Анатомия гиппокампа


Анатомия гиппокампа описывает физические аспекты и свойства гиппокампа , нервную структуру в медиальной височной мозга доле . Он имеет отличительную, изогнутую форму, которая была сравнена с морским монстром греческой мифологии и рогам Амуна в египетской мифологии . Этот общий макет удерживается во всем диапазоне видов млекопитающих , от ежи до человека, хотя детали различаются. Например, у крысы два гиппокампа похожи на пару бананов, соединенных на стеблях. В мозге приматов , включая людей, часть гиппокампа возле основания височной доли намного шире, чем часть вверху. Из-за трехмерной кривизны этой структуры обычно наблюдаются двумерные срезы, такие как показанные. На рисунках нейровизуализации могут показать ряд различных форм, в зависимости от угла и расположения разреза.
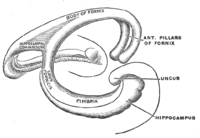
Топологически поверхность полушария головного мозга можно рассматривать как сферу с отступлением, где она прикрепляется к среднему мозгу. Структуры, которые выстраивают край отверстия в совокупности, составляют так называемую лимбическую систему (латинский лимб = граница ), с гиппокампом, выстилающим задний край этого отверстия. Эти лимбические структуры включают в себя гиппокамп, поясную кору , обонятельную кору и миндалину . Пол Маклин однажды предложил, как часть своей триединой теории мозга, что лимбические структуры представляют собой нервную основу эмоций . В то время как большинство нейробиологов больше не верят в концепцию единой «лимбической системы», эти области сильно взаимосвязаны и взаимодействуют друг с другом. [ Цитация необходима ]
Основная цепь гиппокампа
[ редактировать ]
Начиная с зубчатой извилины и работая внутрь по S-кривой гиппокампа означает пересечение Серия узких зон. Первая из них, зубчатая извилина (DG), на самом деле отдельная Структура, плотно упакованный слой маленьких гранулярных ячеек, обернутых вокруг конца самого гиппокампа , образуя заостренный клин в некоторых поперечных сечениях, полукруг в других. Следующий Приходите серию районов Cornu Ammonis : сначала CA4 (который лежит в основе зубчатой извилины), затем CA3 , затем Очень маленькая зона под названием CA2 , затем CA1 . Все области CA заполнены плотно упакованными пирамидальными клетками, похожими на те, которые обнаружены в неокортексе . После CA1 поступает область, называемая субикулумом . После этого приходит пара плохо определенных областей, называемых преподавато-паразобикулумом, затем Переход к собственному коре (в основном энторинальная область коры). Большинство анатомистов Используйте термин «собственно гиппокамп» для обозначения четырех полей CA и образования гиппокампа Чтобы обозначить собственный гиппокамп плюс зубчатая извилина и субикулум. [ 1 ]
Основные сигнальные пути протекают через гиппокамп и объединяются с образованием петли. Большая часть внешнего ввода поступает из соседней энторинальной коры через аксоны так называемого перфорантного пути . Эти аксоны возникают из слоя 2 энторинальной коры (EC) и заканчиваются в зубчатой извилине и CA3. Существует также отчетливый путь от слоя 3 ЕС непосредственно к CA1, который часто называют Themoamamonic или Ta-Ca1. Гранулярные ячейки DG посылают их аксоны (называемые «мшистыми волокнами») в CA3. Пирамидальные ячейки CA3 посылают свои аксоны в CA1. Пирамидальные клетки CA1 посылают свои аксоны в субикулум и глубокие слои ЕС. Субикулярные нейроны посылают свои аксоны в основном в ЕС. Перфорантный путь к днату извивались за традисинаптическую цепь с помощью Андерсена, который отметил, что тонкие срезы могут быть вырезаны из гиппокампа, перпендикулярного его длинной оси, таким образом, который сохраняет все эти соединения. Это наблюдение было основой его пластинчатой гипотезы , который предположил, что гиппокамп можно рассматривать как серию параллельных полос, работающих функционально независимым образом. [ 2 ] Пластичная концепция по -прежнему считается полезным принципом организации, но более поздние данные, демонстрирующие обширные продольные соединения в системе гиппокампа, потребовали, чтобы она была существенно изменена. [ 3 ]
Перфорантный путь ввода от слоя EC II входит в зубчатую извилину и передается в область CA3 (и на мошные ячейки, расположенные в гилусе зубчатой извилины, которая затем отправляет информацию в отдаленные части зубчатой извилины, где повторяется цикл). Региона CA3 объединяет этот вход с сигналами от уровня EC II и посылает обширные соединения в регионе, а также посылает соединения со стратными радиатумом и Oriens ипсилатеральных и контралатеральных областей CA1 через набор волокон, называемых коллатералями Шаффера , и комиссуральные путь, соответственно. [ 4 ] [ 5 ] [ 6 ] Региона CA1 получает вход от подполя CA3, уровня EC III и воссоединения ядра таламуса (которые проецируются только на терминальные апикальные дендритные пучки в лакуносум-молекуляре Stratum ). В свою очередь, CA1 проецирует в субикулум, а также отправляет информацию по вышеупомянутым путям выхода гиппокампа. Субикулум является последней стадией в пути, объединяя информацию из проекции CA1 и уровня EC III, чтобы также отправлять информацию вдоль выходных путей гиппокампа.
Гиппокамп также получает ряд подкорковых входов. В Macaca Fecusicularis эти входные данные включают миндалину (в частности, переднюю аминдалоидную область, базолатеральное ядро и периамигдалоидную кору), медиальную перегородку и диагональную полосу брока , клаустрам , субстанция инноминации и -ядер Мейнера базал таламус (включая передний атомный комплекс, лакологическое ядро, паравентрикулярные и паратиониальные ядра, воссоединение ядра и ядра Centralis Medialis), боковые преоптические и латеральные области гипоталамуса , надммиллярные и ретромамиллярные области, вентральная область , тегментарная устойчивость, raphe nuclei (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи (нуклеи ( нуклеи нуклеи (нуклеи (нукля Sentralis Superior и дорсальный раф Ядро), ядро reticularis tegementi pontis , периакедуктальный серый , дорсальное сегментарное ядро и локус coeruleus . Гиппокамп также получает прямые моносинаптические проекции из ядра мозжечка . [ 7 ]
Основные системы волокон в крысах
[ редактировать ]Угловой пакет
[ редактировать ]Эти волокна начинаются с вентральной части энторинальной коры (EC) и содержат комиссуральный (EC ◀ ▶ Гиппокамп) и перфорантного пути (возбуждающий EC ▶ CA1 и ингибирующий EC ▶ CA2 [ 8 ] ) волокна. Они путешествуют вдоль септоропочной оси гиппокампа. Волокна перфорационного пути, как следует из названия, перфорирует субикулум перед тем, как отправиться в гиппокамп (поля CA) и зубчатую извилину. [ 9 ]
Путь Fringe-Fornix
[ редактировать ]
Волокна Fimbria-Fornix представляют собой гиппокамп и субикулярный шлюз в и из подкорковых областей мозга. [ 10 ] [ 11 ] Различные части этой системы дают разные названия:
- Белые миелинизированные волокна, которые покрывают желудочковые (глубокие) части гиппокампа, делают альвеус .
- Волокна, которые покрывают височные части гиппокампа, делают пакет волокна, который называется Fimbria . Переход от височной в перегородку (дорсальные) части гиппокампа Fimbria собирает все больше и больше гиппокампа и более плотных выходов и становится толще.
- В средней линии и под мозолистом корпуса эти волокна образуют Fornix .
На уровне схемы альве содержит аксональные волокна из DG и из пирамидных нейронов CA3, CA2, CA1 и субикулума ( CA1 ▶ Субикулум и CA1 ▶ Энторинальные проекции), которые собираются во временном гиппокойсе для формирования фимбрии/Форникс, один из Основные выходы гиппокампа. [ 12 ] [ 13 ] [ 14 ] [ 15 ] [ 16 ] У крысы некоторые медиальные и латеральные энторинальные аксоны ( энтроринальная проекция ▶ CA1 ) проходят через альвеус к молекуляру LACUNOSUM CA1, не делая значительного числа эн -пассивных буторов на других слоях CA1 ( ThemoRoammonic Alvear Pathway ). [ 13 ] [ 17 ] Контралатеральные энторинальные проекции CA1 почти исключительно проходят через Альвеус. Чем больше перегородки, тем больше ипсилатеральных проекций Entorhinal-Ca1, которые проходят путь Alvear (вместо перфорантного пути). [ 18 ] Хотя аксоновые посылают аксональные проекции в аксональные проекции в водохранилище, проекция C1 проходит через восток Stratata и молекуляр немного и CA1. [ 19 ] Холинергические и ГАМКергические проекции от MS-DBB до CA1 также проходят через Fimbria. [ 20 ] Стимуляция фимбрии приводит к холинергическому возбуждению клеток CA1 O-LMR . [ 21 ]
Также известно, что внеклеточная стимуляция фимбрии стимулирует антидромически и ортодромически, но она не влияет на клетки гранулярных клеток. [ 22 ] Каждая пирамидальная ячейка CA1 также посылает аксональную ветвь в Фимбрию. [ 23 ] [ 24 ]
Гиппокампальные комиссии
[ редактировать ]Хилальные мшистые клетки и пирамидальные клетки CA3 являются основным источником комиссуральных волокон гиппокампа . Они проходят через гиппокампа, чтобы достичь контралатеральных областей гиппокампа. Комиссы гиппокампа имеют дорсальные и вентральные сегменты. Спинные комиссуральные волокна состоит в основном из энторинальных и преподавательных волокон к или из гиппокампа и зубчатой извилины. [ 9 ] Как правило, можно сказать, что каждая цитоархитектоническая область, которая вносит вклад в комиссуральную проекцию, также имеет параллельное ассоциативное волокно, которое заканчивается в ипсилатеральном гиппокампе. [ 25 ] Внутренний молекулярный слой зубчатой извилины (дендриты как гранулярных клеток, так и ГАМКергических интернейронов) получает проекцию, которая имеет как ассоциативные, так и комиссуральные волокна, главным образом, из Хилар Моссистических клеток и в некоторой степени от пирамидных клеток CA3C. Поскольку эти проекционные волокна происходят как с ипсилатеральных, так и контралатеральных сторон гиппокампа, их называют ассоциативными/комиссуральными проекциями. Фактически, каждая мошивая клетка иннервирует как ипсилатеральную, так и контралатеральную зубчатую извилину. Хорошо известная трисинаптическая цепь гиппокампа простирается в основном горизонтально вдоль гиппокампа. Тем не менее, ассоциативные/комиссуральные волокна, такие как ассоциативные проекции пирамидальных клеток CA2, охватывают в основном продольные (дорсовентрально) вдоль гиппокампа. [ 26 ] [ 27 ] Комиссуральные волокна, которые происходят из пирамидальных клеток CA3, идут в области CA3, CA2 и CA1. Как и мшистые клетки, одна пирамидальная клетка CA3 способствует как комиссуральному, так и в ассоциационных волокнах, и они заканчивают как на основных клетках, так и на интернейронах. [ 28 ] [ 29 ] Слабая комиссуральная проекция соединяет оба региона CA1 вместе. Субикулум не имеет комиссуральных входов или выходов. По сравнению с грызунами, комиссуральные связи гиппокампа гораздо менее распространены у обезьян и людей. [ 30 ] Хотя возбуждающие клетки являются основными участниками комиссуральных путей, среди их терминалов сообщалось о ГАМКергическом компоненте, которые были прослежены на Хилус как происхождение. [ 31 ] Стимуляция комиссуральных волокон стимулирует DG Hilar Perforant-ассоциированный (HIPP) и CA3 триламинные клетки антидромно. [ 32 ]
Клетки и слои гиппокампа
[ редактировать ]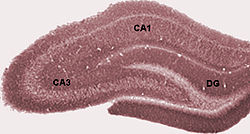

Гиппокамп
[ редактировать ]состоит Свой гиппокамп из ряда подпол. Хотя терминология варьируется среди авторов, наиболее часто используемыми терминами являются зубчатая извилина и аммонис Cornu (буквально « рог Аммона », сокращенно CA ). Значительная извилина содержит фасцию Dentata и Hilus , в то время как CA дифференцируется на подполя CA1, CA2, CA3 и CA4 .
Однако область, известная как CA4, на самом деле является «глубоким, полиморфным слоем зубчатой извилины» [ 33 ] (Как прояснено Теодором Блэкстадом (1956) [ 34 ] и Дэвида Амарала (1978)). [ 35 ]
Вырезанный в поперечном сечении , гиппокамп представляет собой С-образную структуру, которая напоминает оперативной памяти рога . Название Cornu Ammonis относится к египетскому божеству Амуна , у которого есть глава барана. Рогатый вид гиппокампа вызван дифференциалами плотности клеток и различной степенью нейрональных волокон .
У грызунов гиппокамп расположен таким образом, что примерно один конец находится рядом с верхней частью головы (дорсальный или перегороженный конец) и один конец у нижней части головы (вентральный или временный конец). Как показано на рисунке, сама структура изогнута, а подполя или области определяются вдоль кривой, от CA4 до CA1 (маркируются только CA3 и CA1). Области Ca также структурированы в глубине в четко определенных слоях (или слоях):
- Stratum Oriens (str. Oriens) является следующим слоем, поверхностным для альвеуса. Клеточные тела ингибирующих клеток корзины и горизонтальных триламинарных клеток, названные в честь их аксонов, иннервирующих три слоя - Oriens, Pyramidal и Radiatum расположены в этом слое. Базальные дендриты пирамидных нейронов также обнаружены здесь, где они получают вход от других пирамидальных клеток, перегородных волокон и комиссуральных волокон из контралатерального гиппокампа (обычно рецидивирующие связи, особенно у CA3 и CA2.) У грызунов два гиппокампа сильно подключены,, особенно в CA3 и CA2. Но в приматах эта комиссуральная связь намного разрешена.
- Pratum Pyramidale (Str. Pyr.) Содержит клеточные тела пирамидных нейронов, которые являются основными возбуждающими нейронами гиппокампа. Этот слой, как правило, является одним из наиболее заметных слоев для невооруженного глаза. В регионе CA3 этот слой содержит синапсы из мшистых волокнов, которые тратятся через Stratum lucidum. Этот слой также содержит клеточные тела многих интернейронов , в том числе аксо-аксонические клетки, бистратифицированные клетки и радиальные трилизарные клетки.
- Stratum lucidum (str. Luc.) - одна из самых тонких слоев в гиппокампе и только обнаруженная в области CA3. зубчатой извилины Мослистые волокна из гранулярных клеток через этот слой в CA3, хотя синапсы из этих волокон можно найти в STR. пир
- Радиатум Stratum (Str. Rad.), Как Str. Oriens, содержит перегородку и комиссуральные волокна. Он также содержит коллатеральные волокна Шаффера , которые представляют собой проекцию вперед от CA3 до CA1. Некоторые интернейроны, которые можно найти в более поверхностных слоях, также могут быть найдены здесь, включая клетки корзины, бистратифицированные клетки и радиальные триламинные клетки.
- Stratum lacunosum (str. Lac.) - это тонкий слой, который также содержит коллатеральные волокна Schaffer, но также содержит волокна перфорантного пути из поверхностных слоев энторинальной коры. Из-за его небольшого размера его часто сгруппируется вместе с молекуляром слоя в один слой, называемый Stratum lacunosum-Moleculare (Str. Lm.).
- Stratum moleculare (str. Mol.) - самый поверхностный слой в гиппокампе. Здесь волокна перфорантного пути образует синапсы на дистальных апикальных дендритах пирамидальных клеток.
- Служба гиппокампа (борозды) или трещина -это бесклеточная область, которая отделяет поле CA1 от зубной извилины. Поскольку фаза зарегистрированного тета -ритма систематически изменяется через слои, бороздку часто используется в качестве фиксированной отсчета для записи ЭЭГ , поскольку он легко идентифицируется. [ 33 ]
Зубчатая извилина
[ редактировать ]Зубчатая извилина состоит из аналогичной серии слоев:
- Полиморфный слой (Poly. Lay.) Является наиболее поверхностным слоем зубчатой извилины и часто считается отдельным подполе (как Hilus). Этот слой содержит много интернейронов , и аксоны зубчатых гранулярных ячеек проходят через этот слой по дороге до CA3.
- Stratum granulosum (str. Gr.) Содержит клеточные тела зубчатых гранулярных клеток.
- Moleculare Stratum, внутренняя треть (Str. Mol. 1/3) - это то, где оба комиссуральные волокна из контралатеральной зубчатой извилины пробегают и образуют синапсы, а также то, где входные данные из медиальной перегородки заканчиваются, как на проксимальных дендритах гранульных клеток.
- Молекуляр Stratum, внешние две трети (Str. Mol. 2/3) являются самым глубоким из слоев, которые находятся просто поверхностно для бороздки гиппокампа напротив молекуляции слоя в полях CA. Волокна перфорантного пути проходят через эти слои, делая возбуждающие синапсы на дистальные апикальные дендриты гранулярных клеток.
Современная база знаний о типах нейронов гиппокампа, их профиль биомаркера, активных и пассивных электрофизиологических параметров и подключение поддерживается на веб-сайте гиппокампома . [ 36 ]
Ссылки
[ редактировать ]- ^ Амарал, D; Lavenex P (2006). «CH 3. Гиппокампальная нейроанатомия». В Андерсен P; Моррис Р; Амарал D; Блаженство Т; О'Киф Дж. (Ред.). Книга гиппокампа . Издательство Оксфордского университета. ISBN 978-0-19-510027-3 .
- ^ Андерсен, P; Bliss TVP; Skrede KK (1971). «Пластичная организация возбуждающих путей гиппокампа». Эксплуат Brain Res . 13 (2): 222–238. doi : 10.1007/bf00234087 . PMID 5570425 . S2CID 12075886 .
- ^ Андерсен, P; Soleng AF; Раастад М. (2000). «Гипотеза гиппокампа Lamella пересмотрела». Brain Res . 886 (1–2): 165–171. doi : 10.1016/s0006-8993 (00) 02991-7 . PMID 11119694 . S2CID 8455285 .
- ^ Hjorth-Simonsen, A (15 января 1973 г.). «Некоторые внутренние связи гиппокампа у крысы: экспериментальный анализ». Журнал сравнительной неврологии . 147 (2): 145–61. doi : 10.1002/cne.901470202 . PMID 4118866 . S2CID 28989051 .
- ^ Swanson, LW; Wyss, JM; Коуэн, Wm (15 октября 1978 г.). «Авторадиографическое исследование организации путей интрагиппокампальной ассоциации у крыс». Журнал сравнительной неврологии . 181 (4): 681–715. doi : 10.1002/cne.901810402 . PMID 690280 . S2CID 30954240 .
- ^ Лорберг, S (15 апреля 1979 г.). «Коммиссуральные и внутренние связи крысиного гиппокампа». Журнал сравнительной неврологии . 184 (4): 685–708. doi : 10.1002/cne.901840405 . PMID 422759 . S2CID 27256712 .
- ^ Хит Р.Г., Харпер Дж.В. (ноябрь 1974 г.). «Восходящие проекции мозжечкового ядра быстрого мозжечка до гиппокампа, миндалины и других мест височной доли: вызванные потенциал и гистологические исследования у обезьян и кошек». Эксплуат Неврол . 45 (2): 268–87. doi : 10.1016/0014-4886 (74) 90118-6 . PMID 4422320 .
- ^ Мелцер, с.; Майкл, М.; Caputi, A.; Eliava, M.; Fuchs, EC; Уиттингтон, Массачусетс; Monyer, H. (22 марта 2012 г.). «Большое расстояние, пробуждающие ГАМКергические нейроны, модулируют ингибирование в гиппокампе и энторинальной коре» . Наука . 335 (6075): 1506–1510. Bibcode : 2012sci ... 335.1506M . doi : 10.1126/science.1217139 . PMID 22442486 . S2CID 206539012 .
- ^ Jump up to: а беременный Андерсен, за; и др., ред. (2007). Книга гиппокампа . Нью -Йорк: издательство Оксфордского университета. п. 47,63,123. ISBN 9780195100273 .
- ^ Пауэлл, TP; Гилью, RW; Коуэн, Wm (октябрь 1957). «Количественное исследование системы форникмамилло-талама» . Журнал анатомии . 91 (4): 419–37. PMC 1244899 . PMID 13475143 .
- ^ Дайц, HM; Пауэлл, TP (февраль 1954 г.). «Исследования связи системы Fornix» . Журнал неврологии, нейрохирургии и психиатрии . 17 (1): 75–82. doi : 10.1136/jnnp.17.1.75 . PMC 503161 . PMID 13131081 .
- ^ Ноулз, WD; Schwartzkroin, PA (ноябрь 1981 г.). «Аксональные последствия гиппокампа пирамидальных клеток Ca1» . Журнал нейробиологии . 1 (11): 1236–41. doi : 10.1523/jneurosci.01-11-01236.1981 . PMC 6564220 . PMID 6171629 .
- ^ Jump up to: а беременный Книга гиппокампа . Нью -Йорк: издательство Оксфордского университета. 2007. с. 47. ISBN 9780199723164 .
- ^ Allingay, Thomas C. Pritchard, Kevin D. (1999). Медицинская нейробиология (1 -е изд.). Мэдисон, штат Коннектикут: Паб -паб Fence Creek. п. 28. ISBN 978-1889325293 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Гаудрон, Анри М. Дюверноя, Франсуаз Каттин, Пьер-Ив Ризлд; Рисунки и иллюстрации Дж. Л. Ваннсона и М. (2013). Функциональная анатомия гиппокампа человека, васкуляризация и серийные срезы с МРТ (4 -е изд.). Берлин: Спрингер. п. 28. ISBN 978-3-642-33603-4 .
{{cite book}}: Cs1 maint: несколько имен: список авторов ( ссылка ) - ^ Тамамаки, н; Абэ, К; Nojyo, Y (14 июня 1988 г.). «Трехмерный анализ всего аксонов, происходящих из отдельных пирамидных нейронов CA2 в гиппокампе крыс с помощью компьютерного графического метода». Исследование мозга . 452 (1–2): 255–72. doi : 10.1016/0006-8993 (88) 90030-3 . PMID 3401733 . S2CID 25038544 .
- ^ Шетти, А.К. (2002). «Энторинальные аксоны демонстрируют прорастание в подполе CA1 взрослого гиппокампа в модели крыс эпилепсии височной доли». Гиппокамп . 12 (4): 534–42. doi : 10.1002/hipo.10031 . PMID 12201638 . S2CID 24965222 .
- ^ Deller, T; Адельманн, G; Nitsch, r; Frotscher, M (декабрь 1996 г.). «Путь Альвира крысиного гиппокампа». Клеточные и тканевые исследования . 286 (3): 293–303. doi : 10.1007/s004410050699 . PMID 8929332 . S2CID 36438302 .
- ^ Харрис, E; Стюарт, М (23 марта 2001 г.). «Распространение синхронных эпилептиформных событий из субикулума назад в область CA1 срезов мозга крысы». Исследование мозга . 895 (1–2): 41–9. doi : 10.1016/s0006-8993 (01) 02023-6 . PMID 11259758 . S2CID 23300272 .
- ^ Гуляс, ИИ; Görcs, TJ; Freund, TF (1990). «Иннервация различных пептидсодержащих нейронов в гиппокампе ГАМКергическими афферентами септации». Нейробиология . 37 (1): 31–44. doi : 10.1016/0306-4522 (90) 90189-b . PMID 1978740 . S2CID 24486668 .
- ^ Leäo, Rn; Микулович, S; Ле, Ке; Мунгуба, ч; Gezelius, H; Enjin, а; Patra, K; Эрикссон, а; Loew, LM; Деликт, ab; Kullander, K (ноябрь 2012 г.). «Интернейроны OLM дифференциально модулируют входы CA3 и энторинала в нейроны CA1 гиппокампа» . Nature Neuroscience . 15 (11): 1524–30. doi : 10.1038/nn.3235 . PMC 3483451 . PMID 23042082 .
- ^ Шарфман, он (25 июня 1993 г.). «Активация зубчатых нейронов Хилара путем стимуляции фимбрии в срезах гиппокампа крысы» . Нейробиологические буквы . 156 (1–2): 61–6. doi : 10.1016/0304-3940 (93) 90440-v . PMC 3281807 . PMID 8105429 .
- ^ Ян, Санггу; Ян, Сунгхил; Морейра, Тайс; Хоффман, Глория; Карлсон, Грег С.; Бендер, Кевин Дж.; Alger, Bradley E.; Тан, Ча-Мин (2014-09-02). «Межламеллярная сеть CA1 в гиппокампе» . Труды Национальной академии наук . 111 (35): 12919–12924. Bibcode : 2014pnas..11112919Y . doi : 10.1073/pnas.1405468111 . ISSN 0027-8424 . PMC 4156755 . PMID 25139992 .
- ^ Hunsaker, Michael R.; Кеснер, Рэймонд П. (2013-01-01). «Работа процессов разделения шаблонов и завершения шаблона, связанных с различными атрибутами или областями памяти». Нейробиологии и биобеводовые обзоры . 37 (1): 36–58. doi : 10.1016/j.neubiorev.2012.09.014 . PMID 23043857 . S2CID 22493885 .
- ^ Swanson, LW; Wyss, JM; Коуэн, Wm (15 октября 1978 г.). «Авторадиографическое исследование организации путей интрагиппокампальной ассоциации у крыс». Журнал сравнительной неврологии . 181 (4): 681–715. doi : 10.1002/cne.901810402 . PMID 690280 . S2CID 30954240 .
- ^ Амарал, DG; Witter, MP (1989). «Трехмерная организация формирования гиппокампа: обзор анатомических данных». Нейробиология . 31 (3): 571–91. doi : 10.1016/0306-4522 (89) 90424-7 . PMID 2687721 . S2CID 28430607 .
- ^ Кохара, К; Pignatelli, M; Rivest, AJ; Юнг, Хи; Lookmura, t; Suh, J; Фрэнк, D; Каджикава, K; Миза, н; Обата, y; Уикершхм, Ир; Tonegawa, S (февраль 2014 г.). «Генетические и оптогенные инструменты, специфичные для типа клеток, показывают цепи CA2 гиппокампа» (PDF ) Nature Neuroscience 17 (2): 269–7 Doi : 10.1038/ nnn.3 4004172PMC PMID 2436151
- ^ Blackstad, TW (октябрь 1956 г.). «Коммиссуральные соединения области гиппокампа у крысы, с особой ссылкой на их способ завершения». Журнал сравнительной неврологии . 105 (3): 417–537. doi : 10.1002/cne.901050305 . PMID 13385382 . S2CID 41672064 .
- ^ Фрике, R; Коуэн, Wm (15 сентября 1978 г.). «Авторадиографическое исследование комиссуральных и ипсилатеральных проекций гиппокампо у взрослой крысы». Журнал сравнительной неврологии . 181 (2): 253–69. doi : 10.1002/cne.901810204 . PMID 567658 . S2CID 46320248 .
- ^ Амарал, DG; Шарфман, он; Lavenex, P (2007). «Зубчатая извилина: фундаментальная нейроанатомическая организация (зубчатая извилина для манекенов)». Зубчатая извилина: всеобъемлющее руководство по структуре, функции и клиническим последствиям . Прогресс в исследовании мозга. Тол. 163. С. 3–22. doi : 10.1016/s0079-6123 (07) 63001-5 . ISBN 9780444530158 Полем PMC 2492885 . PMID 17765709 .
- ^ Рибак, CE; Сересса, L; Петерсон, GM; Сероги, KB; Фаллон, JH; Schmued, LC (декабрь 1986 г.). «ГАМКергический ингибирующий компонент в пределах комиссурального пути гиппокампа» . Журнал нейробиологии . 6 (12): 3492–8. doi : 10.1523/jneurosci.06-12-03492.1986 . PMC 6568657 . PMID 2432200 .
- ^ Сик, Аттила; Пенттонен, Маркку; Бузсаки, Гирги (март 1997 г.). «Интернейроны в гиппокампальной зубчатой извилине: внутриклеточное исследование in vivo». Европейский журнал нейробиологии . 9 (3): 573–588. doi : 10.1111/j.1460-9568.1997.tb01634.x . PMID 9104599 . S2CID 25960013 .
- ^ Jump up to: а беременный Андерсен, за; и др. (2007). Книга гиппокампа . Издательство Оксфордского университета.
- ^ Blackstad, TW (1956). «Комиссуральные соединения региона гиппокампа у крыс, со специальной ссылкой на их режим термины». J Comp Neurol . 105 (3): 417–537. doi : 10.1002/cne.901050305 . PMID 13385382 . S2CID 41672064 .
- ^ Амарал, Д.Г. (1978). «Исследование типов клеток Гольджи в области гиппокампа у крыс». J Comp Neurol . 182 (5): 851–914. doi : 10.1002/cne.901820508 . PMID 730852 . S2CID 44257239 .
- ^ "Гиппокампум" . hippocampome.org .
Внешние ссылки
[ редактировать ]- Схематическая схема срезов мозга гиппокампа
- Гиппокамп в Национальной библиотеке медицинской библиотеки США медицинские заголовки (Mesh)
- Окрашенные изображения срезов мозга, которые включают в себя «гиппокамп» в проекте Brainmaps
- Анатомия и связь гиппокампа