Губернатор Симарубы
| Губернатор Симарубы | |
|---|---|

| |
| Навес Симаруба амара | |
| Научная классификация | |
| Королевство: | Растения |
| Клэйд : | Трахеофиты |
| Клэйд : | покрытосеменные растения |
| Клэйд : | Эвдикоты |
| Клэйд : | Розиды |
| Заказ: | Сапиндейлс |
| Семья: | Симарубовые |
| Род: | Симаруба |
| Разновидность: | С. Амара
|
| Биномиальное имя | |
| Губернатор Симарубы | |
| Подвиды [ 2 ] | |
| Синонимы | |
Симаруба амара — вид дерева семейства Simaroubaceae , произрастающий в тропических лесах и саваннах Южной Центральной и Америки и Карибского бассейна . Впервые он был описан Облом . во Французской Гвиане в 1775 году и является одним из шести видов Симарубы . Дерево вечнозеленое , но раз в год дает новый набор листьев. Для роста ему требуется относительно высокий уровень света, и он быстро растет в этих условиях, но живет относительно короткое время. В Панаме он цветет в засушливый сезон в феврале и марте, тогда как в Коста-Рике, где нет засушливого сезона, цветет позже, с марта по июль. Поскольку этот вид раздельнополый , деревья бывают мужскими или женскими и дают только мужские или женские цветки. Считается, что маленькие желтые цветки опыляются насекомыми , полученные плоды разносятся животными, в том числе обезьянами, птицами и летучими мышами, питающимися фруктами, а семена также разносятся муравьями-листорезами .
Симаруба амара широко изучалась учеными в попытке понять дерево, а также лучше понять экологию тропического леса в целом. Многие из этих исследований проводились на острове Барро-Колорадо в Панаме или на биологической станции Ла-Сельва в Коста-Рике . Особый интерес представляет то, как он конкурирует с другими видами и с особями того же вида на разных стадиях своего жизненного цикла . Саженцы обычно ограничены количеством света и питательных веществ, содержащихся там, где они растут, и саженцы считаются относительно светолюбивыми по сравнению с другими видами. Молодые особи с большей вероятностью выживут, если они растут дальше от своих родителей и когда рядом с ними растет мало других особей, что может быть связано с тем, что они могут избежать болезней. Физиологи растений исследовали, как листья деревьев различаются в зависимости от их расположения в пологе леса, обнаружив, что они толще в пологе и тоньше в подлеске. Они также измерили, насколько изменяется водный потенциал их листьев и время открытия и закрытия устьиц в течение дня; Результаты показывают, что вместо того, чтобы закрывать устьица, чтобы контролировать потерю воды, вместо этого она контролируется площадью листа. Популяционные генетики изучили, как варьируются его гены , как в местном масштабе, так и по всему ареалу , используя микросателлиты . Он генетически разнообразен, что указывает на то, что между популяциями происходит поток генов , а семена могут распространяться на расстояние до 1 км. Листья S. amara поедаются несколькими видами гусениц , особенно рода Atteva . несколько видов термитов и муравьев На дереве или вокруг него обитают лианы и эпифиты , а на дереве растут .
Кора S. amara использовалась людьми, живущими в ее ареале, для лечения дизентерии и диареи , а также других заболеваний, а также экспортировалась в Европу в восемнадцатом веке для лечения этих болезней. С тех пор из коры был выделен ряд соединений, которые, как было доказано, обладают противомикробным действием. Местные жители используют древесину дерева для различных целей, ее также выращивают на плантациях и заготавливают древесину, часть которой идет на экспорт.
Описание
[ редактировать ]Симаруба амара вырастает до высоты 35 метров, с максимальным диаметром ствола 125 см и максимальным предполагаемым возрастом 121 год. [ 5 ] [ 6 ] [ 7 ] У него сложные листья длиной около 60 см каждый, черешки длиной 4–7 см, каждый лист имеет 9–16 листочков. Каждый листочек имеет длину 2,5–11 см и ширину 12–45 мм, причем те, что ближе к концу сложного листа, имеют тенденцию быть меньше. Цветки расположены на тычиночной метелке длиной около 30 см, широко разветвленной и густо покрытой цветками. [ 8 ] Цветки однополые, мелкие (длиной менее 1 см), бледно-желтого цвета. Считается, что они опыляются насекомыми, такими как маленькие пчелы и мотыльки . На острове Барро-Колорадо (BCI), Панама, он, как правило, цветет в засушливый сезон с конца января до конца апреля, продолжаясь от 11 до 15 недель каждый год. [ 5 ] В Коста-Рике цветет немного позже, с марта по июль, достигая пика в апреле. [ 9 ] Плоды формируются через 1–3 месяца после опыления. Плоды от ярко-зеленого до пурпурно-черного цвета, длиной около 17 мм, содержат крупные семена (10–14 мм), собраны группами по 3–5 костянок . Семена не могут оставаться в состоянии покоя и распространяются позвоночными животными. [ 5 ] Каждое семя весит примерно 0,25 г. [ 6 ]
Это вечнозеленый вид, новая волна листьев которого растет в период с января по апрель, в засушливый сезон, когда в тропических лесах наблюдается самый высокий уровень освещенности. такая фенология Считается, что позволяет S. amara фотосинтезировать наиболее эффективно, поскольку новые листья более эффективны, чем те, которые они заменяют. [ 7 ] [ 9 ] Имеет видимые, но нечеткие годичные кольца шириной в среднем 7 мм. [ 7 ] Исследование людей в Панаме показало, что они вырастают в среднем на 8,4 мм в диаметре каждый год. [ 10 ] в Коста-Рике зафиксированы темпы роста до 18 мм в год, при этом стебель растет постоянно в течение года. [ 9 ] Сосуды ксилемы у взрослых деревьев имеют диаметр от 20 до 90 мкм, при этом на мм приходится около 50 сосудов. 2 филиала. [ 11 ] Плотность . древесины 0,37–0,44 г/см 3 , [ 6 ] ниже, чем у многих других видов тропических лесов. [ 12 ]
Это быстрорастущий, светолюбивый и теневыносливый вид. Саженцы обычно представляют собой один прямой полюс с несколькими сложными листьями и только одной точкой роста. Это позволяет саженцу добиться наибольшего вертикального роста при минимальном количестве биомассы. [ 13 ] Они начинают ветвиться, когда достигают высоты 2–5 м. [ 14 ]
Исследование графика динамики леса на BCI показало, что в период с 1982 по 2000 год около 65% особей умерли, при этом самая высокая смертность среди мелких особей (<1 см dbh ). Большие деревья (> 20 см глубины) встречаются относительно редко, в среднем 2,4 дерева на гектар по сравнению с 40 деревьями на гектар с глубиной > 1 см. [ 5 ]
Таксономия
[ редактировать ]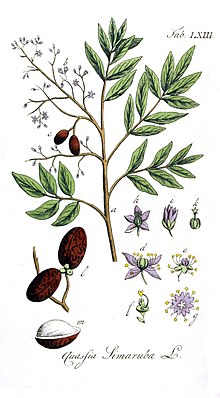
Симаруба амара была впервые описана Жаном Батистом Кристофором Фюзе Обле во Французской Гвиане в 1775 году и является типовым видом рода Симаруба . [ 1 ] [ 2 ] В 1790 году Уильям Райт описал Quassia simarouba , [ 15 ] который, как предположил Огюст Пирам Декандоль, принадлежал к тому же виду, что и S. amara . Однако, поскольку S. amara Обле описал как однодомный, а Райт Q. simarouba описал как раздельнополый, в 1829 году они все еще считались отдельными видами. [ 16 ] К 1874 году, когда была опубликована Flora Brasiliensis , они считались синонимами. [ 17 ]
Среди шести видов симарубы , помимо S. amara, на континенте встречаются еще два: S. glauca и S. versicolor . S. amara можно отличить от других континентальных видов по меньшим цветкам, пыльникам и плодам, а также по прямым, а не изогнутым лепесткам. [ 18 ] Листья Simarouba amara subsp. opaca не имеют сизоватого цвета (голубовато-серого или зеленого цвета) на нижней стороне, тогда как у Simarouba amara subsp. типичные . [ 2 ]
| Структура | С. Амара | С. глаука | С. разноцветный |
|---|---|---|---|
| Цветок | 3–5 мм в длину | Длина 4–7,5 мм | Длина 4–7,5 мм |
| Пыльники | Длина 0,4–1,2 мм | Длина 1,3–2,0 мм | Длина 1,0–1,5 мм |
| Лепестки | Прямые, тусклые от желто-зеленого до белого цвета. | Изогнутый, ярко-желтый с оттенком оранжевого или красного. | изогнутый |
| Фрукты | 1,0–1,5 х 0,6–1,0 см | 2,0–2,5 х 1,2–1,5 см | 2,0–2,5 х 1,5–2,0 см |
Общие имена
[ редактировать ]Симаруба амара известна под многими распространенными названиями в Неотропах. В Боливии он известен как чириуана, в Бразилии — как марупа, марупуаба, парахиба, параиба и таманкейра. В Колумбии его называют симаруба, в Эквадоре - горький кедр, колыбель и гитара, во Французской Гвиане - симаруба, в Гайане - симарупа, в Перу - марупа, в Суринаме - сомароба, а в Венесуэле - кедр бланко и симаруба. [ 19 ]
В Европе он был известен под разными названиями в девятнадцатом веке, когда его использовали в качестве лекарства; в эти имена входил горький пепел, [ 20 ] горькая древесина, [ 19 ] Маунтин-Дэмсон [ 21 ] и клепка. [ 22 ]
Распределение
[ редактировать ]
Естественный ареал S. amara находится в Неотропах , экорегионе Центральной Южной и Америки . Его ареал простирается от Гватемалы на севере до Боливии на юге и от Эквадора на западе до восточного побережья Бразилии . [ 18 ] Он был завезен на острова Доминика и Пуэрто-Рико в Карибском море и натурализовался в Пуэрто-Рико. [ 23 ] [ 24 ] На BCI взрослые деревья (>10 см на гектар) встречаются с частотой 5 на гектар, в Эквадоре - 0,7 на гектар и во Французской Гвиане - 0,4 на гектар. Генетический анализ популяций показывает, что он всегда был относительно распространен в своем ареале. [ 25 ] Растет в тропических лесах и саваннах . [ 26 ] сеянцы S. amara встречаются редко В девственном лесу из-за светолюбивости. [ 13 ]
Генетика
[ редактировать ]Популяции S. amara демонстрируют высокий уровень гетерозиготности , что указывает на их генетическое разнообразие . Это согласуется с распространением деревьев на большие расстояния в результате потока пыльцы на большие расстояния и с тем, что между популяциями существует достаточный поток генов на большие расстояния , чтобы противодействовать эффектам генетического дрейфа . Исследование 478 растений из 14 популяций Южной Америки показало, что 24% всех аллелей встречаются только в одной популяции. [ 25 ] Исследование 300 растений на острове Барро-Колорадо показало, что гетерозиготность по 5 микросателлитным локусам варьировала от 0,12 до 0,75. 8 из 50 выявленных аллелей встречались только у одного растения. [ 5 ]
Воспроизведение
[ редактировать ]Особи обычно не размножаются, пока диаметр ствола не достигнет 30 см. Повзрослев, деревья ежегодно цветут, но не все самки ежегодно приносят плоды. [ 5 ] Морфология их цветков типична для опыления мелкими насекомыми широкого профиля, такими как пчелы и мотыльки. [ 5 ] Сообщается, что его опыляют несфингидные бабочки . [ 27 ] но другие авторы сомневаются, правильно ли это. [ 28 ]
Распространение семян
[ редактировать ]
Семена S. amara распространяются позвоночными животными, преимущественно крупными птицами и млекопитающими, в том числе чачалаками , мухоловками , мотмотами , дроздами , ревунами , тамаринами. [ 29 ] и обезьяны-пауки . Также было замечено, что муравьи-листорезы рассеивают семена, и проростков . на участках, куда они сбрасывают отходы, образуются плотные ковры [ 5 ] но большая часть сеянцев погибает, и считается, что расселение муравьев не имеет значения для определения долгосрочных закономерностей пополнения и расселения. [ 30 ] Семена, которые едят обезьяны, имеют больше шансов прорасти, чем семена, которые этого не сделали. [ 31 ] питающиеся фруктами, -филлостомиды, Также было отмечено, что летучие мыши рассеивают свои семена; это может способствовать регенерации лесов, поскольку они разносят семена более поздних сукцессионных видов, питаясь S. amara . [ 32 ]
На основе обратного моделирования данных семенных ловушек на BCI расчетное среднее расстояние распространения семян составляет 39 м. [ 5 ] Изучение сеянцев и родительских деревьев на BCI с использованием микросателлитов ДНК показало, что на самом деле в среднем саженцы растут на расстоянии 392 м от своих родителей со стандартным отклонением ± 234 м и диапазоном от 9 м до 1 км. В лесу под репродуктивными самками много семян и саженцев; генетические данные показывают, что саженцы вряд ли произошли от близлежащих взрослых особей, а скорее были рассеяны там позвоночными, которые питались на одном дереве, а затем переместились, чтобы питаться на другое, испражняясь, находясь в кроне, и откладывая семена. [ 29 ]
Физиология
[ редактировать ]различные аспекты физиологии S. amara . Были изучены Устьичная воды проводимость листьев, показатель скорости испарения , у взрослых деревьев в полдень колеблется от 200 до 270 ммоль H 2 O·m. −2 с −1 . листьев Водный потенциал в полдень колеблется от -0,56 до -1,85 МПа , составляя в среднем около -1,2 МПа. Кавитация широко распространена в туловище, и устьица не закрываются до возникновения кавитации. Хотя обычно это считается вредным для дерева, оно может буферизовать потенциал воды в листьях и, следовательно, быть полезным. Устьичная проводимость и гидравлическая проводимость ветвей более высоких деревьев (~30 м) значительно выше, чем у ветвей более мелких деревьев (~20 м). Филлипс, Бонд и Райан предположили, что это, вероятно, связано с тем, что у ветвей более высоких деревьев соотношение листа и заболони ниже, чем у мелких ветвей. Окрашивание красителем показывает, что в ветвях S. amara часто встречается кавитация . Они пришли к выводу, что поток воды у S. amara контролируется структурными (площадь листьев), а не физиологическими (закрытие устьиц) способами. [ 11 ]
Листья поглощают свет в фотосинтетически активного излучения (ФАР) спектре на длинах волн от 400 до 700 нм с высокой эффективностью, но эффективность снижается при более длинных волнах. Обычно растения поглощают ФАР с эффективностью около 85%; Считается, что более высокие значения, обнаруженные у S. amara, обусловлены высокой влажностью среды его обитания. Коэффициент отражения и пропускания листьев низок в диапазоне от 400 до 700 нм. Оптические свойства и масса листьев варьируются в зависимости от их местоположения в пологе леса , при этом листья становятся толще и эффективнее по мере увеличения их высоты внутри полога. Однако из-за своего веса листья в подлеске более эффективно улавливают свет, чем листья в кроне. [ 33 ]
| Поглощение (400-700 нм) | Поглощение (700-750 нм) | Отражательная способность (400-700 нм) | Пропускание (400-700 нм) | Масса листа (г/м 2 ) | |
|---|---|---|---|---|---|
| Подлесок (~2 м) | 91.7% | 37.3% | 6.3% | 2.1% | 36.9 |
| Средний навес (~10 м) | 92.8% | 41.6% | 6.1% | 1.2% | 55.4 |
| Навес (>20 м) | 93.1% | 46.1% | 5.2% | 0.5% | 135.2 |
концентрация биодоступного фосфата Было обнаружено, что под самками выше, чем под самцами, хотя общая концентрация фосфатов одинакова. Роудс и др. пришли к выводу, что эта разница связана с тем, что самки меняют доступность фосфатов, а не самки растут только в районах с высокой доступностью фосфатов. Считается, что это вызвано либо фруктами, содержащими высокий уровень фосфатов, которые могут упасть с дерева и сгнить, либо фруктами, привлекающими животных, которые откладывают фосфат под самками. Также возможно, что представители разных полов производят разные корневые экссудаты, которые влияют на микробное сообщество в их ризосфере , тем самым влияя на доступность фосфатов. [ 34 ]
древесные ткани S. amara Было обнаружено, что дышат со скоростью 1,24 мкмоль CO 2 м. −2 с −1 , и эта скорость дыхания положительно коррелирует со скоростью роста стебля. Поддерживающее дыхание рассчитывали на уровне 31,1 мкмоль CO 2 м. −3 с −1 и эта скорость положительно коррелировала с объемом заболони . [ 35 ]
Физиология рассады
[ редактировать ]
| Питательное вещество | Концентрация (мг/г листа)
(сухой вес) |
|---|---|
| Азот | 20 |
| Фосфор | 1 |
| Калий | 16 |
| Кальций | 2 |
| Магний | 5 |
выкапывали траншеи Эксперименты с BCI, когда вокруг саженцев S. amara или над ними делали промежутки в кроне, показывают, что их относительная скорость роста может быть увеличена обоими способами. Это показывает, что их рост обычно ограничивается как надземной конкуренцией за свет, так и подземной конкуренцией за питательные вещества и воду. Конкуренция за свет обычно более важна, о чем свидетельствует увеличение скорости роста почти в 7 раз и снижение смертности при размещении сеянцев в промежутках по сравнению с подлеском. Когда вокруг рассады в промежутках выкапывали траншею, чтобы предотвратить подземную конкуренцию, их рост еще больше увеличивался, на 50%, демонстрируя, что в просветах сеянцы ограничены подземной конкуренцией. Выкапывание траншей вокруг саженцев в подлеске не привело к значительному увеличению их роста, показывая, что обычно они ограничиваются только конкуренцией за свет. [ 36 ]
Более крупные саженцы с большей вероятностью переживут засушливый сезон при использовании BCI, чем более мелкие. [ 37 ] Между сеянцами происходит ингибирование, зависящее от плотности : они с большей вероятностью выживут на участках, где меньше сеянцев S. amara растет . Исследование людей на BCI показало, что эта закономерность может быть вызвана различиями в биоте почвы, а не травоядными насекомыми или грибковыми патогенами. [ 37 ] Наблюдения, основанные на расстоянии сеянцев от их родителей, показывают, что гипотеза Янцена-Коннелла применима к сеянцам S. amara : они с большей вероятностью выживут вдали от своих родителей, поскольку избегают вредителей, таких как травоядные животные , и патогенов растений , которые чаще встречаются под землей. родительские деревья. [ 29 ]
Физиология саженца
[ редактировать ]Саженцы S. amara требовательны к свету и встречаются в более ярких участках тропического леса по сравнению с саженцами Pitheullobium elegans и Lecythis ampla . Исследование, проведенное на биологической станции Ла-Сельва, показало, что листья весят примерно 30 г/м. 2 (сухой вес), похож на P. elegans , но примерно в два раза тяжелее L. ampla . Фотосинтетическая способность листьев S. amara выше, чем у двух других видов, и составляет в среднем около 6 мкмоль · м. −1 с −1 . Темновое дыхание составляет в среднем 0,72 мкмоль·м. −1 с −1 , выше, чем у двух других видов. Максимальная скорость фотосинтеза коррелирует как с диаметром стебля, так и с вертикальным ростом. Считается, что рассеянный свет более важен для роста рассады, чем солнечные блики . [ 13 ] Другое исследование саженцев в Ла-Сельве показало, что они вырастают на 7 см в год. −1 по высоте и 0,25 мм год −1 в диаметре. В среднем у них было девять сложных листьев, индекс площади листьев составлял 0,54, а общая площадь поверхности листьев составляла 124 см2. 2 . Саженцы с наименьшей площадью листьев чаще всего погибали во время исследования, а саженцы с большей площадью листьев росли быстрее, чем другие саженцы. [ 14 ]
Исследование саженцев диаметром от одного до четырех сантиметров на BCI показало, что рост саженцев не менялся в зависимости от того, какие виды росли рядом с ними, вопреки предсказаниям о том, что происходит торможение в зависимости от плотности. Модель, основанная на этих результатах, предсказала, что саженцы диаметром 2 см способны расти с максимальной скоростью 13 мм в год. −1 и что если в пределах 5 м от саженца растет другое дерево диаметром 10 см, его рост сокращается всего до 12 мм в год. −1 , что указывает на то, что скученность на них не влияет. Деревья, растущие на расстоянии более 15 м от саженца, не влияют на их рост. [ 38 ]
Экология
[ редактировать ]
Лианы относительно редки на взрослых (> 20 см по высоте) особях S. amara по сравнению с другими деревьями на BCI, и только около 25% из них имеют лианы, растущие на них. Путц предположил, что это может быть связано с тем, что деревья имеют большие листья, но механизм, с помощью которого это приведет к уменьшению количества лиан, неизвестен. [ 10 ] Более мелкие особи также имеют меньше лиан и древесных гемиэпифитов, чем другие виды деревьев в тех же лесах. [ 39 ]
Было зарегистрировано, что паутинный червь айлант ( Atteva aurea ) и другие представители рода Atteva поедают кончики новых побегов S. amara в Коста-Рике. [ 40 ] Личинки вида бабочек Bungalotis diophorus питаются исключительно саженцами и побегами S. amara . [ 41 ] Два вида термитов были замечены живущими на S. amara в Панаме: Calcaritermes brevicollis в валежной древесине и Microcerotermes arboreus, гнездящимися в галерее на ветке. [ 42 ] Было обнаружено, что муравьи-пули ( Paraponera clavata ) гнездятся у подножия деревьев S. amara . [ 43 ] Полужесткокрылые ( Fulgoridae Enchophora sanguinea . amara ) обнаруживаются преимущественно на S. стволах [ 44 ] [ 45 ]
Использование
[ редактировать ]Материалы
[ редактировать ]Симаруба амара используется на местном уровне для производства бумаги, мебели, фанеры и спичек, а также в строительстве. [ 5 ] Его также выращивают на плантациях, поскольку его яркая и легкая древесина пользуется большим спросом на европейских рынках для изготовления изысканной мебели и шпона . [ 6 ] [ 46 ] Древесина быстро сохнет, и с ней легко работать обычными инструментами. Цвет от кремово-белого до светло-желтого, с грубой текстурой и прямыми волокнами. Его необходимо обрабатывать, чтобы предотвратить грибками , древоточцами и термитами его поедание . Ядро . имеет плотность 0,35–0,45 г/см 3 . [ 19 ] Было отмечено, что это один из лучших видов древесины, который можно выращивать в перуанской Амазонке , наряду с Cedrelinga catenaeformis , из-за его характеристик быстрого роста. [ 47 ] Всемирный фонд природы рекомендует потребителям убедиться, что древесина S. amara сертифицирована Лесным попечительским советом , чтобы она не способствовала вырубке лесов . [ 48 ] Древесные стружки S. amara использовались в качестве подстилки для животных, что приводило к отравлению лошадей и собак. [ 49 ]
Медицинский
[ редактировать ]Этот раздел нуждается в более надежных медицинских справках для проверки или слишком сильно полагается на первоисточники . ( январь 2017 г. ) |  |
Листья и кора S. amara используются в качестве лекарственного растения для лечения дизентерии , диареи , малярии и других заболеваний в районах, где он растет. [ нужна ссылка ] В 1713 году его экспортировали во Францию, где его использовали для лечения дизентерии, будучи эффективным средством лечения во время эпидемий между 1718 и 1725 годами. В 1918 году его эффективность была подтверждена исследованием, в ходе которого солдатам в военном госпитале давали чай из коры. для лечения амебной дизентерии . [ нужна медицинская ссылка ] В исследовании 1944 года Институт Мерк обнаружил, что его эффективность при лечении кишечного амебиаза у людей составляет 92%. [ нужна медицинская ссылка ] В 1990-х годах ученые продемонстрировали, что он может убить наиболее распространенную причину дизентерии — Entamoeba histolytica , а также виды бактерий Salmonella и Shigella , вызывающие диарею. [ 50 ] большее количество доказательств Для доказательства эффективности Симарубы требуется .
Основными биологически активными соединениями, обнаруженными в S. amara, являются квассиноиды — группа тритерпенов , включающая илалантинон , глаукарубинон и холакантон . Сообщалось, что они убивают простейших, амебу и плазмодий (причину малярии). Противомалярийные свойства были впервые исследованы учеными в 1947 году; они обнаружили, что у кур 1 мг экстракта коры на 1 кг массы тела обладает сильной противомалярийной активностью. [ нужна медицинская ссылка ] В 1997 году в США был подан патент на использование экстракта в средствах по уходу за кожей. [ 50 ] Симаруба амара не следует путать с Симаруба глаука , которая в Индии известна как Лакшми Тару .
Ссылки
[ редактировать ]- ^ Jump up to: а б Кристофор Фюзе Обле, Жан Батист (1775). История растений Гвианы Франсуаза . Лондон и Париж: Пьер-Франсуа Дидо. стр. 860–861.
- ^ Jump up to: а б с Кронквист, А. (1944). «Исследования Simaroubaceae-II. Род Simarouba». Бюллетень Ботанического клуба Торри . 71 (3): 226–234. дои : 10.2307/2481702 . JSTOR 2481702 .
- ^ Jump up to: а б с Ботанический сад Миссури. «Синонимы Симаруба амара » . tropicos.org . Проверено 12 января 2011 г.
- ^ Герберт Стоун (1904). «Деревья коммерции и их идентификация» . Природа . 71 (1837): 247. Бибкод : 1905Natur..71..247. . дои : 10.1038/071247b0 . hdl : 2027/gri.ark:/13960/t18m0qr47 . S2CID 34933195 . Проверено 13 января 2011 г.
- ^ Jump up to: а б с д и ж г час я дж Хардести, Б.; Дик, К.; Кремер, А.; Хаббелл, С.; Бермингем, Э. (2005). «Пространственная генетическая структура Simarouba amara Aubl. (Simaroubaceae), раздельнополого неотропического дерева, расселенного среди животных, на острове Барро-Колорадо, Панама» . Наследственность . 95 (4): 290–297. дои : 10.1038/sj.hdy.6800714 . hdl : 2027.42/83301 . ПМИД 16094303 . S2CID 25442574 .
- ^ Jump up to: а б с д Шульце, М.; Гроган, Дж.; Лэндис, Р.; Видаль, Э. (2008). «Насколько редки или слишком редки, чтобы их можно было собирать? Проблемы управления, связанные с древесными породами, встречающимися с низкой плотностью в бразильской Амазонии». Лесная экология и управление . 256 (7): 1443. doi : 10.1016/j.foreco.2008.02.051 .
- ^ Jump up to: а б с Фихтлер, Э.; Кларк, округ Колумбия; Ворбс, М. (2003). «Возраст и долгосрочный рост деревьев в старовозрастном тропическом лесу, на основе анализа годичных колец и 14 C". Biotropica . 35 (3): 306–317. doi : 10.1111/ . JSTOR 30043047. . S2CID 86366029 j.1744-7429.2003.tb00585.x
- ^ Вудсон, Р.Э.; Шери, RW; Портер, DM (1973). «Флора Панамы. Часть VI. Семейство 90. Simaroubaceae». Анналы ботанического сада Миссури . 60 (1): 23–39. дои : 10.2307/2394767 . JSTOR 2394767 .
- ^ Jump up to: а б с О'Брайен, Джей-Джей; Обербауэр, Сан-Франциско; Кларк, Д.Б.; Кларк, Д.А. (2007). «Фенология и сезонность увеличения диаметра стебля во влажном тропическом лесу Коста-Рики». Биотропика . 40 (2): 151. doi : 10.1111/j.1744-7429.2007.00354.x . S2CID 10787182 .
- ^ Jump up to: а б Путц, Ф.Е. (1984). «Как деревья избегают лиан и сбрасывают их». Биотропика . 16 (1): 19–23. Бибкод : 1984Биотр..16...19П . дои : 10.2307/2387889 . JSTOR 2387889 .
- ^ Jump up to: а б Филлипс, Н.; Бонд, Би Джей; Райан, МГ (2001). «Газообмен и гидравлические свойства крон двух пород деревьев во влажном лесу Панамы» (PDF) . Деревья . 15 (2): 123–130. Бибкод : 2001Деревья..15..123P . дои : 10.1007/s004680000077 . S2CID 420536 . Архивировано из оригинала (PDF) 9 июня 2010 г. Проверено 2 августа 2010 г.
- ^ Сантьяго, Л.; Гольдштейн, Г.; Мейнцер, Ф.; Фишер, Дж.; МакХадо, К.; Вудрафф, Д.; Джонс, Т. (2004). «Фотосинтетические характеристики листьев зависят от гидравлической проводимости и плотности древесины в пологах панамских лесов». Экология . 140 (4): 543–550. Бибкод : 2004Oecol.140..543S . дои : 10.1007/s00442-004-1624-1 . ПМИД 15232729 . S2CID 4981403 .
- ^ Jump up to: а б с Обербауэр, Сан-Франциско; Кларк, Д.Б.; Кларк, округ Колумбия; Рич, премьер-министр; Вега, Г. (2009). «Световая среда, газообмен и ежегодный рост саженцев трех видов деревьев тропических лесов Коста-Рики». Журнал тропической экологии . 9 (4): 511–523. дои : 10.1017/S0266467400007586 . S2CID 85727139 .
- ^ Jump up to: а б Стерк, Ф.Дж.; Кларк, Д.Б.; Кларк, округ Колумбия; Бонгерс, Ф. (1999). «Колебания света, особенности кроны и задержки реакции саженцев деревьев в низменных тропических лесах Коста-Рики» . Журнал тропической экологии . 15 (1): 83–95. дои : 10.1017/S0266467499000681 . JSTOR 2560198 . S2CID 10434491 .
- ^ «Квасия симаруба» . Zipcodezoo.com . Проверено 10 января 2011 г.
- ^ Эндрю Дункан (1829). Дополнение к новой амбулатории в Эдинбурге . Напечатано для Bell & Bradfute. стр. 85–86 .
- ^ Нолан, Эдв. Дж; Джонс, Олив (31 декабря 2010 г.). «Флора бразильская» . Наука . 35 (909): 864–5. Бибкод : 1912Sci....35..864N . дои : 10.1126/science.35.909.864 . ПМИД 17820223 . S2CID 5174658 . Проверено 4 января 2011 г.
- ^ Jump up to: а б с Франческинелли, EV; Ямамото, К.; Шеперд, Дж.Дж. (1998). «Различия между тремя видами симарубы». Систематическая ботаника . 23 (4): 479–488. дои : 10.2307/2419379 . JSTOR 2419379 .
- ^ Jump up to: а б с «Тропикс 6.0 – Марупа» (PDF) . ТРОПИКС ЦИРАД. 2 марта 2009 г. Архивировано из оригинала (PDF) 2 июля 2011 г. Проверено 1 января 2011 г.
- ^ Ривьер, Питер (2006). Путешествие Роберта Шомбургка по Гвиане, 1835–1844, Том 17 . Издательство Эшгейт. п. 34. ISBN 978-0-904180-88-6 .
- ^ Роберт Эглсфельд Гриффит (1847). Медицинская ботаника . Леа и Бланшар. стр. 198 – . Проверено 10 января 2011 г.
- ^ АХР Гризебах (1864 г.). Флора островов Британской Вест-Индии . Ловелл Рив и Ко. р. 788 . Проверено 10 января 2011 г.
- ^ Мозес Кайро, Биби Али, Оливер Чизман, Карен Хейсом и Шон Мерфи (2003). «Угрозы инвазивных видов в Карибском регионе» (PDF) . КАБИ. п. 122. Архивировано из оригинала (PDF) 1 октября 2016 г. Проверено 10 января 2011 г.
{{cite web}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ «Инвазивные и интродуцированные виды деревьев на Доминике» . Продовольственная и сельскохозяйственная организация Объединенных Наций. 01 апреля 2009 г. Проверено 14 января 2011 г. [ постоянная мертвая ссылка ]
- ^ Jump up to: а б Хардести, Б.Д.; Дик, CW; Хэмрик, Дж.Л.; Деген, Б.; Хаббелл, СП; Бермингем, Э. (2010). «Географическое влияние на генетическую структуру широко распространенного неотропического дерева Симаруба амара (Simaroubaceae)». Биология тропических растений . 3 : 28–39. дои : 10.1007/s12042-010-9044-3 . hdl : 2027.42/83290 . S2CID 10722698 .
- ^ Санайотти, ТМ; Магнуссон, МЫ (1995). «Влияние ежегодных пожаров на производство мясистых фруктов, поедаемых птицами в бразильской амазонской саванне». Журнал тропической экологии . 11 (1): 53–65. дои : 10.1017/S0266467400008397 . JSTOR 2560140 . S2CID 84023140 .
- ^ Реннер, СС; Фейл, JP (1993). «Опылители тропических двудомных покрытосеменных» . Американский журнал ботаники . 80 (9): 1100–1107. дои : 10.2307/2445757 . JSTOR 2445757 .
- ^ Бава, Канзас (1994). «Опылители тропических двудомных покрытосеменных: переоценка? Нет, еще нет». Американский журнал ботаники . 81 (4): 456–460. дои : 10.2307/2445495 . JSTOR 2445495 .
- ^ Jump up to: а б с Хардести, Б.Д.; Хаббелл, СП; Бермингем, Э. (2006). «Генетические свидетельства частого пополнения на большие расстояния среди деревьев, расселённых по позвоночным». Экологические письма . 9 (5): 516–525. дои : 10.1111/j.1461-0248.2006.00897.x . ПМИД 16643297 .
- ^ Хардести, Б.Д. (2011). «Эффективность распространения семян муравьями на неотропическом дереве». Интегративная зоология . 6 (3): 222–6. дои : 10.1111/j.1749-4877.2011.00246.x . ПМИД 21910841 .
- ^ Стивенсон, PR; Кастелланос, MC; Писарро, Дж. К.; Гаравито, М. (2002). «Влияние распространения семян тремя видами обезьян-ателинов на прорастание семян в национальном парке Тинигуа, Колумбия». Международный журнал приматологии . 23 (6): 1187. doi : 10.1023/A:1021118618936 . S2CID 5560227 .
- ^ Кельм, Д. .; Визнер, К. .; Фон Хельверсен, О. . (2008). «Влияние искусственных насестов плодоядных летучих мышей на распространение семян на мозаике неотропических лесных пастбищ». Биология сохранения . 22 (3): 733–741. Бибкод : 2008ConBi..22..733K . дои : 10.1111/j.1523-1739.2008.00925.x . ПМИД 18445078 . S2CID 23047687 .
- ^ Jump up to: а б Пуртер, Л.; Обербауэр, Сан-Франциско; Кларк, Д.Б. (1995). «Оптические свойства листьев вдоль вертикального градиента в пологе тропического леса в Коста-Рике». Американский журнал ботаники . 82 (10): 1257–1263. дои : 10.2307/2446248 . JSTOR 2446248 .
- ^ Роудс, CC; Сэнфорд, РЛ; Кларк, Д.Б. (1994). «Гендерзависимое влияние на почвенный фосфор двудомного низинного тропического дерева Симаруба амара» (PDF) . Биотропика . 26 (4): 362–368. Бибкод : 1994Биотр..26..362Р . дои : 10.2307/2389229 . JSTOR 2389229 .
- ^ Райан, Миннесота; Хаббард, РМ; Кларк, округ Колумбия; Сэнфорд, РЛ (1994). «Дыхание древесных тканей Simarouba amara и Minquartia guianensis, двух тропических влажных лесных деревьев с разными особенностями роста» (PDF) . Экология . 100 (3): 213–220. Бибкод : 1994Oecol.100..213R . дои : 10.1007/BF00316947 . ПМИД 28307003 . S2CID 2296105 . [ постоянная мертвая ссылка ]
- ^ Jump up to: а б Барберис, И.М.; Таннер, EVJ (2005). «Зазоры и корневые траншеи увеличивают рост саженцев деревьев в полувечнозеленых лесах Панамы». Экология . 86 (3): 667–674. Бибкод : 2005Ecol...86..667B . дои : 10.1890/04-0677 . JSTOR 3450661 . S2CID 84821595 .
- ^ Jump up to: а б Манган, С.; Шнитцер, С.; Эрре, Э.; Мак, К.; Валенсия, М.; Санчес, Э.; Бевер, Дж. (2010). «Отрицательная обратная связь между растениями и почвой предсказывает относительное изобилие видов деревьев в тропическом лесу» . Природа . 466 (7307): 752–755. Бибкод : 2010Natur.466..752M . дои : 10.1038/nature09273 . ПМИД 20581819 . S2CID 4327725 .
- ^ Уриарте, Массачусетс; Кондит, Р.; Кэнхэм, CD; Хаббелл, СП (2004). «Пространственно явная модель роста саженцев в тропическом лесу: имеет ли значение идентичность соседей?» . Журнал экологии . 92 (2): 348–360. Бибкод : 2004JEcol..92..348U . дои : 10.1111/j.0022-0477.2004.00867.x . JSTOR 3599599 .
- ^ Кларк, Д.Б.; Кларк, Д.А. (1990). «Распространение и влияние на рост деревьев лиан и древесных гемиэпифитов в тропическом влажном лесу Коста-Рики». Журнал тропической экологии . 6 (3): 321–331. дои : 10.1017/S0266467400004570 . JSTOR 2559832 . S2CID 84035738 .
- ^ Уилсон, Дж.; Лэндри, JFO; Янзен, Д.; Хальвахс, В.; Назари, В.; Хаджибабаи, М.; Хеберт, П. (2010). «Идентификация мотылька айланта (Lepidoptera, Yponomeutidae), комплекса двух видов: данные штрих-кодирования ДНК, морфологии и экологии» . ZooKeys (46): 41–60. Бибкод : 2010ZooK...46...41W . дои : 10.3897/zookeys.46.406 .
- ^ Джефф, Миллер; Дэниел Х. Янзен; Уинифред Хальвахс (2009). Сто бабочек и мотыльков . Издательство Гарвардского университета. ISBN 978-0-674-02334-5 .
- ^ Ройзен, Ю. .; Дежан, А. .; Корбара, Б. .; Оривел, Дж. .; Саманиего, М. .; Лепонс, М. . (2006). «Вертикальная стратификация скопления термитов в неотропическом тропическом лесу» (PDF) . Экология . 149 (2): 301–311. Бибкод : 2006Oecol.149..301R . дои : 10.1007/s00442-006-0449-5 . ПМИД 16791633 . S2CID 7700084 .
- ^ Белк, MC; Блэк, ХЛ; Йоргенсен, CD; Хаббелл, СП; Фостер, РБ (1989). «Избирательность гнездового дерева тропического муравья Paraponera clavata». Биотропика . 21 (2): 173–177. Бибкод : 1989Биотр..21..173Б . дои : 10.2307/2388707 . JSTOR 2388707 .
- ^ Джонсон, ЛК; Фостер, РБ (1985). «Ассоциации крупных равнокрылых и деревьев в тропическом лесу». Журнал Канзасского энтомологического общества . 58 (3): 565. JSTOR 25084689 .
- ^ Наскрецкий, П.; Нисида, К. (2007). «Новые трофобиотические взаимодействия у клопов-фонарей (Insecta: Auchenorryncha: Fulgoridae)» (PDF) . Журнал естественной истории . 41 (37–40): 2397. Бибкод : 2007JNatH..41.2397N . дои : 10.1080/00222930701633570 . S2CID 54791767 .
- ^ Гранье, А.; Хук, Р.; Колин, Ф. (1992). «Транспирация и устьичная проводимость двух видов тропических лесов, растущих на плантациях (Simarouba amara и Goupia glabra) во Французской Гвиане» . Анналы лесных наук . 49 (1): 17. дои : 10.1051/forest:19920102 .
- ^ Макклейн, Майкл; Рейнальдо Л. Виктория; Джеффри Эдвард Ричи (2001). Биогеохимия бассейна Амазонки . Издательство Оксфордского университета. п. 111. ИСБН 978-0-19-511431-7 .
- ^ «Фотогид для идентификации вашей древесины» . Всемирный фонд природы (WWF) . Проверено 4 января 2011 г.
- ^ Деклерк, Дж. (2004). «Подозрение на отравление древесиной, вызванное стружками Симаруба амара (marupá/caixeta) у двух собак с эрозивным стоматитом и дерматитом». Ветеринарная дерматология . 15 (3): 188–193. дои : 10.1111/j.1365-3164.2004.00377.x . ПМИД 15214956 .
- ^ Jump up to: а б Виртуальный полевой гербарий. «Simaroubaceae Simarouba amara Aublet» . Гербарий Оксфордского университета. Архивировано из оригинала 18 июля 2011 г. Проверено 11 января 2011 г.