Несинаптическая пластичность

Несинаптическая пластичность — это форма нейропластичности , которая включает модификацию функции ионных каналов в аксоне , дендритах и теле клетки, что приводит к специфическим изменениям в интеграции возбуждающих постсинаптических потенциалов и тормозных постсинаптических потенциалов . Несинаптическая пластичность — это модификация внутренней возбудимости нейрона. Он взаимодействует с синаптической пластичностью , но считается отдельной сущностью от синаптической пластичности. Внутренняя модификация электрических свойств нейронов играет роль во многих аспектах пластичности, от гомеостатической пластичности до обучения и самой памяти . Несинаптическая пластичность влияет на синаптическую интеграцию , подпороговое распространение , генерацию спайков и другие фундаментальные механизмы нейронов на клеточном уровне. Эти отдельные нейрональные изменения могут привести к изменениям в высших функциях мозга, особенно в обучении и памяти. является новой областью Однако, поскольку нейробиология , большая часть знаний о несинаптической пластичности остается неопределенной и все еще требует дальнейшего изучения, чтобы лучше определить ее роль в функционировании и поведении мозга.
Против. синаптическая пластичность
[ редактировать ]Нейропластичность — это способность определенной части или региона нейрона изменять силу с течением времени. Существуют две общепризнанные категории пластичности: синаптическая и несинаптическая. Синаптическая пластичность напрямую связана с силой связи между двумя нейронами, включая количество нейромедиаторов, высвобождаемых из пресинаптического нейрона , и реакцию, генерируемую в постсинаптическом нейроне . Несинаптическая пластичность предполагает изменение возбудимости нейронов в аксоне , дендритах и соме отдельного нейрона, удаленного от синапса.
Синаптическая пластичность
[ редактировать ]Синаптическая пластичность — это способность синапса между двумя нейронами изменять силу с течением времени. Синаптическая пластичность вызвана изменениями в использовании синаптического пути, а именно частоты синаптических потенциалов и рецепторов, используемых для передачи химических сигналов. Синаптическая пластичность играет большую роль в обучении и памяти в мозге. Синаптическая пластичность может происходить за счет внутренних механизмов, при которых изменения силы синапсов происходят из-за его собственной активности, или за счет внешних механизмов, при которых изменения силы синапсов происходят через другие нервные пути. Кратковременное ингибирование синаптической пластичности часто возникает из-за ограниченного поступления нейротрансмиттеров в синапс, а долговременное ингибирование может происходить за счет снижения экспрессии рецепторов в постсинаптических клетках . Кратковременная комплементарная синаптическая пластичность часто возникает из-за остаточного или повышенного потока ионов как в пресинаптическом, так и в постсинаптическом терминале, в то время как долговременная синаптическая пластичность может возникать за счет увеличения продукции ионов. Глутаматные рецепторы AMPA и NMDA , среди прочего, находятся в постсинаптической клетке. [ 1 ]
Несинаптическая пластичность
[ редактировать ]Для сравнения, несинаптическая пластичность — менее известная и несколько новая и продолжающаяся область исследований в нейробиологии. Это проявляется через изменения характеристик несинаптических структур, таких как сома (биология) , аксон или дендриты. Несинаптическая пластичность может иметь краткосрочные или долгосрочные эффекты. Одним из способов возникновения этих изменений является модификация потенциалзависимых каналов в дендритах и аксонах, что меняет интерпретацию возбуждающих или тормозящих потенциалов, распространяющихся в клетку. Например, аксональную несинаптическую пластичность можно наблюдать, когда потенциал действия не достигает пресинаптического окончания из-за низкой проводимости или накопления ионов. [ 2 ]

Синергетические эффекты
[ редактировать ]Общие возбуждающие эффекты
[ редактировать ]Было показано, что несинаптическая и синаптическая пластичность действуют одновременно различными способами, оказывая стимулирующее воздействие на нейрон. Сюда входит генерация спайков, продукта несинаптической регуляции калиевых и других пресинаптических ионных каналов, которые усиливают реакцию возбуждающего постсинаптического потенциала за счет высвобождения нейромедиатора и увеличения потенциала действия. [ 3 ] Несинаптическая дендритная пластичность также усиливает эффекты синаптической пластичности за счет расширения потенциала действия. головного мозга Как будет обсуждаться далее, нейротрофический фактор (BNDF) продуцируется нейронами для координации несинаптической и синаптической пластичности. [ 4 ] Несинаптические изменения в сомальном теле, аксоне или дендритах нейрона неразрывно связаны с синаптической силой.
Интеграция памяти и обучения
[ редактировать ]Хотя о роли синаптической пластичности в памяти и обучении известно гораздо больше, как синаптическая, так и несинаптическая пластичность важны для памяти и обучения в мозге. Существует множество свидетельств того, что оба механизма работают синергетически для достижения наблюдаемых эффектов. Ключевым примером этого является формирование памяти в синапсе, при котором модификация пресинаптических механизмов высвобождения и постсинаптических рецепторов влияет либо на долговременную потенциацию, либо на депрессию. С другой стороны, непрерывная сомальная деполяризация была предложена как метод обучения поведению и памяти посредством несинаптической пластичности. Несинаптическая пластичность также увеличивает эффективность формирования синаптической памяти за счет регуляции потенциалзависимых ионных каналов . Несинаптическая пластичность — это механизм, ответственный за модификации этих каналов в аксоне, приводящие к изменению силы потенциала действия нейронов, неизменно влияющие на силу синаптических механизмов и, следовательно, на глубину и длину кодирования памяти. [ 5 ] [ 6 ]
Регуляция синаптической пластичности
[ редактировать ]Несинаптическая пластичность также обладает способностью регулировать эффекты синаптической пластичности посредством отрицательной обратной связи механизмов . Изменение количества и свойств ионных каналов в аксоне или дендритах может уменьшить эффекты гиперстимулированного синапса. [ 5 ] [ 6 ] В случае чрезмерного перевозбуждения этих ионных каналов произойдет обратный поток ионов в клетку, что приведет к эксайтотоксичности и гибели клеток в результате апоптоза или некроза . [ 7 ]
Внутренние механизмы
[ редактировать ]Несинаптические области нейронов, такие как аксон, также обладают присущими им свойствами, влияющими на синапс. Эти важные механизмы включают задержку деполяризации, которой подвергается потенциал действия при движении по аксону. Это внутреннее качество замедляет распространение потенциалов действия и обусловлено движением деполяризующего тока вниз по цитоплазме и прерывистым размещением натриевых каналов в узлах Ранвье . Эти механизмы существуют всегда, но могут меняться в зависимости от состояния сомы, аксона и дендритов клетки в данный момент. Следовательно, латентность или задержка распространения потенциалов действия или возбуждающих постсинаптических потенциалов может быть переменной. Каждый возбуждающий постсинаптический потенциал , который распространяется на постсинаптическую клетку, сначала передается через потенциал действия вниз по аксону пресинаптической клетки, и, таким образом, несинаптическая пластичность по своей сути влияет на синаптическую пластичность. [ 1 ]
Типы
[ редактировать ]
Собственная возбудимость нейрона
[ редактировать ]Возбудимость нейрона в любой точке зависит от внутреннего и внешнего состояния клетки в момент раздражения. Поскольку нейрон обычно получает несколько входящих сигналов одновременно, распространение потенциала действия зависит от интеграции всех входящих возбуждающих и тормозных постсинаптических потенциалов, поступающих к аксонному бугорку . Если сумма всех возбуждающих и тормозящих сигналов деполяризует клеточную мембрану до порогового напряжения, возникает потенциал действия. Изменение внутренней возбудимости нейрона изменит функцию этого нейрона.
Генерация шипов
[ редактировать ]Несинаптическая пластичность оказывает возбуждающее действие на генерацию спайков . Увеличение генерации спайков коррелирует со снижением порога спайков . [ 3 ] ответ несинаптической пластичности. Этот ответ может быть результатом модуляции определенных пресинаптических K + Токи ионов калия (IA , IK ,Ca и IKs ), которые повышают возбудимость сенсорных нейронов, расширяют потенциал действия и усиливают высвобождение нейромедиаторов . Эти модуляции K + проводимости служат общими механизмами регуляции возбудимости и силы синапсов . [ 5 ]
Регуляция синаптической пластичности
[ редактировать ]Несинаптическая пластичность связана с синаптической пластичностью как через синергетические, так и через регуляторные механизмы. Степень синаптической модификации определяет полярность несинаптических изменений, влияющих на изменение клеточной возбудимости. Умеренные уровни синаптической пластичности вызывают несинаптические изменения, которые будут синергически действовать с синаптическими механизмами, усиливая ответ. И наоборот, более устойчивые уровни синаптической пластичности будут вызывать несинаптические реакции, которые будут действовать как механизм отрицательной обратной связи . Механизмы отрицательной обратной связи работают для защиты от насыщения или подавления активности схемы в целом. [ 5 ]
Аксональная модуляция
[ редактировать ]Аксональная модуляция — это тип пластичности, при котором изменяется количество, активность или расположение ионных каналов в аксоне. Это заставляет нейрон вести себя по-разному при стимуляции. Модуляция ионных каналов является ответом на изменение частот стимуляции нейрона.
Пластичность распространения
[ редактировать ]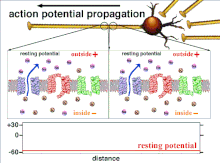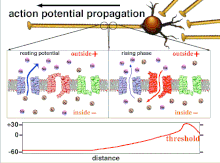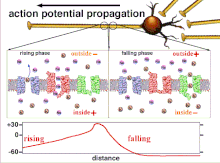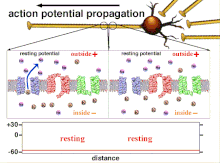
Поскольку именно суммирование потенциалов действия в конечном итоге приводит к пересечению пороговой поляризации, временные взаимоотношения различных входных сигналов очень важны для определения того, сработает ли и когда постсинаптический нейрон. Со временем время, необходимое потенциалу действия для распространения по длине определенного аксона, может измениться. В одном эксперименте многоэлектродные матрицы использовались для измерения времени, необходимого потенциалу действия для прохождения от одного электрода к другому, называемого задержкой. Затем нейроны стимулировали и значение задержки записывали с течением времени. Значения латентности со временем менялись, что позволяет предположить, что аксональная пластичность влияет на распространение потенциалов действия. [ 8 ]
Шунтирование
[ редактировать ]Шунтирование — это процесс, при котором аксональные ионные каналы открываются во время пассивного потока (не требующего ионного насоса ) подпороговой деполяризации по аксону. Обычно возникает в точках ветвления аксонов, [ 9 ] время открытия этих каналов при поступлении подпорогового сигнала в эту область приводит к добавлению гиперполяризации к пассивно протекающей деполяризации. Следовательно, клетка способна контролировать, через какие ветви аксона протекает ток подпороговой деполяризации, в результате чего некоторые ветви аксона более гиперполяризуются, чем другие. Эти различные мембранные потенциалы приводят к тому, что определенные области нейрона становятся более возбудимыми, чем другие, в зависимости от конкретного местоположения и возникновения шунтирования.
Высокочастотная стимуляция
[ редактировать ]Краткосрочные эффекты: Высокочастотная стимуляция нейрона в течение короткого периода времени увеличивает возбудимость нейрона за счет снижения величины напряжения, необходимого для возникновения потенциала действия. [ 3 ] Высокочастотная стимуляция приводит к увеличению внутриклеточной концентрации ионов натрия и кальция за счет повторного открытия потенциалзависимых натриевых и кальциевых каналов в аксоне и терминали. По мере увеличения частоты стимулов между каждым стимулом остается меньше времени для реполяризации клетки и возвращения к нормальному потенциалу покоя . Следовательно, потенциал покоя становится более деполяризованным, а это означает, что для запуска потенциала действия необходим меньший деполяризующий ток.
Однако эта модуляция обычно очень недолговечна. Если стимуляция прекратится, нейрон вернется к своему первоначальному потенциалу покоя, поскольку у ионных каналов и насосов будет достаточно времени, чтобы восстановиться после последнего стимула.
Долгосрочные эффекты: Высокочастотная стимуляция нейрона в течение длительного периода времени вызывает два результирующих нейрональных изменения. Первоначально нейрон реагирует так же, как и при кратковременной стимуляции, повышением возбудимости. Продолжение высокочастотной стимуляции после этого момента приводит к резкому, необратимому изменению возбудимости. Когда концентрация натрия в аксоне достигает достаточно высокого уровня, натрий/кальциевые насосы меняют направление потока, заставляя кальций импортироваться в клетку, а натрий выводится наружу. Повышенная концентрация кальция (и последующая деполяризация мембраны) инактивирует натриевые каналы и направляет их на эндоцитоз и лизосомальный гидролиз . [ 10 ] Это приводит к значительному уменьшению аксональных натриевых каналов, которые необходимы для распространения потенциала действия. Если стимуляция продолжится, в конечном итоге нейрон перестанет передавать потенциалы действия и умрет. Гибель нейронов из-за чрезмерной стимуляции называется эксайтотоксичностью .
Низкочастотная стимуляция
[ редактировать ]Краткосрочные эффекты: Все живые нейроны имеют базальную скорость распространения потенциала действия и синаптического высвобождения. Таким образом, низкочастотная стимуляция нейрона в краткосрочном периоде аналогична активности нейрона в состоянии покоя в мозге. Никаких серьезных изменений во внутренней возбудимости нейрона не происходит.
Долгосрочные эффекты: Низкочастотная стимуляция нейрона в течение длительного периода времени снижает возбудимость нейрона за счет активации кальций-зависимых фосфатаз, которые метят АМРА-рецепторы для интернализации. [ 11 ] Низкочастотная стимуляция приводит к снижению уровня кальция в клетке. При низких концентрациях кальция активные кальций-зависимые фосфатазы доминируют над кальций-зависимыми киназами. По мере активации большего количества фосфатаз они помечают больше АМРА-рецепторов для интернализации посредством эндоцитоза. Поскольку АМРА-рецепторы являются одними из основных возбуждающих рецепторов нейронов, их удаление из клеточной мембраны эффективно угнетает клетку (если клетка не может реагировать на возбуждающие сигналы, она не может генерировать собственный потенциал действия). Таким образом, низкочастотная стимуляция может фактически обратить вспять эффекты долговременной потенциации . [ 12 ] однако эти концепции обычно считаются типами синаптической пластичности.
Гомеостатическая и Хеббианская пластичность
[ редактировать ]Нейроны центральной нервной системы (ЦНС) интегрируют сигналы от многих нейронов. В краткосрочной перспективе важно иметь изменения в активности нейрона, потому что именно так информация передается в нервной системе ( пластичность Хебба ). Однако для долгосрочной устойчивости дрейф в сторону возбудимости или невозбудимости нарушит способность цепи передавать информацию ( гомеостатическая пластичность ). Долговременная потенциация (LTP) вызывает более высокую частоту импульсов в постсинаптических нейронах. Была выдвинута гипотеза, что внутренние свойства нейрона должны быть организованы так, чтобы максимально использовать динамический диапазон, действуя как гомеостатический механизм. [ 13 ] Однако было показано, что внутренняя возбудимость подчиняется логарифмически нормальному распределению, что требует поддержания активного обучения по Хеббу. [ 14 ] Исследования in vitro показали, что когда спонтанная активность нейрональных культур подавляется, нейроны становятся гипервозбудимыми, а когда увеличение активности индуцируется в течение длительного времени, скорость возбуждения культуры падает. [ 15 ] [ 16 ] Напротив, существует множество свидетельств того, что также имеет место противоположная форма регуляции — обучение по Хеббу или LTP-IE/LTD-IE. [ 17 ] а теоретические аргументы показывают, что пластичность Хебба должна быть доминирующей формой пластичности и для внутренней возбудимости. [ 14 ] Поскольку гомеостатическая пластичность возникает и между отдельными синапсами, [ 18 ] Было показано, что более ранняя точка зрения, предполагающая, что гомеостатическая пластичность и внутренняя пластичность связаны между собой, не соответствует фактическим данным.
Механизм
[ редактировать ]Одним из механизмов сохранения динамического диапазона нейрона является синаптическое масштабирование , гомеостатическая форма пластичности, которая восстанавливает активность нейронов до ее нормального «базового» уровня путем изменения постсинаптического ответа синапсов нейрона в зависимости от активности. Гомеостатическая модуляция внутренней возбудимости нейрона — еще один способ поддержания стабильности. Регулирования ионной проводимости можно добиться разными способами, в основном за счет высвобождения нейромодуляторов, таких как дофамин , серотонин и т. д. [ 19 ] Другой способ — контролируемое высвобождение нейротрофического фактора головного мозга (BDNF). Также было обнаружено, что BDNF влияет на синаптическое масштабирование, что позволяет предположить, что этот нейротрофический фактор может быть ответственным за координацию синаптических и несинаптических механизмов гомеостатической пластичности. [ 4 ]
Дендритная возбудимость
[ редактировать ]Дендриты — это области, ответственные за интеграцию входных сигналов от других нейронов . Одним из способов, с помощью которого нейроны манипулируют интеграционными свойствами дендритов, является изменение количества и свойств потенциалзависимых ионных каналов. Индукция долговременной потенциации (LTP) в определенном синапсе приводит к увеличению возбудимости дендритных ветвей, специфичных для этого синапса. [ 20 ] Дендритная возбудимость важна для распространения и интеграции синаптических сигналов. Считается, что дендритная возбудимость способствует потенцированию ES или увеличению вероятности того, что данный вход приведет к срабатыванию потенциала действия. [ 21 ]
Известно, что изменения возбудимости дендритов влияют на обратное распространение потенциала действия. Потенциалы действия начинаются возле бугорка аксона и распространяются по всей длине аксона, но они также распространяются назад через сому в дендритное ветвь. Активное обратное распространение зависит от ионных каналов, и изменение плотности или свойств этих каналов может влиять на степень ослабления сигнала . [ 21 ] Пластичность обратного распространения в дендритах происходит менее чем за одну минуту и продолжается более 25 минут. [ 22 ] Обратное распространение — это метод передачи сигнала синапсам о срабатывании потенциала действия. Это важно для пластичности, зависящей от времени всплеска . Экспериментально наблюдалась быстрая дендритная адаптация за несколько секунд, что указывает на потенциально значимый глобальный механизм обучения. [ 23 ] [ 24 ]
Внутренняя пластичность
[ редактировать ]Внутренняя пластичность — это форма пластичности, зависящей от активности, отличная от синаптической пластичности , которая включает изменения в синапсе между двумя нейронами, а не изменения электрических свойств внутри одного нейрона. [ 25 ] [ 26 ] Есть некоторые тесно связанные явления, которые могут повлиять на возбудимость нейрона, такие как нейромодуляция , структурная пластичность, кратковременная пластичность из-за кинетики каналов и развитие нейронов . [ 27 ] [ 28 ] Не существует единого мнения относительно величины, которую регулирует внутренняя пластичность, например, частота срабатывания нейрона, его прирост или внутренняя концентрация кальция. Функционально внутренняя пластичность может позволить нейронам узнавать интенсивность стимулов и представлять статистику этой интенсивности в их возбудимости. [ 29 ] [ 30 ] Внутренняя пластичность способствует кодированию памяти и дополняет другие формы пластичности, зависящей от активности, включая синаптическую пластичность . [ 31 ]
Высшая функция мозга
[ редактировать ]Долговременная ассоциативная память
[ редактировать ]Экспериментальные доказательства
[ редактировать ]Эксперимент Кеменеса и др . [ 2 ] продемонстрировали, что во внешнем модуляторном нейроне несинаптическая пластичность влияет на выражение долговременной ассоциативной памяти . Взаимосвязь несинаптической пластичности и памяти оценивали с использованием гигантских клеток головного мозга (CGC). Деполяризация от условных раздражителей усиливала реакцию нейрональной сети. Эта деполяризация длилась столько же, сколько и долговременная память . Стойкая деполяризация и проявление поведенческой памяти произошли более чем через 24 часа после тренировки, что указывает на долгосрочные последствия. В этом эксперименте электрофизиологическое выражение следа долговременной памяти представляло собой реакцию на кормление, вызванную условным стимулом. CGC были значительно более деполяризованы у тренированных организмов, чем у контрольной группы, что указывает на связь с изменениями обучения и возбудимости. Когда CGC были деполяризованы, они демонстрировали повышенную реакцию на условные раздражители и более сильную фиктивную реакцию на кормление. Это продемонстрировало, что деполяризации достаточно, чтобы вызвать значительную пищевую реакцию на условные раздражители. Кроме того, не наблюдалось существенной разницы в скорости питания между кондиционированными организмами и организмами, которые были искусственно деполяризованы, что еще раз подтверждает, что деполяризации достаточно, чтобы вызвать поведение, связанное с долговременной памятью. [ 2 ]
Память
[ редактировать ]Несинаптическая активность в клетке обычно выражается в изменении возбудимости нейронов. Это происходит посредством модуляции компонентов мембраны, таких как каналы покоя и потенциалзависимые каналы и ионные насосы . Считается, что несинаптические процессы участвуют в хранении памяти. Один из возможных механизмов этого действия предполагает маркировку нейрона, который недавно был активен, с изменением возбудимости. Это помогло бы связать разделенные во времени стимулы. Другой потенциальный механизм исходит из вычислительной модели, которая показывает, что несинаптическая пластичность может стимулировать цепи для модификации в обучении, поскольку изменения возбудимости могут регулировать порог синаптической пластичности. [ 5 ]
Емкость систем хранения памяти на основе синапсов очень велика, что делает их привлекательным механизмом для изучения. Есть примерно 10 4 синапсов на нейрон и 10 11 нейроны в человеческом мозге. [ 25 ] Несинаптическую пластичность часто упускают из виду просто потому, что ее емкость хранения не так высока. Регулирование плотности ионных каналов в аксоне и соме нейрона изменит пропускную способность и повлияет на все синапсы. Следовательно, его емкость хранения будет значительно меньше, чем у синаптической пластичности.
Хотя его емкость слишком мала, чтобы сделать его единственным механизмом хранения, несинаптическая пластичность может способствовать синаптическим методам хранения. Было показано, что модуляция ионных каналов может происходить в таких маленьких областях, как отдельные дендриты. [ 20 ] Эта специфичность делает емкость несинаптической пластичности большей, чем если бы она рассматривалась как модуляция целого нейрона. Процедурная память хорошо подходит для этого типа системы хранения, поскольку не требует такой высокой специфичности, как декларативная память. Обобщение двигательных задач и условных раздражителей может быть эффективным способом хранения этой информации. [ 25 ]
Обучение
[ редактировать ]Изменения возбудимости в результате обучения, которые действуют как часть следа памяти , служат праймерами для инициирования дальнейших изменений в нейронах или механизмом кратковременного хранения кратковременной памяти. Несинаптическая пластичность может возникать во время обучения в результате клеточных процессов, хотя время, устойчивость и взаимосвязь между несинаптической пластичностью и синаптическим выходом плохо изучены. Исследования показали, что несинаптическая пластичность играет косвенную, но важную роль в формировании воспоминаний. Несинаптическая пластичность, индуцированная обучением, связана с деполяризацией сомы. [ 5 ]
Классическое кондиционирование
[ редактировать ]Эксперименты показали, что во время условного обучения происходят несинаптические изменения . Вуди и др . [ 32 ] продемонстрировали, что кондиционирование глазного моргания (EBC), форма классического кондиционирования для изучения нервных структур и механизмов, лежащих в основе обучения и памяти, у кошек связана с повышенной возбудимостью и активностью нейронов в сенсомоторных областях коры и в лицевом ядре . Было замечено, что повышение возбудимости, вызванное классической обусловливанием, продолжалось после прекращения реакции. Это предполагает, что повышенная возбудимость может функционировать как механизм хранения памяти. [ 5 ]
При обусловливании моргания у кроликов несинаптические изменения происходили во всем дорсальном гиппокампе . Это указывает на то, что хотя одних изменений возбудимости недостаточно для объяснения процессов хранения памяти, несинаптическая пластичность может быть механизмом хранения фаз памяти, ограниченных временем. Несинаптические изменения влияют на другие типы пластичности, связанные с памятью. Например, несинаптические изменения, такие как деполяризация мембранного потенциала покоя , возникающие в результате условного обучения, могут вызвать синаптическую пластичность в будущем обучении. [ 5 ]
Правило обучения и экономии
[ редактировать ]Способность изучать правила зависит от несинаптической пластичности. Одно исследование было направлено на то, чтобы научить крыс различать различные запахи, и потребовалось несколько дней, чтобы научить их различать первую пару запахов. Однако, научившись этому, крыса смогла научиться различать разные запахи гораздо быстрее. Изменения возбудимости пирамидных нейронов у этих крыс наблюдались в течение трех суток после тренировки. Эти изменения со временем исчезли, что позволяет предположить, что нейроны участвовали в изучении правил, а не в хранении памяти. [ 5 ] Даудаль и Дебанн попытались определить, применимы ли те же правила обучения и механизмы индукции, определенные для синаптической пластичности, к несинаптической пластичности, влияющей на ионные каналы. Они определили, что несинаптическая и синаптическая пластичность имеют общие правила обучения и пути индукции, например, рецептора NMDA долговременную потенциацию, зависимую от (LTP) и долговременную депрессию (LTD). Они также показали, что несинаптическая и синаптическая пластичность синергически образуют связную энграмму для хранения следов памяти. [ 22 ]
Экономия – это возможность переучить забытую информацию гораздо быстрее, чем она была усвоена изначально. Несинаптическая пластичность является возможным механизмом этого эффекта экономии. Во время тренировок многие нейроны испытывают повышение внутренней возбудимости. Такое повышение возбудимости сохраняется даже после угасания памяти. [ 5 ] [ 25 ]
Зависимость от веществ
[ редактировать ]Злоупотребление наркотиками обычно влияет на мезолимбическую систему или, точнее, на путь вознаграждения нервной системы. Среди распространенных наркотиков, вызывающих злоупотребление, никотин является одним из сильнейших агонистов никотинового холинергического синапса. [ 33 ] Никотин, конкурируя с ацетилхолином (АХ), действует через несинаптический претерминальный никотиновый ацетилхолиновый рецептор (нАХР), инициируя изменение мембранного потенциала и распространяя внутриклеточный Са. 2+ сигнал, тем самым способствуя высвобождению нейротрансмиттеров. Специфическая и характерная роль активности nAChR, опосредованной кальциевым током, имеет иную зависимость от напряжения, чем у других Ca 2+ проницаемые ионные каналы, а также различное временное и пространственное распределение и, как следствие, несинаптическая активность нАХР усиливает индукцию синаптической потенциации, способствуя обучению зависимости от веществ. [ 34 ]
Приложения к болезням
[ редактировать ]После повреждения
[ редактировать ]Несинаптическая пластичность может смягчать последствия повреждения головного мозга . Когда один из вестибулярных нервов поврежден, несоответствие частоты импульсов нейронов вестибулярных ядер вызывает ненужные вестибулярные рефлексы. Симптомы этого повреждения со временем исчезают. Вероятно, это связано с изменениями внутренней возбудимости нейронов вестибулярного ядра. [ 25 ] [ 35 ]
Судорожная активность
[ редактировать ]Несинаптическая пластичность также играет ключевую роль в судорожной активности. Фебрильные судороги, судороги, вызванные лихорадкой в раннем возрасте, могут привести к повышенной возбудимости нейронов гиппокампа. Эти нейроны становятся высокочувствительными к противосудорожным агентам. Было показано, что судороги в раннем возрасте могут предрасполагать к возникновению новых припадков посредством несинаптических механизмов. [ 36 ]
Травма, включая инсульт , приводящий к повреждению коры головного мозга , часто приводит к эпилепсии . Повышенная возбудимость и проводимость NMDA приводят к эпилептической активности, что позволяет предположить, что несинаптическая пластичность может быть механизмом, посредством которого индуцируется эпилепсия после травмы. [ 37 ]
Аутизм
[ редактировать ]Вальпроевая кислота (VPA) — это средство для лечения эпилепсии , мигрени и биполярного расстройства , которое связано со многими заболеваниями, включая аутизм . Существует животная модель аутизма , при которой беременным крысам дают VPA. Потомство имеет черты, схожие с чертами людей с аутизмом. Вскоре после рождения у этих животных наблюдается снижение возбудимости и увеличение NMDA токов . Эти эффекты корректируются на более поздних этапах жизни. Изменения внутренней возбудимости у этих животных помогли компенсировать влияние повышенных токов NMDA на сетевую активность, форму гомеостатической пластичности. Считается, что это помогает смягчить пагубные последствия, которые могут иметь повышенные токи NMDA. [ 38 ]
Текущие и будущие исследования
[ редактировать ]Для более широкого понимания несинаптической пластичности необходимы дополнительные исследования. Темы, которые требуют дальнейшего изучения по состоянию на январь 2010 г. [update] включать:
- Локальные и глобальные изменения возбудимости в нейронных сетях и поддержание следа памяти [ 5 ]
- Специфика индукции изменений возбудимости, зависящих от обучения [ 5 ]
- Манипулирование зависящими от обучения изменениями возбудимости с помощью фармацевтических продуктов или генетических мутаций и их влияние на след памяти [ 5 ]
- Сходство молекулярных механизмов синаптической и несинаптической пластичности [ 5 ]
- Сравнение паттернов несинаптической пластичности in vivo с результатами in vitro [ 5 ]
- Изменения в экспрессии генов, вызванные нейронной активностью [ 39 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Бирн, Джон Х. (1997). «Синаптическая пластичность» . Нейронаука онлайн . Медицинская школа UT в Хьюстоне . Проверено 28 октября 2011 г.
- ^ Перейти обратно: а б с Кеменес И., Штрауб В.А., Никитин Е.С., Старас К., О'Ши М., Кеменес Г., Бенджамин П.Р. (июль 2006 г.). «Роль отсроченной несинаптической пластичности нейронов в долговременной ассоциативной памяти» . Современная биология . 16 (13): 1269–79. дои : 10.1016/j.cub.2006.05.049 . ПМИД 16824916 . S2CID 16726488 .
- ^ Перейти обратно: а б с Гензель С., Линден DJ, Д'Анджело Э (май 2001 г.). «За пределами параллельных волокон LTD: разнообразие синаптической и несинаптической пластичности в мозжечке». Природная неврология . 4 (5): 467–75. дои : 10.1038/87419 . ПМИД 11319554 . S2CID 13919286 .
- ^ Перейти обратно: а б Десаи Н.С., Резерфорд Л.К., Турриджано Г.Г. (1999). «BDNF регулирует внутреннюю возбудимость корковых нейронов» . Обучение и память . 6 (3): 284–91. дои : 10.1101/lm.6.3.284 . ПМК 311300 . ПМИД 10492010 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Моццачиоди Р., Бирн Дж. Х. (январь 2010 г.). «Больше, чем синаптическая пластичность: роль несинаптической пластичности в обучении и памяти» . Тенденции в нейронауках . 33 (1): 17–26. дои : 10.1016/j.tins.2009.10.001 . ПМЦ 2815214 . ПМИД 19889466 .
- ^ Перейти обратно: а б Дебанн Д., Копысова И.Л., Брас Х., Ферран Н. (сентябрь 1999 г.). «Запуск распространения потенциала действия посредством аксональной А-подобной калиевой проводимости в гиппокампе: новый тип несинаптической пластичности». Журнал физиологии, Париж . 93 (4): 285–96. дои : 10.1016/S0928-4257(00)80057-1 . ПМИД 10574118 . S2CID 30737916 .
- ^ Шидловска К., Тимянски М. (февраль 2010 г.). «Кальций, ишемия и эксайтотоксичность». Клеточный кальций . 47 (2): 122–9. дои : 10.1016/j.ceca.2010.01.003 . ПМИД 20167368 .
- ^ Баккум DJ, Чао З.К., Поттер С.М. (май 2008 г.). «Долговременная пластичность задержки и амплитуды распространения потенциала действия в корковых сетях, зависящая от активности» . ПЛОС ОДИН . 3 (5): е2088. Бибкод : 2008PLoSO...3.2088B . дои : 10.1371/journal.pone.0002088 . ПМК 2324202 . ПМИД 18461127 .

- ^ Дебанн Д., Гэвилер Б.Х., Томпсон С.М. (1996). «Синаптическая и несинаптическая пластичность между отдельными пирамидными клетками в гиппокампе крысы in vitro». Журнал физиологии, Париж . 90 (5–6): 307–9. дои : 10.1016/s0928-4257(97)87903-x . ПМИД 9089497 . S2CID 31639170 .
- ^ Ахмед З., Верашко А (январь 2009 г.). «Зависимая от активности аксональная пластичность: влияние электрической стимуляции на потенциалы действия соединений, зарегистрированные в нервной системе мыши in vitro». Открытый журнал неврологии . 3 (1): 6. Бибкод : 2009ONMJ....1....6P . doi : 10.2174/1874082000903010001 (неактивен 27 апреля 2024 г.). S2CID 14646213 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Лин Дж.В., Джу В., Фостер К., Ли Ш., Ахмадиан Г., Вышински М. и др. (декабрь 2000 г.). «Различные молекулярные механизмы и различные эндоцитотические пути интернализации рецептора AMPA» . Природная неврология . 3 (12): 1282–90. дои : 10.1038/81814 . ПМИД 11100149 . S2CID 18109725 .
- ^ О'Делл Т.Дж., Кандел Э.Р. (1994). «Низкочастотная стимуляция уничтожает LTP посредством активации протеинфосфатаз, опосредованной рецептором NMDA» . Обучение и память . 1 (2): 129–139. дои : 10.1101/lm.1.2.129 . ПМИД 10467591 . S2CID 41429873 .
- ^ Стеммлер М., Кох С. (июнь 1999 г.). «Как проводимость, зависящая от напряжения, может адаптироваться для максимизации информации, кодируемой скоростью срабатывания нейронов». Природная неврология . 2 (6): 521–7. дои : 10.1038/9173 . ПМИД 10448216 . S2CID 15134486 .
- ^ Перейти обратно: а б Шелер Г. (2017). «Логарифмические распределения доказывают, что внутреннее обучение является хеббианским» . F1000Исследования . 6 : 1222. arXiv : 1410.5610 . дои : 10.12688/f1000research.12130.2 . ПМК 5639933 . ПМИД 29071065 .
- ^ Корнер М.А., Рамакерс Г.Дж. (январь 1992 г.). «Спонтанная стимуляция как эпигенетический фактор развития мозга - физиологические последствия хронического воздействия тетродотоксина и пикротоксина на культивируемые нейроны неокортекса крыс». Исследования мозга. Исследования развития мозга . 65 (1): 57–64. дои : 10.1016/0165-3806(92)90008-К . ПМИД 1551233 .
- ^ Турриджано Г.Г., Лесли К.Р., Десаи Н.С., Резерфорд Л.К., Нельсон С.Б. (февраль 1998 г.). «Зависимое от активности масштабирование квантовой амплитуды в нейронах неокортекса». Природа . 391 (6670): 892–6. Бибкод : 1998Natur.391..892T . дои : 10.1038/36103 . ПМИД 9495341 . S2CID 4328177 .
- ^ Махон С., Шарпье С. (август 2012 г.). «Двунаправленная пластичность внутренней возбудимости контролирует эффективность сенсорных входов в нейронах коры головного мозга 5-го слоя in vivo» . Журнал неврологии . 32 (33): 11377–89. doi : 10.1523/JNEUROSCI.0415-12.2012 . ПМК 6621180 . ПМИД 22895720 .
- ^ Кек Т., Келлер ГБ, Якобсен Р.И., Эйзель Ю.Т., Бонхёффер Т., Хюбенер М. (октябрь 2013 г.). «Синаптическое масштабирование и гомеостатическая пластичность в зрительной коре мыши in vivo» . Нейрон . 80 (2): 327–34. дои : 10.1016/j.neuron.2013.08.018 . ПМИД 24139037 . S2CID 13151568 .
- ^ Шелер Г. (2014). «Изучение внутренней возбудимости средних шипиковых нейронов» . F1000Исследования . 2 : 88. arXiv : q-bio/0502023 . дои : 10.12688/f1000research.2-88.v2 . ПМЦ 4264637 . ПМИД 25520776 .
- ^ Перейти обратно: а б Фрик А., Маги Дж., Джонстон Д. (февраль 2004 г.). «ЛТП сопровождается повышенной местной возбудимостью дендритов пирамидных нейронов». Природная неврология . 7 (2): 126–35. дои : 10.1038/nn1178 . ПМИД 14730307 . S2CID 11964239 .
- ^ Перейти обратно: а б Сьёстрем П.Дж., Ранц Э.А., Рот А., Хойссер М. (апрель 2008 г.). «Дендритная возбудимость и синаптическая пластичность». Физиологические обзоры . 88 (2): 769–840. doi : 10.1152/physrev.00016.2007 . ПМИД 18391179 . S2CID 1261675 .
- ^ Перейти обратно: а б Даудал Дж., Дебанн Д. (2003). «Долговременная пластичность внутренней возбудимости: правила и механизмы обучения» . Обучение и память . 10 (6): 456–65. дои : 10.1101/lm.64103 . ПМИД 14657257 .
- ^ Ходассман, Шири; Варди, Рони; Тугендхафт, Яэль; Голденталь, Амир; Кантер, Идо (декабрь 2022 г.). «Эффективное дендритное обучение как альтернатива гипотезе синаптической пластичности» . Научные отчеты . 12 (1): 6571. Бибкод : 2022NatSR..12.6571H . дои : 10.1038/s41598-022-10466-8 . ISSN 2045-2322 . ПМК 9051213 . ПМИД 35484180 .
- ^ Сарди, Шира; Варди, Рони; Голденталь, Амир; Шейнин, Антон; Узан, Херут; Кантер, Идо (23 марта 2018 г.). «Адаптивные узлы обогащают нелинейное совместное обучение, выходя за рамки традиционной адаптации посредством связей» . Научные отчеты . 8 (1): 5100. Бибкод : 2018НатСР...8.5100С . дои : 10.1038/s41598-018-23471-7 . ISSN 2045-2322 . ПМЦ 5865176 . ПМИД 29572466 .
- ^ Перейти обратно: а б с д и Чжан В., Linden DJ (ноябрь 2003 г.). «Другая сторона энграммы: обусловленные опытом изменения внутренней возбудимости нейронов». Обзоры природы. Нейронаука . 4 (11): 885–900. дои : 10.1038/nrn1248 . ПМИД 14595400 . S2CID 17397545 .
- ^ Дебанн Д., Инглеберт Ю., Руссье М. (февраль 2019 г.). «Пластичность внутренней возбудимости нейронов» (PDF) . Современное мнение в нейробиологии . 54 : 73–82. дои : 10.1016/j.conb.2018.09.001 . ПМИД 30243042 . S2CID 52812190 .
- ^ Триш, Йохен. « Синергия между внутренней и синаптической пластичностью в отдельных модельных нейронах » . Достижения в области нейронных систем обработки информации . 2004.
- ^ В. Чжан и DJ Линден. Другая сторона энграммы: вызванные опытом изменения внутренней возбудимости нейронов. Nature Reviews Neuroscience, 4:885-900, 2003.
- ^ Т. Монк, К. Савин и Дж. Лак. « Оптимальный нейронный вывод об интенсивности стимулов ». Научные отчеты, 8:1, 2018.
- ^ Т. Монк, К. Савин и Дж. Лак. Нейроны, обладающие внутренней пластичностью, изучают статистику интенсивности стимулов. Достижения в области нейронных систем обработки информации . 2016.
- ^ Грасселли Г., Боеле Х.Дж., Титли Х.К., Брэдфорд Н., ван Бирс Л., Джей Л. и др. (январь 2020 г.). «Каналы SK2 в клетках Пуркинье мозжечка способствуют модуляции возбудимости в следах памяти, специфичных для моторного обучения» . ПЛОС Биология . 18 (1): e3000596. дои : 10.1371/journal.pbio.3000596 . ПМК 6964916 . ПМИД 31905212 .
- ^ Компакт-диск Вуди, Black-Cleworth P (ноябрь 1973 г.). «Различия в возбудимости корковых нейронов в зависимости от двигательной проекции у кондиционированных кошек». Журнал нейрофизиологии . 36 (6): 1104–16. дои : 10.1152/jn.1973.36.6.1104 . ПМИД 4761722 .
- ^ Кауэр Дж.А., Маленка Р.С. (ноябрь 2007 г.). «Синаптическая пластичность и зависимость» . Обзоры природы. Нейронаука . 8 (11): 844–58. дои : 10.1038/nrn2234 . ПМИД 17948030 . S2CID 38811195 .
- ^ Дэни Дж.А., Джи Д., Чжоу Ф.М. (август 2001 г.). «Синапсическая пластичность и никотиновая зависимость» . Нейрон . 31 (3): 349–52. дои : 10.1016/S0896-6273(01)00379-8 . ПМИД 11516393 . S2CID 10062998 .
- ^ Дарлингтон К.Л., Дутиа М.Б., Смит П.Ф. (июнь 2002 г.). «Вклад собственной возбудимости нейронов вестибулярного ядра в восстановление после вестибулярного повреждения». Европейский журнал неврологии . 15 (11): 1719–27. дои : 10.1046/j.1460-9568.2002.02024.x . ПМИД 12081651 . S2CID 19939794 .
- ^ Бендер Р.А., Дубе С., Гонсалес-Вега Р., Мина Э.В., Барам Т.З. (2003). «Пластичность мшистых волокон и повышенная возбудимость гиппокампа без потери клеток гиппокампа или изменения нейрогенеза на животной модели длительных фебрильных судорог» . Гиппокамп . 13 (3): 399–412. дои : 10.1002/hipo.10089 . ПМЦ 2927853 . ПМИД 12722980 .
- ^ Буш ПК, Принс Д.А., Миллер К.Д. (октябрь 1999 г.). «Повышенная пирамидная возбудимость и проводимость NMDA могут объяснить посттравматический эпилептогенез без растормаживания: модель». Журнал нейрофизиологии . 82 (4): 1748–58. дои : 10.1152/jn.1999.82.4.1748 . ПМИД 10515964 . S2CID 5927906 .
- ^ Уолкотт Э.К., Хиггинс Э.А., Десаи Н.С. (сентябрь 2011 г.). «Синаптический и внутренний баланс во время постнатального развития у крысят, подвергшихся воздействию вальпроевой кислоты внутриутробно» . Журнал неврологии . 31 (37): 13097–109. doi : 10.1523/JNEUROSCI.1341-11.2011 . ПМЦ 6623264 . ПМИД 21917793 .
- ^ Лесли Дж. Х., Недиви Э (август 2011 г.). «Гены, регулирующие активность, как медиаторы пластичности нейронных цепей» (PDF) . Прогресс нейробиологии . 94 (3): 223–37. doi : 10.1016/j.pneurobio.2011.05.002 . hdl : 1721.1/102275 . ПМК 3134580 . ПМИД 21601615 .