серию параллельных полосок, действующих функционально независимым образом. [2] Ламеллярную концепцию до сих пор иногда считают полезным организующим принципом, но более поздние данные, показывающие обширные продольные связи внутри гиппокампальной системы, потребовали ее существенной модификации. [3]
Входной сигнал от перфорантного пути от слоя II EC поступает в зубчатую извилину и передается в область CA3 (и в мшистые клетки, расположенные в воротах зубчатой извилины, которые затем отправляют информацию в отдаленные части зубчатой извилины, где цикл повторяется). Регион CA3 объединяет этот входной сигнал с сигналами от слоя II EC и отправляет обширные соединения внутри региона, а также отправляет соединения в лучистые и ориентальные слои ипсилатеральных и контралатеральных регионов CA1 через набор волокон, называемых коллатералями Шаффера и комиссуральными волокнами.путь соответственно. [4] [5] [6] Область CA1 получает входные данные от подполя CA3, слоя III EC и восходящего ядра таламуса (которые проецируются только на терминальные апикальные дендритные пучки в молекулярном слое lacunosum ). В свою очередь, CA1 проецируется на субикулюм, а также отправляет информацию по вышеупомянутым выходным путям гиппокампа. Субикулум является заключительной стадией пути, объединяющей информацию из проекции CA1 и слоя III EC, чтобы также отправлять информацию по выходным путям гиппокампа.
Гиппокамп также получает ряд подкорковых сигналов. У Macaca fascicleis эти входы включают миндалевидное тело (в частности, переднюю миндалевидную область, базолатеральное ядро и периамигдалоидную кору), медиальную перегородку и диагональную полоску Брока , клауструм , безымянную субстанцию и базальное ядро Мейнерта , таламус (включая передний ядерный комплекс, латеродорсальное ядро, паравентрикулярное и паратениальное ядра, воссоединяющее ядро и центральное медиальное ядро), латеральные преоптические и латеральные гипоталамические области, супрамаммилярную и ретромамилярную области, вентральную покрышку , покрышку ретикулярные поля, ядра шва (верхнее центральное ядро и дорсальное ядро шва), ретикулярное ядро tegementi pontis , периакведуктальное серое , дорсальное тегментальное ядро и голубое пятно .Гиппокамп также получает прямые моносинаптические проекции от фастигиального ядра мозжечка . [7]
Основные системы волокон у крысы
[ редактировать ]Угловая связка
[ редактировать ]Эти волокна начинаются от вентральной части энторинальной коры (ЭК) и содержат комиссуральный (ЭК◀▶Гиппокамп) и перфорантный пути (возбуждающий ЭК▶СА1 и тормозной ЭК◀▶СА2). [8] ) волокна. Они путешествуют вдоль септовисочной оси гиппокампа. Волокна перфорантного пути, как следует из названия, перфорируют субикулюм, прежде чем попасть в гиппокамп (поля СА) и зубчатую извилину. [9]
Путь фимбрия-свод
[ редактировать ]
Волокна бахромчатого свода являются гиппокампальными и субикулярными воротами в области мозга и обратно подкорковые . [10] [11] Разные части этой системы имеют разные названия:
- Белые миелиновые волокна, покрывающие желудочковые (глубинные) части гиппокампа, образуют альвеусы .
- Волокна, покрывающие височные отделы гиппокампа, образуют пучок волокон, называемый фимбриями . Переходя от височной к перегородочной (дорсальной) части бахромок гиппокампа, собирается все больше гиппокампальных и субикулярных выходов и утолщается.
- По средней линии и под мозолистым телом эти волокна образуют свод .
На уровне контура альвея содержит аксональные волокна от DG и пирамидных нейронов СА3, СА2, СА1 и субикулюма ( СА1 ▶ субикулюм и СА1 ▶ энторинальные проекции), которые собираются в височном гиппокампе, образуя фимбрии/свод, один из Основные выходы гиппокампа. [12] [13] [14] [15] [16] У крысы некоторые медиальные и латеральные энторинальные аксоны ( энторинальные ▶ проекции CA1) проходят через альвеусы по направлению к молекулярному слою CA1, не создавая значительного количества проходных бутонов на других слоях CA1 ( темпороаммонический альвеарный путь ). [13] [17] Контралатеральные энторинальные ▶ выступы CA1 почти всегда проходят через альвеус. Чем больше перегородки, тем больше ипсилатеральных проекций энторинального СА1, которые идут по альвеарному пути (вместо перфорантного пути). [18] Хотя субикулюм посылает аксональные проекции в альвеус, субикулюм ▶ Проекция СА1 проходит через восточные и молекулярные слои субикулюма и СА1. [19] Холинергические и ГАМКергические проекции от MS-DBB к CA1 также проходят через фимбрии. [20] Стимуляция фимбрии приводит к холинергическому возбуждению клеток CA1 O-LMR . [21]
Известно также, что внеклеточная стимуляция фимбрии стимулирует пирамидные клетки CA3 антидромно и ортодромно, но не оказывает влияния на зубчатые гранулярные клетки. [22] Каждая пирамидальная клетка CA1 также посылает аксональную ветвь к фимбриям. [23] [24]
Спайки гиппокампа
[ редактировать ]Хилярные мшистые клетки и пирамидные клетки CA3 являются основным источником комиссуральных волокон гиппокампа . Они проходят через спайки гиппокампа и достигают контралатеральных областей гиппокампа. Комиссуры гиппокампа имеют дорсальный и вентральный сегменты. Дорсальные комиссуральные волокна состоят в основном из энторинальных и пресубикулярных волокон, идущих к гиппокампу и зубчатой извилине или от них. [9] Как правило, можно сказать, что каждое цитоархитектоническое поле, которое вносит вклад в комиссуральную проекцию, также имеет параллельное ассоциативное волокно, которое заканчивается в ипсилатеральном гиппокампе. [25] Внутренний молекулярный слой зубчатой извилины (дендриты как гранулярных клеток, так и ГАМКергических интернейронов) получает выступ, имеющий как ассоциативные, так и комиссуральные волокна преимущественно из прикорневых мшистых клеток и в некоторой степени из пирамидных клеток СА3с. Поскольку эти проекционные волокна берут начало как с ипсилатеральной, так и с контралатеральной стороны гиппокампа, их называют ассоциативными/комиссуральными проекциями. Фактически каждая мшистая клетка иннервирует как ипсилатеральную, так и контрлатеральную зубчатую извилину. Хорошо известная трисинаптическая цепь гиппокампа простирается преимущественно горизонтально вдоль гиппокампа. Однако ассоциативные/комиссуральные волокна, такие как ассоциативные проекции пирамидальных клеток CA2, простираются в основном продольно (дорсовентрально) вдоль гиппокампа. [26] [27] Комиссуральные волокна, происходящие из пирамидных клеток СА3, идут в области СА3, СА2 и СА1. Подобно мшистым клеткам, одна пирамидальная клетка CA3 вносит вклад как в комиссуральные, так и в ассоциативные волокна, и они заканчиваются как на главных клетках, так и на интернейронах. [28] [29] Слабый комиссуральный выступ соединяет обе области CA1 вместе. Субикулум не имеет комиссуральных входов и выходов. По сравнению с грызунами комиссуральные связи гиппокампа у обезьян и человека гораздо менее распространены. [30] Хотя возбуждающие клетки вносят основной вклад в комиссуральные пути, среди их окончаний обнаружен ГАМКергический компонент, происхождение которого было прослежено до ворот. [31] Стимуляция комиссуральных волокон антидромно стимулирует DG внутригрудной перфорантный путь (HIPP) и триламинарные клетки CA3. [32]
Клетки и слои гиппокампа
[ редактировать ]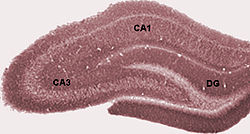

Собственно гиппокамп
[ редактировать ]состоит Собственно гиппокамп из ряда подполей. Хотя терминология у разных авторов различается, наиболее часто используемые термины — это зубчатая извилина и аммониевый рог (буквально « CA рог Аммона», сокращенно ) . Зубчатая извилина содержит зубчатую фасцию и ворота , тогда как СА дифференцируется на подполя СА1, СА2, СА3 и СА4 .
Однако область, известная как CA4, на самом деле представляет собой «глубокий полиморфный слой зубчатой извилины». [33] (как пояснил Теодор Блэкстад (1956) [34] и Дэвид Амарал (1978)). [35]
В поперечном разрезе гиппокамп представляет собой С-образную структуру, напоминающую бараньи рога . Название cornu ammonis относится к египетскому божеству Амону , имеющему голову барана. Рогатый вид гиппокампа вызван разницей плотности клеток и различной степенью расположения нейрональных волокон .
У грызунов гиппокамп расположен так, что примерно один его конец находится вблизи верхней части головы (дорсальный или перегородочный конец), а другой конец — вблизи нижней части головы (вентральный или височный конец). Как показано на рисунке, сама структура изогнута, а вдоль кривой определены подполя или области, от СА4 до СА1 (помечены только СА3 и СА1). Регионы СА также структурированы по глубине в четко выраженные пласты (или слои):
- Stratum oriens (str. oriens) — следующий поверхностный к альвеусу слой. клеточные тела тормозных корзиночных клеток В этом слое расположены базальные дендриты и горизонтальных триламинарных клеток, названных в честь их аксонов, иннервирующих три слоя — восточный, пирамидальный и лучистый. Здесь также находятся пирамидных нейронов, где они получают входные сигналы от других пирамидных клеток, септальных волокон и комиссуральных волокон из контралатерального гиппокампа (обычно рекуррентные соединения, особенно в СА3 и СА2). У грызунов два гиппокампа тесно связаны между собой, но у приматов эта комиссуральная связь гораздо реже.
- Пирамидный слой (str.pyr.) содержит тела пирамидальных нейронов, которые являются основными возбуждающими нейронами гиппокампа. Этот слой, как правило, является одним из наиболее видимых невооруженным глазом слоев. В области CA3 этот слой содержит синапсы мшистых волокон, проходящих через блестящий слой. Этот слой также содержит клеточные тела многих интернейронов , включая аксо-аксонические клетки, бистратифицированные клетки и радиальные триламинарные клетки.
- Stratum lucidum (str. luc.) — один из самых тонких слоев гиппокампа, обнаруженный только в области CA3. зубчатой извилины Мшистые волокна из гранулярных клеток проходят через этот слой в CA3, хотя синапсы этих волокон можно обнаружить в str. пир.
- Stratum radiatum (str. rad.), как и str. oriens, содержит перегородочные и комиссуральные волокна. Он также содержит коллатеральные волокна Шаффера , которые являются проекциями вперед от СА3 к СА1. Здесь также можно найти некоторые интернейроны, которые можно найти в более поверхностных слоях, включая корзиночные клетки, бистратифицированные клетки и радиальные триламинарные клетки.
- Stratum lacunosum (str. lac.) — тонкий слой, который также содержит коллатеральные волокна Шаффера, но также содержит волокна перфорантного пути из поверхностных слоев энторинальной коры. Из-за своего небольшого размера его часто группируют вместе с молекулярным слоем в единый слой, называемый stratum lacunosum-moleculare (str. lm.).
- Молекулярный слой (str. мол.) — самый поверхностный слой гиппокампа. Здесь волокна перфорантного пути образуют синапсы на дистальных апикальных дендритах пирамидных клеток.
- Борозда гиппокампа (sulc.) или щель — бесклеточная область, отделяющая поле СА1 от зубчатой извилины. Поскольку фаза регистрируемого тета-ритма систематически варьируется в разных слоях, борозду часто используют в качестве фиксированной контрольной точки для регистрации ЭЭГ , поскольку ее легко идентифицировать. [33]
Зубчатая извилина
[ редактировать ]состоит Зубчатая извилина из аналогичного ряда слоев:
- Полиморфный слой (poly.lay.) является наиболее поверхностным слоем зубчатой извилины и часто рассматривается как отдельное подполе (как ворота). Этот слой содержит множество интернейронов , и аксоны зубчатых гранулярных клеток проходят через этот слой на пути к CA3.
- Stratum granulosum (str. gr.) содержит клеточные тела зубчатых клеток-зерен.
- Молекулярный слой, внутренняя треть (str.mol.1/3), — это то место, где проходят оба комиссуральных волокна контралатеральной зубчатой извилины и образуют синапсы, а также место, где заканчиваются входы от медиальной перегородки , как на проксимальных дендритах гранулярных клеток.
- Молекулярный слой, внешние две трети (молекулярный слой 2/3), является самым глубоким из слоев, расположен поверхностным слоем от борозды гиппокампа напротив молекулярного слоя в полях СА. Волокна перфорантного пути проходят через эти слои, образуя возбуждающие синапсы на дистальных апикальных дендритах гранулярных клеток.
поддерживается актуальная база знаний о типах нейронов формирования гиппокампа, их профиле биомаркеров, активных и пассивных электрофизиологических параметрах и связях На веб-сайте «Гиппокамп» . [36]
Ссылки
[ редактировать ]- ^ Амарал, Д; Лавенекс П (2006). «Глава 3. Нейроанатомия гиппокампа». В Андерсене П; Моррис Р.; Амарал Д; Блисс Т; О'Киф Дж. (ред.). Книга «Гиппокамп» . Издательство Оксфордского университета. ISBN 978-0-19-510027-3 .
- ^ Андерсен, П; Блисс ТВП; Скреде К.К. (1971). «Ламеллярная организация возбуждающих путей гиппокампа». Эксп. Мозговой Рес . 13 (2): 222–238. дои : 10.1007/BF00234087 . ПМИД 5570425 . S2CID 12075886 .
- ^ Андерсен, П; Соленг А.Ф.; Раастад М (2000). «Пересмотр гипотезы пластинок гиппокампа». Мозговой Рес . 886 (1–2): 165–171. дои : 10.1016/S0006-8993(00)02991-7 . ПМИД 11119694 . S2CID 8455285 .
- ^ Йорт-Симонсен, А. (15 января 1973 г.). «Некоторые внутренние связи гиппокампа у крысы: экспериментальный анализ». Журнал сравнительной неврологии . 147 (2): 145–61. дои : 10.1002/cne.901470202 . ПМИД 4118866 . S2CID 28989051 .
- ^ Суонсон, LW; Висс, Дж. М.; Коуэн, WM (15 октября 1978 г.). «Ауторадиографическое исследование организации внутригиппокампальных ассоциативных путей у крыс». Журнал сравнительной неврологии . 181 (4): 681–715. дои : 10.1002/cne.901810402 . ПМИД 690280 . S2CID 30954240 .
- ^ Лаурберг, С. (15 апреля 1979 г.). «Комиссуральные и внутренние связи гиппокампа крысы». Журнал сравнительной неврологии . 184 (4): 685–708. дои : 10.1002/cne.901840405 . ПМИД 422759 . S2CID 27256712 .
- ^ Хит Р.Г., Харпер Дж.В. (ноябрь 1974 г.). «Восходящие проекции фастигиального ядра мозжечка в гиппокамп, миндалевидное тело и другие участки височной доли: вызванные потенциальные и гистологические исследования на обезьянах и кошках». Эксп. Нейрол . 45 (2): 268–87. дои : 10.1016/0014-4886(74)90118-6 . ПМИД 4422320 .
- ^ Мельцер, С.; Майкл, М.; Капути, А.; Элиава, М.; Фукс, ЕС; Уиттингтон, Массачусетс; Моньер, Х. (22 марта 2012 г.). «Дальнопроецирующие ГАМКергические нейроны модулируют торможение в гиппокампе и энторинальной коре» . Наука . 335 (6075): 1506–1510. Бибкод : 2012Sci...335.1506M . дои : 10.1126/science.1217139 . ПМИД 22442486 . S2CID 206539012 .
- ^ Jump up to: а б Андерсен, Пер; и др., ред. (2007). Книга «Гиппокамп . Нью-Йорк: Издательство Оксфордского университета. п. 47,63,123. ISBN 9780195100273 .
- ^ ПАУЭЛЛ, ТП; ГИЛЛЕРИ, RW; КОУЭН, WM (октябрь 1957 г.). «Количественное исследование свода мамилло-таламической системы» . Журнал анатомии . 91 (4): 419–37. ПМЦ 1244899 . ПМИД 13475143 .
- ^ ДАЙЦ, ХМ; ПАУЭЛЛ, Т.П. (февраль 1954 г.). «Исследование связей сводовой системы» . Журнал неврологии, нейрохирургии и психиатрии . 17 (1): 75–82. дои : 10.1136/jnnp.17.1.75 . ПМК 503161 . ПМИД 13131081 .
- ^ Ноулз, штат Вашингтон; Шварцкройн, Пенсильвания (ноябрь 1981 г.). «Аксональные разветвления пирамидальных клеток Ca1 гиппокампа» . Журнал неврологии . 1 (11): 1236–41. doi : 10.1523/JNEUROSCI.01-11-01236.1981 . ПМК 6564220 . ПМИД 6171629 .
- ^ Jump up to: а б Книга «Гиппокамп . Нью-Йорк: Издательство Оксфордского университета. 2007. с. 47. ИСБН 9780199723164 .
- ^ Аллоуэй, Томас К. Притчард, Кевин Д. (1999). Медицинская нейробиология (1-е изд.). Мэдисон, Коннектикут: Паб Fence Creek. п. 28. ISBN 978-1889325293 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Годрон, Анри М. Дювернуа, Франсуаза Каттен, Пьер-Ив Рисольд; рисунки и иллюстрации Дж. Л. Ваннсона и М. (2013). Функциональная анатомия гиппокампа человека, васкуляризация и серийные срезы с помощью МРТ (4-е изд.). Берлин: Шпрингер. п. 28. ISBN 978-3-642-33603-4 .
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тамамаки, Н; Абэ, К; Нодзё, Ю. (14 июня 1988 г.). «Трехмерный анализ всех аксональных ветвей, происходящих от одиночных пирамидальных нейронов CA2 в гиппокампе крысы, с помощью метода компьютерной графики». Исследования мозга . 452 (1–2): 255–72. дои : 10.1016/0006-8993(88)90030-3 . ПМИД 3401733 . S2CID 25038544 .
- ^ Шетти, АК (2002). «Энторинальные аксоны демонстрируют прорастание в подполе CA1 гиппокампа взрослого человека на крысиной модели височной эпилепсии». Гиппокамп . 12 (4): 534–42. дои : 10.1002/hipo.10031 . ПМИД 12201638 . S2CID 24965222 .
- ^ Деллер, Т; Адельманн, Г; Нитч, Р; Фротчер, М. (декабрь 1996 г.). «Альвеарный путь гиппокампа крысы». Исследования клеток и тканей . 286 (3): 293–303. дои : 10.1007/s004410050699 . ПМИД 8929332 . S2CID 36438302 .
- ^ Харрис, Э; Стюарт, М. (23 марта 2001 г.). «Распространение синхронных эпилептиформных событий из субикулюма назад в область CA1 срезов мозга крысы». Исследования мозга . 895 (1–2): 41–9. дои : 10.1016/s0006-8993(01)02023-6 . ПМИД 11259758 . S2CID 23300272 .
- ^ Гуляш, А.И.; Гёркс, Т.Дж.; Фройнд, Т.Ф. (1990). «Иннервация различных пептидсодержащих нейронов в гиппокампе ГАМКергическими перегородочными афферентами». Нейронаука . 37 (1): 31–44. дои : 10.1016/0306-4522(90)90189-б . ПМИД 1978740 . S2CID 24486668 .
- ^ Леан, Р.Н.; Микулович, С; Леан, Кентукки; Мунгуба, Х; Гезелиус, Х; Энджин, А; Патра, К; Эрикссон, А; Лоу, LM; Торт, АБ; Кулландер, К. (ноябрь 2012 г.). «Интернейроны OLM дифференциально модулируют CA3 и энторинальные входы к нейронам CA1 гиппокампа» . Природная неврология . 15 (11): 1524–30. дои : 10.1038/nn.3235 . ПМЦ 3483451 . ПМИД 23042082 .
- ^ Шарфман, HE (25 июня 1993 г.). «Активация зубчатых внутригрудных нейронов путем стимуляции фимбрии в срезах гиппокампа крысы» . Письма по неврологии . 156 (1–2): 61–6. дои : 10.1016/0304-3940(93)90440-в . ПМК 3281807 . ПМИД 8105429 .
- ^ Ян, Сунгу; Ян, Сунгчил; Морейра, Тайланд; Хоффман, Глория; Карлсон, Грег К.; Бендер, Кевин Дж.; Алджер, Брэдли Э.; Тан, Ча-Мин (2 сентября 2014 г.). «Межламеллярная сеть CA1 в гиппокампе» . Труды Национальной академии наук . 111 (35): 12919–12924. Бибкод : 2014PNAS..11112919Y . дои : 10.1073/pnas.1405468111 . ISSN 0027-8424 . ПМК 4156755 . ПМИД 25139992 .
- ^ Хансакер, Майкл Р.; Кеснер, Раймонд П. (01 января 2013 г.). «Операция разделения шаблонов и процессы завершения шаблонов, связанные с различными атрибутами или областями памяти». Неврологические и биоповеденческие обзоры . 37 (1): 36–58. doi : 10.1016/j.neubiorev.2012.09.014 . ПМИД 23043857 . S2CID 22493885 .
- ^ Суонсон, LW; Висс, Дж. М.; Коуэн, WM (15 октября 1978 г.). «Ауторадиографическое исследование организации внутригиппокампальных ассоциативных путей у крыс». Журнал сравнительной неврологии . 181 (4): 681–715. дои : 10.1002/cne.901810402 . ПМИД 690280 . S2CID 30954240 .
- ^ Амарал, генеральный директор; Виттер, член парламента (1989). «Трехмерная организация образования гиппокампа: обзор анатомических данных». Нейронаука . 31 (3): 571–91. дои : 10.1016/0306-4522(89)90424-7 . ПМИД 2687721 . S2CID 28430607 .
- ^ Кохара, К; Пиньятелли, М; Ривест, Эй Джей; Юнг, HY; Китамура, Т; Су, Джей; Фрэнк, Д; Кадзикава, К; Мизе, Н; Обата, Ю; Уикершем, ИК; Тонегава, С. (февраль 2014 г.). «Генетические и оптогенетические инструменты, специфичные для типа клеток, выявляют цепи CA2 в гиппокампе» (PDF) . Природная неврология . 17 (2): 269–79. дои : 10.1038/nn.3614 . ПМК 4004172 . ПМИД 24336151 .
- ^ Блэкстад, ТВ (октябрь 1956 г.). «Комиссуральные соединения области гиппокампа у крысы с особым упором на способ их завершения». Журнал сравнительной неврологии . 105 (3): 417–537. дои : 10.1002/cne.901050305 . ПМИД 13385382 . S2CID 41672064 .
- ^ Фрике, Р; Коуэн, WM (15 сентября 1978 г.). «Авторадиографическое исследование комиссуральных и ипсилатеральных гиппокампо-зубчатых проекций у взрослой крысы». Журнал сравнительной неврологии . 181 (2): 253–69. дои : 10.1002/cne.901810204 . ПМИД 567658 . S2CID 46320248 .
- ^ Амарал, генеральный директор; Шарфман, HE; Лавенекс, П. (2007). «Зубчатая извилина: фундаментальная нейроанатомическая организация (Зубчатая извилина для манекенов)». Зубчатая извилина: полное руководство по структуре, функциям и клиническим последствиям . Прогресс в исследованиях мозга. Том. 163. стр. 3–22. дои : 10.1016/S0079-6123(07)63001-5 . ISBN 9780444530158 . ПМЦ 2492885 . ПМИД 17765709 .
- ^ Рибак, CE; Сересс, Л; Петерсон, генеральный директор; Сереги, КБ; Фэллон, Дж. Х.; Шмуед, LC (декабрь 1986 г.). «ГАМКергический ингибирующий компонент в комиссуральном пути гиппокампа» . Журнал неврологии . 6 (12): 3492–8. doi : 10.1523/JNEUROSCI.06-12-03492.1986 . ПМК 6568657 . ПМИД 2432200 .
- ^ Сик, Аттила; Пенттонен, Маркку; Бужаки, Дьёрдь (март 1997 г.). «Интернейроны в зубчатой извилине гиппокампа: внутриклеточное исследование in vivo». Европейский журнал неврологии . 9 (3): 573–588. дои : 10.1111/j.1460-9568.1997.tb01634.x . ПМИД 9104599 . S2CID 25960013 .
- ^ Jump up to: а б Андерсен, Пер; и др. (2007). Книга «Гиппокамп» . Издательство Оксфордского университета.
- ^ Блэкстад, ТВ (1956). «Комиссуральные соединения области гиппокампа у крысы, с особым вниманием к способу их завершения». Джей Комп Нейрол . 105 (3): 417–537. дои : 10.1002/cne.901050305 . ПМИД 13385382 . S2CID 41672064 .
- ^ Амарал, генеральный директор (1978). «Исследование типов клеток по методу Гольджи в прикорневой области гиппокампа крысы». Джей Комп Нейрол . 182 (5): 851–914. дои : 10.1002/cne.901820508 . ПМИД 730852 . S2CID 44257239 .
- ^ «Гиппокамп» . сайт hippocampame.org .
Внешние ссылки
[ редактировать ]- Схематическая диаграмма среза мозга гиппокампа
- Гиппокамп Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)
- Окрашенные изображения срезов мозга, включающие «гиппокамп», в проекте BrainMaps.
- Анатомия гиппокампа и его связь