Связывающий сайт

В биохимии и молекулярной биологии сайт связывания — это участок макромолекулы , например белка связывается с другой молекулой , который специфично . [ 1 ] Партнера по связыванию макромолекулы часто называют лигандом . [ 2 ] Лиганды могут включать другие белки (приводящие к белок-белковому взаимодействию ), [ 3 ] ферментные субстраты , [ 4 ] вторичные мессенджеры , гормоны или аллостерические модуляторы . [ 5 ] Событие связывания часто, но не всегда, сопровождается конформационными изменениями белка , которые изменяют функцию . [ 6 ] Связывание с сайтами связывания белка чаще всего является обратимым (временным и нековалентным ), но может быть и ковалентным обратимым. [ 7 ] или необратимый. [ 8 ]
Функция
[ редактировать ]Связывание лиганда с сайтом связывания на белке часто вызывает изменение конформации белка и приводит к изменению клеточной функции. Следовательно, сайт связывания на белке является важной частью путей передачи сигнала . [ 9 ] Типы лигандов включают нейротрансмиттеры , токсины , нейропептиды и стероидные гормоны . [ 10 ] Сайты связывания вызывают функциональные изменения в ряде контекстов, включая ферментный катализ, передачу сигналов молекулярных путей, гомеостатическую регуляцию и физиологические функции. Электрический заряд , стерическая форма и геометрия сайта избирательно позволяют связываться высокоспецифичным лигандам, активируя определенный каскад клеточных взаимодействий, за которые отвечает белок. [ 11 ] [ 12 ]
Катализ
[ редактировать ]
Ферменты осуществляют катализ, более прочно связываясь с переходными состояниями, чем субстраты и продукты. В сайте каталитического связывания на субстрат могут действовать несколько различных взаимодействий. Они варьируются от электрического катализа, кислотного и основного катализа, ковалентного катализа и катализа ионами металлов. [ 10 ] Эти взаимодействия уменьшают энергию активации химической реакции, обеспечивая благоприятные взаимодействия для стабилизации молекулы с высокой энергией. Связывание ферментов обеспечивает более близкое сближение и исключение веществ, не имеющих отношения к реакции. Это специфическое связывание также препятствует побочным реакциям. [ 13 ] [ 10 ]
Типы ферментов, которые могут выполнять эти действия, включают оксидоредуктазы, трансферазы, гидролазы, лиазы, изомеразы и лигазы. [ 14 ]
Например, трансфераза-гексокиназа катализирует фосфорилирование глюкозы с образованием глюкозо-6-фосфата. Остатки активного центра гексокиназы позволяют стабилизировать молекулу глюкозы в активном центре и стимулируют начало альтернативного пути благоприятных взаимодействий, уменьшая энергию активации. [ 15 ]
Торможение
[ редактировать ]Ингибирование белка путем связывания ингибитора может вызвать нарушение регуляции путей, гомеостатической регуляции и физиологических функций.
Конкурентные ингибиторы конкурируют с субстратом за связывание со свободными ферментами в активных центрах и, таким образом, препятствуют образованию фермент-субстратного комплекса при связывании. Например, отравление угарным газом вызвано конкурентным связыванием угарного газа с кислородом в гемоглобине.
неконкурентные ингибиторы Альтернативно, связываются одновременно с субстратом в активных центрах. При связывании с комплексом фермент-субстрат (ES) образуется комплекс ингибитора фермент-субстрат (ESI). Как и в случае с конкурентными ингибиторами, скорость образования продукта также снижается. [ 4 ]
Наконец, смешанные ингибиторы способны связываться как со свободным ферментом, так и с комплексом фермент-субстрат. Однако, в отличие от конкурентных и неконкурентных ингибиторов, смешанные ингибиторы связываются с аллостерическим сайтом. Аллостерическое связывание вызывает конформационные изменения, которые могут повысить сродство белка к субстрату. Это явление называется положительной модуляцией. И наоборот, аллостерическое связывание, которое снижает сродство белка к субстрату, является отрицательной модуляцией. [ 16 ]
Типы
[ редактировать ]Активный сайт
[ редактировать ]В активном центре субстрат связывается с ферментом, вызывая химическую реакцию. [ 17 ] [ 18 ] Субстраты, переходные состояния и продукты могут связываться с активным центром, а также с любыми конкурентными ингибиторами. [ 17 ] Например, в контексте функции белка связывание кальция с тропонином в мышечных клетках может вызывать конформационные изменения тропонина. Это позволяет тропомиозину обнажить участок связывания актин-миозина, с которым связывается головка миозина, образуя поперечный мостик и вызывая мышечное сокращение . [ 19 ]
В контексте крови примером конкурентного связывания является окись углерода, которая конкурирует с кислородом за активный центр гема . Высокое сродство монооксида углерода может превзойти кислород в присутствии низкой концентрации кислорода. В этих обстоятельствах связывание монооксида углерода вызывает изменение конформации, которое препятствует связыванию гема с кислородом, что приводит к отравлению угарным газом. [ 4 ]

Аллостерический сайт
[ редактировать ]В регуляторном сайте связывание лиганда может вызывать усиление или ингибирование функции белка. [ 4 ] [ 20 ] Связывание лиганда с аллостерическим сайтом мультимерного фермента часто индуцирует положительную кооперативность, то есть связывание одного субстрата вызывает благоприятное изменение конформации и увеличивает вероятность связывания фермента со вторым субстратом. [ 21 ] Лиганды регуляторного сайта могут включать гомотропные и гетеротропные лиганды, в которых один или несколько типов молекул соответственно влияют на активность фермента. [ 22 ]
Ферменты, которые строго регулируются, часто играют важную роль в метаболических путях. Например, фосфофруктокиназа (ФФК), которая фосфорилирует фруктозу при гликолизе, в значительной степени регулируется АТФ. Его регуляция в гликолизе необходима, поскольку это обязательный и ограничивающий скорость этап пути. ПФК также контролирует количество глюкозы, предназначенной для образования АТФ по катаболическому пути. Следовательно, при достаточных уровнях АТФ ПФК аллостерически ингибируется АТФ. Эта регуляция эффективно сохраняет запасы глюкозы, которые могут потребоваться для других путей. Цитрат, промежуточный продукт цикла лимонной кислоты, также действует как аллостерический регулятор ПФК. [ 22 ] [ 23 ]
Одно- и многоцепочечные сайты связывания
[ редактировать ]Сайты связывания можно охарактеризовать также их структурными особенностями. Одноцепочечные сайты (у «монодесмических» лигандов, μόνος: одиночный, δεσμός: связывающий) образованы одной белковой цепью, а многоцепочечные сайты (у «полидесмических» лигандов, πολοί: многие) [ 24 ] часто встречаются в белковых комплексах и образуются лигандами, которые связывают более чем одну белковую цепь, обычно в границах раздела белков или вблизи них. Недавние исследования показывают, что структура сайта связывания имеет глубокие последствия для биологии белковых комплексов (эволюция функции, аллостерия). [ 25 ] [ 26 ]
Загадочные сайты связывания
[ редактировать ]Загадочные сайты связывания представляют собой сайты связывания, которые временно образуются в апо-форме или индуцируются связыванием лиганда. Учет загадочных сайтов связывания увеличивает размер потенциально « лекарственного » человеческого протеома с ~40% до ~78% белков, связанных с заболеванием. [ 27 ] Сайты связывания были исследованы с помощью: машины опорных векторов , примененной к набору данных «CryptoSite», [ 27 ] Расширение набора данных «КриптоСайт», [ 28 ] долговременное моделирование молекулярной динамики с использованием модели состояния Маркова и биофизических экспериментов, [ 29 ] и индекс загадочных сайтов, основанный на относительной доступной площади поверхности . [ 30 ]
Привязка кривых
[ редактировать ]
Кривые связывания описывают поведение связывания лиганда с белком. Кривые могут характеризоваться своей формой, сигмоидальной или гиперболической, которая отражает, проявляет ли белок кооперативное или некооперативное связывание соответственно. [ 31 ] Обычно ось X описывает концентрацию лиганда, а ось Y описывает фракционное насыщение лигандов, связанных со всеми доступными сайтами связывания. [ 4 ] Уравнение Михаэлиса Ментена обычно используется при определении формы кривой. Уравнение Михаэлиса Ментена выведено на основе стационарных условий и учитывает ферментативные реакции, происходящие в растворе. Однако когда реакция происходит, когда фермент связан с субстратом, кинетика протекает по-другому. [ 32 ]
Моделирование с помощью кривых связывания полезно при оценке сродства связывания кислорода с гемоглобином и миоглобином в крови. Гемоглобин, имеющий четыре гемовые группы, обладает кооперативным связыванием . Это означает, что связывание кислорода с гемовой группой гемоглобина вызывает благоприятное конформационное изменение, которое позволяет повысить благоприятность связывания кислорода для следующих гемовых групп. В этих обстоятельствах кривая связывания гемоглобина будет иметь сигмоидальную форму из-за его повышенной способности связываться с кислородом. Поскольку миоглобин имеет только одну гемовую группу, он демонстрирует некооперативное связывание, которое на кривой связывания имеет гиперболический характер. [ 33 ]
Приложения
[ редактировать ]Биохимические различия между различными организмами и людьми полезны для разработки лекарств . Например, пенициллин убивает бактерии, ингибируя бактериальный фермент DD-транспептидазу , разрушая развитие бактериальной клеточной стенки и вызывая гибель клеток. Таким образом, изучение сайтов связывания актуально для многих областей исследований, включая механизмы рака, [ 34 ] лекарственная форма, [ 35 ] и физиологическая регуляция. [ 36 ] Приготовление ингибитора для подавления функции белка является распространенной формой фармацевтической терапии. [ 37 ]
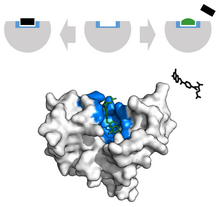
В области рака лиганды, которые отредактированы, чтобы иметь внешний вид, аналогичный природному лиганду, используются для ингибирования роста опухоли. Например, Метотрексат препарат химиотерапевтический действует как конкурентный ингибитор активного центра дигидрофолатредуктазы . [ 38 ] Это взаимодействие ингибирует синтез тетрагидрофолата , прекращая производство ДНК, РНК и белков. [ 38 ] Подавление этой функции подавляет рост опухолей и улучшает состояние при тяжелом псориазе и ревматоидном артрите у взрослых . [ 37 ]
При сердечно-сосудистых заболеваниях такие препараты, как бета-блокаторы, используются для лечения пациентов с гипертонией. Бета-блокаторы (β-блокаторы) представляют собой антигипертензивные средства, которые блокируют связывание гормонов адреналина и норадреналина с β1- и β2-рецепторами в сердце и кровеносных сосудах. Эти рецепторы обычно опосредуют симпатическую реакцию «бей или беги», вызывая сужение кровеносных сосудов. [ 39 ]
Конкурентные ингибиторы также широко встречаются в продаже. Ботулинический токсин , известный под торговым названием «Ботокс», представляет собой нейротоксин , вызывающий вялый паралич мышц из-за связывания с ацетилхолин-зависимыми нервами. Это взаимодействие подавляет мышечные сокращения, создавая видимость гладких мышц. [ 40 ]
Прогноз
[ редактировать ]Был разработан ряд вычислительных инструментов для прогнозирования местоположения сайтов связывания на белках. [ 20 ] [ 41 ] [ 42 ] Их можно в общих чертах разделить на основанные на последовательности и основанные на структуре. [ 42 ] Методы, основанные на последовательностях, основаны на предположении, что последовательности функционально консервативных частей белков, таких как сайт связывания, консервативны. Структурные методы требуют трехмерной структуры белка. Эти методы, в свою очередь, можно разделить на шаблонные и карманные. [ 42 ] Методы, основанные на шаблонах, ищут трехмерное сходство между целевым белком и белками с известными сайтами связывания. Методы, основанные на карманах, ищут вогнутые поверхности или скрытые карманы в целевом белке, которые обладают такими характеристиками, как гидрофобность и способность к образованию водородных связей , которые позволяют им связывать лиганды с высоким сродством. [ 42 ] Несмотря на то, что здесь используется термин «карман», аналогичные методы можно использовать для прогнозирования сайтов связывания, используемых в белок-белковых взаимодействиях, которые обычно более плоские, а не в карманах. [ 43 ]
Ссылки
[ редактировать ]- ^ «Связующий сайт» . Медицинские предметные рубрики (MeSH) . Национальная медицинская библиотека США.
Части макромолекулы, непосредственно участвующие в ее специфическом сочетании с другой молекулой.
- ^ «Лиганды» . Медицинские предметные рубрики (MeSH) . Национальная медицинская библиотека США.
Молекула, которая связывается с другой молекулой, особенно используется для обозначения небольшой молекулы, которая специфически связывается с более крупной молекулой.
- ^ Амос-Бинкс А., Патулеа С., Питре С., Шенрок А., Ги Ю., Грин-младший, Гольшани А., Дене Ф. (июнь 2011 г.). «Предсказание сайта связывания для белок-белковых взаимодействий и открытие новых мотивов с использованием повторяющихся полипептидных последовательностей» . БМК Биоинформатика . 12 :225. дои : 10.1186/1471-2105-12-225 . ПМК 3120708 . ПМИД 21635751 .
- ^ Перейти обратно: а б с д и Хардин CC, Кнопп Дж.А. (2013). «Глава 8: Ферменты». Биохимия – основные понятия . Нью-Йорк: Издательство Оксфордского университета. стр. 51–69. ISBN 978-1-62870-176-0 .
- ^ Кенакин Т.П. (апрель 2016 г.). «Особенности аллостеризма в действии лекарств» . В Бауэри Н.Г. (ред.). Модуляция аллостерических рецепторов при нацеливании лекарств . ЦРК Пресс. п. 26. ISBN 978-1-4200-1618-5 .
- ^ Спитцер Р., Клевес А.Е., Варела Р., Джайн А.Н. (апрель 2014 г.). «Аннотация функции белка по сходству поверхности локального сайта связывания» . Белки . 82 (4): 679–94. дои : 10.1002/прот.24450 . ПМЦ 3949165 . ПМИД 24166661 .
- ^ Бандиопадхьяй А., Гао Дж. (октябрь 2016 г.). «Нацеливание на биомолекулы с помощью обратимой ковалентной химии» . Современное мнение в области химической биологии . 34 : 110–116. дои : 10.1016/j.cbpa.2016.08.011 . ПМК 5107367 . ПМИД 27599186 .
- ^ Беллелли А., Кэри Дж. (январь 2018 г.). «Обратимое связывание лиганда» . Обратимое связывание лиганда: теория и эксперимент . Джон Уайли и сыновья. п. 278. ИСБН 978-1-119-23848-5 .
- ^ Сюй Д., Джалал С.И., Следж Г.В., Меруэ СО (октябрь 2016 г.). «Сайты связывания малых молекул для изучения белок-белковых взаимодействий в протеоме рака» . Молекулярные биосистемы . 12 (10): 3067–87. дои : 10.1039/c6mb00231e . ПМК 5030169 . ПМИД 27452673 .
- ^ Перейти обратно: а б с Уилсон К. (март 2010 г.). Принципы и методы биохимии и молекулярной биологии . Издательство Кембриджского университета. стр. 581–624. дои : 10.1017/cbo9780511841477.016 . ISBN 9780511841477 .
- ^ Ахерн К. (2015). Биохимия бесплатно для всех . Государственный университет Орегона. стр. 110–141.
- ^ Кумар А.П., Лукман С (6 июня 2018 г.). «Аллостерические сайты связывания в Rab11 для потенциальных кандидатов в лекарства» . ПЛОС ОДИН . 13 (6): e0198632. Бибкод : 2018PLoSO..1398632K . дои : 10.1371/journal.pone.0198632 . ПМК 5991966 . ПМИД 29874286 .
- ^ Добсон Дж.А., Джеррард Эй.Дж., Пратт Дж.А. (2008). Основы химической биологии . Издательство Оксфордского университета. ISBN 9780199248995 . OCLC 487962823 .
- ^ Аззарони О, Шлейфер I (04 декабря 2017 г.). Полимерные и биополимерные щетки . дои : 10.1002/9781119455042 . ISBN 978-1-119-45501-1 .
- ^ Словарь пищевой науки и технологий (2-е изд.). Международная служба информации о продуктах питания. 2009. ISBN 978-1-4051-8740-4 .
- ^ Кларк К.Г. (2013). Биотехнологическая инженерия . Издательство Вудхед. стр. 79–84. дои : 10.1533/9781782421689 . ISBN 978-1-78242-167-2 .
- ^ Перейти обратно: а б Уилсон К. (март 2010 г.). «Ферменты» . В Уилсон К., Уокер Дж. (ред.). Принципы и методы биохимии и молекулярной биологии . Издательство Кембриджского университета. стр. 581–624. дои : 10.1017/cbo9780511841477.016 . ISBN 9780511841477 . Проверено 1 ноября 2018 г.
- ^ Шашке С (2014). Словарь химической инженерии . Издательство Оксфордского университета. ISBN 978-1-62870-844-8 .
- ^ Моррис Дж (2016). Биология Как устроена жизнь . Соединенные Штаты Америки: WH Freeman and Company. стр. 787–792. ISBN 978-1-4641-2609-3 .
- ^ Перейти обратно: а б Конц Дж, Янежич Д (апрель 2014 г.). «Сравнение сайтов связывания для прогнозирования функций и открытия фармацевтических препаратов». Современное мнение в области структурной биологии . 25 : 34–9. дои : 10.1016/j.sbi.2013.11.012 . ПМИД 24878342 .
- ^ Фукуа С, Уайт Д (2004). «Прокариотическая межклеточная передача сигналов». Передача сигналов в клетках у прокариот и низших многоклеточных животных . Спрингер Нидерланды. стр. 27–71. дои : 10.1007/978-94-017-0998-9_2 . ISBN 9789048164837 .
- ^ Перейти обратно: а б Крейтон Т.Э. (2010). Биофизическая химия нуклеиновых кислот и белков . Гельветская пресса. ISBN 978-0956478115 . OCLC 760830351 .
- ^ Каррелл Б.Р., ван Дам-Миерас MC (1997). Биотехнологические инновации в химическом синтезе . Оксфорд: Баттерворт-Хайнеманн. стр. 125–128. ISBN 978-0-7506-0561-8 .
- ^ Абрусан Дж., Марш Дж.А. (2019). «Структура сайта связывания лиганда формирует сворачивание, сборку и деградацию гомомерных белковых комплексов» . Журнал молекулярной биологии . 431 (19): 3871–3888. дои : 10.1016/j.jmb.2019.07.014 . ПМК 6739599 . ПМИД 31306664 .
- ^ Абрусан Г., Марш Дж.А. (2018). «Структура сайта связывания лиганда влияет на эволюцию функции и топологии белкового комплекса» . Отчеты по ячейкам . 22 (12): 3265–3276. дои : 10.1016/j.celrep.2018.02.085 . ПМЦ 5873459 . ПМИД 29562182 .
- ^ Абрусан Дж., Марш Дж.А. (2019). «Структура сайта связывания лиганда формирует аллостерическую передачу сигнала и эволюцию аллостерии в белковых комплексах» . Молекулярная биология и эволюция . 36 (8): 1711–1727. дои : 10.1093/molbev/msz093 . ПМК 6657754 . ПМИД 31004156 .
- ^ Перейти обратно: а б Чимерманчич П., Вайнкам П., Реттенмайер Т.Дж., Бичманн Л., Киди Д.А., Волдайес Р.А. и др. (февраль 2016 г.). «Криптосайт: расширение протеома, поддающегося лекарственному воздействию, путем характеристики и прогнозирования загадочных сайтов связывания» . Журнал молекулярной биологии . 428 (4): 709–719. дои : 10.1016/j.jmb.2016.01.029 . ПМЦ 4794384 . ПМИД 26854760 .
- ^ Беглов Д., Холл Д.Р., Уэйкфилд А.Е., Луо Л., Аллен К.Н., Козаков Д. и др. (апрель 2018 г.). «Изучение структурного происхождения загадочных участков белков» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (15): Е3416–Е3425. Бибкод : 2018PNAS..115E3416B . дои : 10.1073/pnas.1711490115 . ПМК 5899430 . ПМИД 29581267 .
- ^ Боуман Г.Р., Болин Э.Р., Харт К.М., Магуайр Б.С., Маркузи С. (март 2015 г.). «Обнаружение множества скрытых аллостерических сайтов путем объединения моделей состояния Маркова и экспериментов» . Труды Национальной академии наук Соединенных Штатов Америки . 112 (9): 2734–9. Бибкод : 2015PNAS..112.2734B . дои : 10.1073/pnas.1417811112 . ПМЦ 4352775 . ПМИД 25730859 .
- ^ Иида С., Накамура Х.К., Машимо Т., Фукуниси Ю. (ноябрь 2020 г.). «Структурные колебания ароматических остатков в апо-форме выявляют загадочные места связывания: значение для разработки лекарств на основе фрагментов». Журнал физической химии Б. 124 (45): 9977–9986. doi : 10.1021/acs.jpcb.0c04963 . ПМИД 33140952 . S2CID 226244554 .
- ^ Ахерн К. (январь 2017 г.). «Преподавание биохимии онлайн в Университете штата Орегон» . Образование в области биохимии и молекулярной биологии . 45 (1): 25–30. дои : 10.1002/bmb.20979 . ПМИД 27228905 .
- ^ Энн А., Демайль С. (октябрь 2012 г.). «Кинетика действия ферментов на поверхностно-прикрепленные субстраты: практическое руководство по анализу кривой прогресса в любой кинетической ситуации». Ленгмюр . 28 (41): 14665–71. дои : 10.1021/la3030827 . ПМИД 22978617 .
- ^ Моррис-младший, Хартл Д.Л., Нолл А.Х. (19 ноября 2015 г.). Биология: как устроена жизнь (Второе изд.). Нью-Йорк, штат Нью-Йорк. ISBN 9781464126093 . OCLC 937824456 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Спитцер Р., Клевес А.Е., Варела Р., Джайн А.Н. (апрель 2014 г.). «Аннотация функции белка по сходству поверхности локального сайта связывания» . Белки . 82 (4): 679–94. дои : 10.1002/прот.24450 . ПМЦ 3949165 . ПМИД 24166661 .
- ^ Пэн Дж, Ли XP (ноябрь 2018 г.). «Аполипопротеин A-IV: потенциальная терапевтическая мишень при атеросклерозе». Простагландины и другие липидные медиаторы . 139 : 87–92. doi : 10.1016/j.prostaglandins.2018.10.004 . ПМИД 30352313 . S2CID 53023273 .
- ^ Макнамара Дж.В., Садаяппан С. (декабрь 2018 г.). «Скелетный миозин-связывающий протеин-С: все более важный регулятор физиологии поперечно-полосатых мышц» . Архив биохимии и биофизики . 660 : 121–128. дои : 10.1016/j.abb.2018.10.007 . ПМК 6289839 . ПМИД 30339776 .
- ^ Перейти обратно: а б Видеманн, Британская Колумбия, Адамсон, ПК (июнь 2006 г.). «Понимание и управление нефротоксичностью метотрексата» . Онколог . 11 (6): 694–703. doi : 10.1634/теонколог.11-6-694 . ПМИД 16794248 .
- ^ Перейти обратно: а б Раджагопалан П.Т., Чжан З., МакКорт Л., Дуайер М., Бенкович С.Дж., Хаммес Г.Г. (октябрь 2002 г.). «Взаимодействие дигидрофолатредуктазы с метотрексатом: ансамблевая и одномолекулярная кинетика» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (21): 13481–6. Бибкод : 2002PNAS...9913481R . дои : 10.1073/pnas.172501499 . ПМК 129699 . ПМИД 12359872 .
- ^ Фришман В.Х., Ченг-Лай А., Чен Дж., ред. (2000). Современные сердечно-сосудистые препараты . дои : 10.1007/978-1-4615-6767-7 . ISBN 978-1-57340-135-7 . S2CID 38187984 .
- ^ Монтекукко К., Молго Дж. (июнь 2005 г.). «Ботулинические нейротоксины: возрождение старого убийцы». Современное мнение в фармакологии . 5 (3): 274–9. дои : 10.1016/j.coph.2004.12.006 . ПМИД 15907915 .
- ^ Рош Д.Б., Бракенридж Д.А., Макгаффин Ж.Дж. (декабрь 2015 г.). «Белки и их взаимодействующие партнеры: введение в методы прогнозирования сайта связывания белка с лигандом» . Международный журнал молекулярных наук . 16 (12): 29829–42. дои : 10.3390/ijms161226202 . ПМЦ 4691145 . ПМИД 26694353 .
- ^ Перейти обратно: а б с д Брумхед Н.К., Солиман М.Е. (март 2017 г.). «Можем ли мы полагаться на вычислительные прогнозы для правильной идентификации сайтов связывания лигандов на новых мишенях белковых лекарств? Оценка методов прогнозирования сайтов связывания и протокола проверки предсказанных сайтов связывания». Клеточная биохимия и биофизика . 75 (1): 15–23. дои : 10.1007/s12013-016-0769-y . ПМИД 27796788 . S2CID 6705144 .
- ^ Джонс С., Торнтон Дж. М. (сентябрь 1997 г.). «Анализ мест белок-белкового взаимодействия с использованием поверхностных пятен». Журнал молекулярной биологии . 272 (1): 121–32. дои : 10.1006/jmbi.1997.1234 . ПМИД 9299342 .
Внешние ссылки
[ редактировать ]- Связывающие сайты Национальной медицинской библиотеки США в медицинских предметных рубриках (MeSH)
- Рисунок активного центра фермента