Химическая библиотека, закодированная в ДНК
Химические библиотеки, кодированные ДНК ( DECL ) — это технология синтеза и скрининга в беспрецедентных масштабах коллекций низкомолекулярных соединений. DECL используется в медицинской химии для объединения областей комбинаторной химии и молекулярной биологии . Целью технологии DECL является ускорение процесса открытия лекарств и, в частности, деятельности по открытию на ранних стадиях, таких как проверка цели и идентификация попадания.
Технология DECL предполагает конъюгацию химических соединений или строительных блоков с короткими фрагментами ДНК , которые служат идентификационными штрих-кодами, а в некоторых случаях также направляют и контролируют химический синтез. Этот метод позволяет массово создавать и опрашивать библиотеки посредством аффинной селекции, обычно на иммобилизованной белковой мишени. Недавно был разработан гомогенный метод скрининга библиотек, кодируемых ДНК (DEL), который использует технологию эмульсии вода в масле для выделения, подсчета и идентификации отдельных комплексов лиганд-мишень в однопробирочном подходе. В отличие от традиционных процедур скрининга, таких как высокопроизводительный скрининг , биохимические анализы не требуются для идентификации связующего вещества, что в принципе позволяет изолировать связующие вещества для широкого спектра белков, которые исторически было трудно решить с помощью традиционных технологий скрининга. Таким образом, в дополнение к общему открытию целевых специфических молекулярных соединений, доступность связующих фармакологически важных, но пока «не поддающихся лечению» целевых белков открывает новые возможности для разработки новых лекарств от заболеваний, которые до сих пор не поддавались лечению. При устранении требования первоначальной оценки активности совпадений можно надеяться и ожидать, что многие из идентифицированных связывающих веществ с высокой аффинностью будут показаны как активные в независимом анализе выбранных совпадений, что предложит эффективный метод выявления высококачественных совпадений и фармацевтических потенциальных клиентов. .
Химические библиотеки, закодированные ДНК, и технологии отображения
[ редактировать ]До недавнего времени применение молекулярной эволюции в лаборатории ограничивалось технологиями отображения биологических молекул, тогда как открытие малых молекул считалось выходящим за рамки этого биологического подхода. DEL открыли область технологий отображения, включив в нее неприродные соединения, такие как небольшие молекулы, расширяя применение молекулярной эволюции и естественного отбора до идентификации низкомолекулярных соединений с желаемой активностью и функцией.Химические библиотеки, закодированные в ДНК, имеют сходство с технологиями биологического дисплея, такими как технология фагового дисплея антител , дрожжевой дисплей , дисплей мРНК и аптамер SELEX . При фаговом дисплее антител антитела физически связаны с фаговыми частицами, несущими ген, кодирующий присоединенное антитело, что эквивалентно физическому соединению «фенотипа » (белка) и « генотипа » (гена, кодирующего белок). ). [1] Антитела, отображаемые на фагах, можно выделить из больших библиотек антител, имитируя молекулярную эволюцию : посредством раундов селекции (на иммобилизованном белке-мишени), амплификации и трансляции. [2] В DEL связь небольшой молекулы с кодом ДНК-идентификатора позволяет легко идентифицировать связывающие молекулы. DEL подвергаются процедурам селекции по аффинности на выбранном иммобилизованном целевом белке, после чего несвязывающие вещества удаляются этапами промывания, а связующие впоследствии могут быть амплифицированы с помощью полимеразной цепной реакции (ПЦР) и идентифицированы на основании их кода ДНК (например, по ДНК секвенирование). В основанных на эволюции технологиях DEL результаты могут быть дополнительно обогащены за счет проведения раундов селекции, ПЦР-амплификации и трансляции по аналогии с биологическими системами отображения, такими как фаговый дисплей на основе антител. Это дает возможность работать с гораздо большими библиотеками.
История
[ редактировать ]«Синтезировать многокомпонентную смесь соединений в одном процессе и фильтровать ее также в одном процессе». Это принцип комбинаторной химии, изобретенный профессором Фуркой А. (Университет Этвёша Лоранда в Будапеште, Венгрия) в 1982 году и описал его, включая метод синтеза комбинаторных библиотек и стратегию деконволюции, в документе, нотариально заверенном в том же году. [3] Мотивы, которые привели к изобретению, были опубликованы в 2002 году. [4] Химические библиотеки, кодируемые ДНК (DECL), синтезируются по принципу комбинаторной химии, что явно согласуется с их применением. [5]

Концепция ДНК-кодирования была впервые описана в теоретической статье Бреннера и Лернера в 1992 году, в которой было предложено связать каждую молекулу химически синтезированного объекта с определенной олигонуклеотидной последовательностью, сконструированной параллельно, и использовать эту кодирующую генетическую метку для идентификации и обогащают активными соединениями. [6] В 1993 году первая практическая реализация этого подхода была представлена Дж. Нильсеном, С. Бреннером и К. Янда, а также группой М. А. Галлопа. [7] [8] Бреннер и Янда предложили генерировать отдельные кодируемые члены библиотеки путем попеременного параллельного комбинаторного синтеза гетерополимерного химического соединения и соответствующей олигонуклеотидной последовательности на одной и той же бусине методом «разделения и объединения» (см. ниже). [7]
Поскольку незащищенная ДНК ограничена узким окном обычных условий реакции, до конца 1990-х годов рассматривался ряд альтернативных стратегий кодирования (т. е. на основе MS мечение соединений , кодирование пептидов , галоароматическое мечение, кодирование вторичными аминами , полупроводниковые устройства). ), главным образом, чтобы избежать неудобного твердофазного синтеза ДНК и создать легко проверяемые комбинаторные библиотеки с высокой производительностью. [9] Однако избирательная амплификация ДНК значительно облегчает скрининг библиотек и становится незаменимой для кодирования библиотек органических соединений такого беспрецедентного размера. Следовательно, в начале 2000-х годов ДНК-комбинаторная химия пережила возрождение.
В начале тысячелетия появилось несколько независимых разработок в области технологии DEL. Эти технологии можно разделить на две общие категории: неэволюционные и эволюционные технологии DEL, способные к молекулярной эволюции . Первая категория выигрывает от возможности использовать готовые реагенты и, следовательно, позволяет довольно просто создавать библиотеки. Совпадения можно идентифицировать с помощью секвенирования ДНК, однако трансляция ДНК и, следовательно, молекулярная эволюция с помощью этих методов невозможны. Под эту категорию подпадают подходы разделения и пула, разработанные исследователями Praecis Pharmaceuticals (ныне принадлежащей GlaxoSmithKline), Nuevolution (Копенгаген, Дания) и технология ESAC, разработанная в лаборатории профессора Д. Нери (Институт фармацевтических наук, Цюрих, Швейцария). . Технология ESAC выделяется как комбинаторный подход самосборки, который напоминает обнаружение совпадений на основе фрагментов (рис. 1b). Здесь отжиг ДНК позволяет отбирать образцы дискретных комбинаций строительных блоков, но между ними не происходит химической реакции. Примерами технологий DEL, основанных на эволюции, являются маршрутизация ДНК, разработанная профессором Д.Р. Хэлпином и профессором П.Б. Харбери (Стэнфордский университет, Стэнфорд, Калифорния), синтез с помощью матрицы ДНК, разработанный проф. Д. Лю (Гарвардский университет, Кембридж, Массачусетс) и коммерциализирован компанией Ensemble Therapeutics (Кембридж, Массачусетс) и технологией YoctoReactor. [10] разработан и коммерциализирован компанией Vipergen (Копенгаген, Дания). Эти технологии более подробно описаны ниже. Синтез по шаблону ДНК и технология YoctoReactor требуют предварительного конъюгирования химических строительных блоков (BB) с меткой ДНК-олигонуклеотида перед сборкой библиотеки, поэтому перед сборкой библиотеки требуется дополнительная предварительная работа. Кроме того, BB, помеченные ДНК, позволяют генерировать генетический код для синтезированных соединений, и возможна искусственная трансляция генетического кода: то есть BB могут быть вызваны с помощью генетического кода, амплифицированного ПЦР, и соединения библиотеки могут быть регенерированы. Это, в свою очередь, позволяет применить дарвиновский принцип естественного отбора и эволюции к отбору малых молекул по прямой аналогии с биологическими системами отображения; через раунды отбора, усиления и перевода.
Неэволюционные технологии
[ редактировать ]Этот раздел пуст. Вы можете помочь, добавив к нему . ( сентябрь 2022 г. ) |
Комбинаторные библиотеки
[ редактировать ]Комбинаторные библиотеки представляют собой специальные многокомпонентные смеси соединений, которые синтезируются в один этап. Они отличаются как от совокупности отдельных соединений, так и от ряда соединений, полученных параллельным синтезом.Комбинаторные библиотеки имеют важные особенности.
″ При их синтезе используются смеси. Использование смесей обеспечивает очень высокую эффективность процесса. Оба реагента могут представлять собой смеси, но по практическим соображениям используется процедура разделения смеси: одна смесь делится на части, которые связаны с ББ. [11] [12] Смеси настолько важны, что не существует комбинаторной библиотеки без использования смеси в синтезе, и если смесь используется в процессе, неизбежно образуется комбинаторная библиотека.
″ Компоненты библиотеки должны присутствовать примерно в равных молярных количествах. Чтобы добиться этого как можно точнее, смеси делят на равные порции и после объединения необходимо тщательное перемешивание.
″ Поскольку структура компонентов неизвестна, при скрининге необходимо использовать методы деконволюции. По этой причине были разработаны методы кодирования. Кодирующие молекулы прикреплены к шарикам твердого носителя, которые записывают связанные ББ и их последовательность. Одним из таких методов является кодирование ДНК-олигомерами.
«Примечательной особенностью комбинаторных библиотек является то, что всю смесь соединений можно проверить за один процесс.
Поскольку и синтез, и скрининг являются очень эффективными процедурами, использование комбинаторных библиотек в фармацевтических исследованиях приводит к огромной экономии.
При твердофазном комбинаторном синтезе в каждой бусине образуется только одно соединение. По этой причине количество компонентов в библиотеке не может превышать количество шариков твердой опоры. Это означает, что количество компонентов в таких библиотеках ограничено. Это ограничение было устранено Харбери и Хэлпином. В их синтезе DEL твердая подложка отсутствует, и BB прикрепляются непосредственно к кодирующим олигомерам ДНК. [13] Этот новый подход позволяет практически неограниченно увеличивать количество компонентов комбинаторных библиотек, кодируемых ДНК (DECL).
Кодирование ДНК с разделением и объединением
[ редактировать ]Чтобы применить комбинаторную химию для синтеза химических библиотек, кодируемых ДНК, был использован подход Split-&Pool. [11] [12] Первоначально набор уникальных ДНК- олигонуклеотидов ( n ), каждый из которых содержит определенную кодирующую последовательность, химически конъюгируется с соответствующим набором небольших органических молекул. Следовательно, соединения-конъюгаты олигонуклеотида смешиваются («Объединить») и разделяются («Расщепляются») на ряд групп ( m ). В соответствующих условиях второй набор строительных блоков (m) соединяют с первым и перед повторным смешиванием ферментативно вводят дополнительный олигонуклеотид , который кодирует вторую модификацию. Эти шаги «разделения и объединения» можно повторять несколько раз ( r ), увеличивая на каждом этапе размер библиотеки комбинаторным образом (т. е. ( n x m ) р ). Альтернативно, пептидные нуклеиновые кислоты использовались для кодирования библиотек, полученных методом «разделения и объединения». [14] Преимущество PNA-кодирования заключается в том, что химию можно выполнять с помощью стандартного SPPS. [15]
Поэтапное соединение кодирующих фрагментов ДНК с возникающими органическими молекулами.
[ редактировать ]
Многообещающая стратегия создания библиотек, кодируемых ДНК, представляет собой использование многофункциональных строительных блоков, ковалентно конъюгированных с олигонуклеотидом, служащим «основной структурой» для синтеза библиотеки. По принципу «объединить и разделить» набор многофункциональных каркасов подвергается ортогональным реакциям с рядом подходящих реактивных партнеров. После каждого этапа реакции идентичность модификации кодируется ферментативным добавлением сегмента ДНК к исходной «ядерной структуре» ДНК. [16] [17] Использование N -защищенных аминокислот, ковалентно присоединенных к фрагменту ДНК, позволяет после подходящей стадии снятия защиты обеспечить дальнейшее амидной связи с рядом карбоновых кислот или восстановительное аминирование альдегидами образование . Аналогичным образом, диенкарбоновые кислоты, используемые в качестве каркасов для построения библиотеки на 5'-конце аминомодифицированного олигонуклеотида , могут быть подвергнуты реакции Дильса-Альдера с различными производными малеимида . После завершения желаемой стадии реакции идентичность химического фрагмента, добавленного к олигонуклеотиду, устанавливают путем отжига частично комплементарного олигонуклеотида и последующей Кленова с заполнением полимеризации ДНК , с получением двухцепочечного фрагмента ДНК. Описанные выше стратегии синтеза и кодирования позволяют легко создавать библиотеки, кодируемые ДНК, размером до 10 4 соединения-члены, несущие два набора «строительных блоков». Однако поэтапное добавление по крайней мере трех независимых наборов химических фрагментов к трехфункциональному строительному блоку для построения и кодирования очень большой библиотеки, кодируемой ДНК (содержащей до 10 6 соединения) также можно предусмотреть. [16] ( Рис.2 )
Комбинаторная самосборка
[ редактировать ]Закодированные самособирающиеся химические библиотеки
[ редактировать ]
Закодированные библиотеки самосборочные химические 10 основаны на принципе , ESAC ( ) согласно которому две подбиблиотеки размером x элементов (например, 3 ), содержащий постоянный комплементарный домен гибридизации, может после гибридизации дать комбинаторную библиотеку ДНК-дуплексов со сложностью x 2 равномерно представленные члены библиотеки (например, 10 6 ). [18] Каждый член подбиблиотеки будет состоять из олигонуклеотида , содержащего вариабельную кодирующую область, фланкированную постоянной последовательностью ДНК, несущей подходящую химическую модификацию на конце олигонуклеотида. [18] Подбиблиотеки ESAC можно использовать по меньшей мере в четырех различных вариантах реализации. [18]
- Подбиблиотеку можно соединить с комплементарным олигонуклеотидом и использовать в качестве библиотеки, кодируемой ДНК, отображающей одно ковалентно связанное соединение, для экспериментов по селекции на основе аффинности.
- Подбиблиотеку можно объединить с олигонуклеотидом, имеющим известное связывание с мишенью, что позволяет реализовать стратегии созревания аффинности.
- Две отдельные подбиблиотеки можно собрать комбинаторно и использовать для de novo идентификации бидентатно- связывающих молекул.
- Три разные подбиблиотеки могут быть собраны в комбинаторную триплексную библиотеку.
Предпочтительные связывающие вещества, выделенные в результате отбора на основе аффинности, могут быть амплифицированы с помощью ПЦР и декодированы на комплементарных олигонуклеотидов. микрочипах [19] или путем конкатенации кодов, субклонирования и секвенирования . [18] Отдельные строительные блоки в конечном итоге могут быть конъюгированы с использованием подходящих линкеров с получением соединения с высоким сродством, подобного лекарственному средству. Характеристики линкера (например, длина, гибкость, геометрия, химическая природа и растворимость) влияют на аффинность связывания и химические свойства полученного связующего. ( Рис.3 ).
Эксперименты по биопэннингу HSA из 600 членов библиотеки ESAC позволили выделить 4-( п -йодфенил)бутановый фрагмент. Это соединение представляет собой основную структуру серии переносимых молекул, связывающих альбумин , и альбуфлюора, недавно разработанного флуоресцентного ангиографического контрастного агента, который в настоящее время проходит клиническую оценку. [20]
Технология ESAC использовалась для выделения мощных ингибиторов бычьего трипсина и для идентификации новых ингибиторов стромелизина -1 ( MMP-3 ), матриксной металлопротеиназы, участвующей как в физиологических, так и в патологических процессах ремоделирования тканей, а также в болезненных процессах. , такие как артрит и метастазы . [21]
Технологии, основанные на эволюции
[ редактировать ]ДНК-маршрутизация
[ редактировать ]В 2004 году Д. Р. Халпин и П. Б. Харбери представили новый интригующий метод создания библиотек, кодируемых ДНК. Впервые конъюгированные с ДНК матрицы послужили как для кодирования, так и для программирования инфраструктуры синтеза компонентов библиотеки по принципу «разделение и объединение». [22] Конструкция Халпина и Харбери позволила чередовать раунды селекции, ПЦР-амплификации и диверсификации с помощью небольших органических молекул, что полностью аналогично технологии фагового дисплея . Механизм маршрутизации ДНК состоит из серии соединенных колонок, несущих связанные со смолой антикодоны, которые могут специфично разделять популяцию ДНК-матриц в пространственно различные местоположения путем гибридизации . [22] В соответствии с этим протоколом разделения и объединения пептидная комбинаторная библиотека, кодирующая ДНК 10 6 члены были созданы. [23]
Синтез по шаблону ДНК
[ редактировать ]
В 2001 году Дэвид Лю и его коллеги показали, что комплементарные ДНК- олигонуклеотиды могут использоваться для содействия определенным синтетическим реакциям , которые неэффективно протекают в растворе при низкой концентрации . [24] [25] ДНК-гетеродуплекс использовался для ускорения реакции между химическими фрагментами, расположенными на концах двух цепей ДНК. Более того, было показано, что «эффект близости», ускоряющий бимолекулярную реакцию, не зависит от расстояния (по крайней мере, на расстоянии 30 нуклеотидов ). [24] [25] с помощью запрограммированной последовательности Олигонуклеотиды, несущие одну химическую реагентную группу, гибридизовали с комплементарными производными олигонуклеотидов, несущими другую реакционноспособную химическую группу. Близость, обеспечиваемая гибридизацией ДНК, резко увеличивает эффективную молярность реакционных реагентов, присоединенных к олигонуклеотидам, позволяя желаемой реакции происходить даже в водной среде при концентрациях, которые на несколько порядков ниже, чем те, которые необходимы для соответствующей традиционной органической реакции. не шаблонизированный ДНК. [26] Используя шаблон ДНК и программируемый последовательность синтеза, Лю и его коллеги создали 64-членную библиотеку макроциклов, кодируемую ДНК . [27]
Трехмерная бесконтактная технология (технология YoctoReactor)
[ редактировать ]YoctoReactor (yR) — это трехмерный подход, основанный на близости, который использует природу самосборки олигонуклеотидов ДНК в 3, 4 или 5-сторонние соединения для направления синтеза малых молекул в центре соединения. Рисунок 5 иллюстрирует основную концепцию четырехстороннего соединения ДНК.
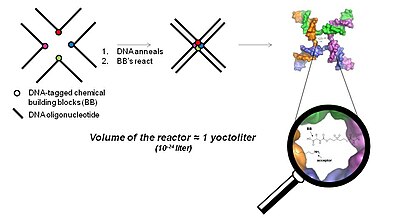
Центр соединения ДНК составляет объем порядка йоктолитра , отсюда и название YoctoReactor. Этот объем содержит реакцию одной молекулы, дающую реакционные концентрации в диапазоне высоких мМ. Эффективная концентрация, обеспечиваемая ДНК, значительно ускоряет химические реакции, которые в противном случае не происходили бы при фактической концентрации на несколько порядков ниже.
Создание библиотеки YR
[ редактировать ]На рисунке 6 показано создание библиотеки yR с использованием трехстороннего соединения ДНК.
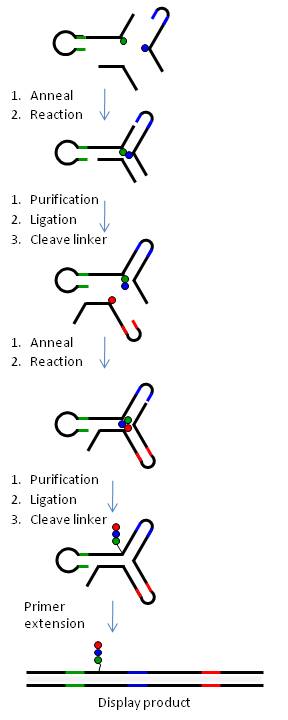
Таким образом, химические строительные блоки (BB) присоединяются через расщепляемые или нерасщепляемые линкеры к трем типам биспецифических ДНК-олигонуклеотидов (олиго-BB), представляющих каждое плечо yR. Чтобы облегчить синтез комбинаторным способом, олиго-BB сконструированы таким образом, что ДНК содержит (а) код прикрепленного BB на дистальном конце олиго (цветные линии) и (b) области постоянной последовательности ДНК (черные линии). линии), чтобы вызвать самосборку ДНК в трехстороннее соединение (независимо от BB) и последующую химическую реакцию. Химические реакции проводятся поэтапно, и после каждого этапа ДНК лигируется, а продукт очищается электрофорезом в полиакриламидном геле. Расщепляемые линкеры (BB-ДНК) используются во всех положениях, кроме одной, образуя библиотеку небольших молекул с единственной ковалентной связью с кодом ДНК. В таблице 1 показано, как с помощью технологии yR можно создавать библиотеки разных размеров.

Подход к проектированию yR обеспечивает неизменное место реакции как в отношении (а) расстояния между реагентами, так и (б) среды последовательности, окружающей место реакции. Более того, тесная связь между кодом и BB на фрагментах олиго-BB, которые комбинаторно смешиваются в одном горшке, обеспечивает высокую точность кодирования библиотеки. Более того, код синтезированных продуктов не предустановлен, а скорее собирается комбинаторно и синтезируется синхронно с врожденным продуктом.
Гомогенный скрининг йореакторных библиотек
[ редактировать ]Недавно был разработан гомогенный метод скрининга йореакторных библиотек (yR), в котором используется технология эмульсии вода в масле для выделения отдельных комплексов лиганд-мишень. Называемое обогащением связующей ловушки (BTE), лиганды белка-мишени идентифицируются путем захвата связывающих пар (белок-мишень, меченный ДНК, и лиганд yR) в каплях эмульсии во время кинетики, в которой доминирует диссоциация. После захвата ДНК-мишень и лиганд соединяются путем лигирования, сохраняя таким образом информацию о связывании.
В дальнейшем идентификация совпадений представляет собой, по сути, подсчет: информация о событиях связывания расшифровывается путем секвенирования и подсчета присоединенной ДНК - селективные связыватели подсчитываются с гораздо большей частотой, чем случайные связыватели. Это возможно, поскольку случайный захват мишени и лиганда «разбавляется» большим количеством капель воды в эмульсии. Низкий уровень шума и фонового сигнала, характерный для заушного слухового аппарата, объясняется «разбавлением» случайного сигнала, отсутствием поверхностных артефактов и высокой точностью библиотеки yR и метода скрининга. Скрининг проводится методом одной пробирки. Биологически активные совпадения выявляются при одном раунде заушной терапии и характеризуются низким уровнем ложноположительных результатов.
BTE имитирует неравновесную природу взаимодействий лиганд-мишень in vivo и предлагает уникальную возможность скрининга специфических лигандов-мишеней на основе времени пребывания лиганд-мишень, поскольку эмульсия, которая улавливает связывающий комплекс, образуется во время фазы динамической диссоциации.
Расшифровка химических библиотек, закодированных ДНК
[ редактировать ]После отбора из химических библиотек, закодированных ДНК, стратегия декодирования для быстрой и эффективной идентификации специфически связывающихся соединений имеет решающее значение для дальнейшего развития технологии DEL . До сих пор декодирование на основе секвенирования по Сэнгеру , методология на основе микрочипов и методы высокопроизводительного секвенирования представляли собой основные методологии декодирования выборок библиотек, закодированных ДНК.
Декодирование на основе секвенирования по Сэнгеру
[ редактировать ]Хотя многие авторы неявно предполагали традиционное декодирование на основе секвенирования Сэнгера , [7] [8] [18] [23] [27] количество кодов, которые нужно упорядочить просто в зависимости от сложности библиотеки, определенно является нереальной задачей для традиционного подхода к секвенированию Сэнгера . Тем не менее, впервые было описано применение секвенирования по Сэнгеру для высокопроизводительного декодирования химических библиотек, закодированных ДНК. [18] После отбора и ПЦР-амплификации ДНК-меток соединений библиотеки были созданы конкатамеры, содержащие множественные кодирующие последовательности, и лигированы в вектор . После секвенирования по Сэнгеру репрезентативного числа полученных колоний были выявлены частоты кодов, присутствующих в образце библиотеки, кодируемой ДНК, до и после отбора. [18]
Декодирование на основе микрочипов
[ редактировать ]ДНК- микрочип — устройство для высокопроизводительных исследований, широко используемое в молекулярной биологии и медицине . Он состоит из серии микроскопических пятен («особенностей» или «локаций»), содержащих несколько пикомолей олигонуклеотидов, несущих определенную последовательность ДНК. Это может быть короткий участок гена или другой элемент ДНК, который используется в качестве зондов для гибридизации образца ДНК или РНК в подходящих условиях. зонда-мишени Гибридизацию обычно выявляют и количественно определяют с помощью флуоресцентного обнаружения меченных флуорофором мишеней для определения относительного содержания последовательностей нуклеиновых кислот- мишеней . Микрочип использовался для успешного декодирования библиотек, закодированных ДНК ESAC. [18] и библиотеки, закодированные в PNA. [28] Кодирующие олигонуклеотиды, представляющие отдельные химические соединения в библиотеке, наносятся и химически связываются на предметных стеклах микроматрицы с помощью робота BioChip Arrayer. Впоследствии олигонуклеотидные метки связывающих соединений, выделенные из селекции, амплифицируются с помощью ПЦР с использованием флуоресцентного праймера и гибридизуются на предметном стекле ДНК- микрочипа . После этого микроматрицы анализируются с помощью лазерного сканирования, а интенсивность пятен обнаруживается и количественно оценивается. Обогащение предпочтительными связывающими соединениями выявляется при сравнении интенсивности пятен на предметном стекле ДНК- микрочипа до и после отбора. [18]
Декодирование с помощью высокопроизводительного секвенирования
[ редактировать ]В зависимости от сложности химической библиотеки, кодируемой ДНК (обычно от 10 3 и 10 6 участники), традиционное декодирование на основе секвенирования по Сэнгеру вряд ли будет применимо на практике как из-за высокой стоимости основания для секвенирования, так и из-за утомительной процедуры. [29] В технологиях высокопроизводительного секвенирования используются стратегии, которые распараллеливают процесс секвенирования, заменяя использование капиллярного электрофореза и создавая тысячи или миллионы последовательностей одновременно. В 2008 году было описано первое внедрение метода высокопроизводительного секвенирования , первоначально разработанного для секвенирования генома (т.е. « технология 454 ») для быстрого и эффективного декодирования химической библиотеки, закодированной ДНК, содержащей 4000 соединений. [16] Это исследование привело к идентификации новых химических соединений с субмикромолярными константами диссоциации по отношению к стрептавидину и определенно показало возможность создания, выполнения селекции и декодирования библиотек, закодированных ДНК, содержащих миллионы химических соединений. [16]
См. также
[ редактировать ]- Открытие лекарств
- Высокопроизводительный скрининг
- Комбинаторная химия
- секвенирование ДНК
- Фаговый дисплей
Ссылки
[ редактировать ]- ^ Смит ГП (июнь 1985 г.). «Нитевидный слитый фаг: новые векторы экспрессии, которые отображают клонированные антигены на поверхности вириона». Наука . 228 (4705): 1315–7. Бибкод : 1985Sci...228.1315S . дои : 10.1126/science.4001944 . ПМИД 4001944 .
- ^ Хугенбум HR (2002). «Обзор технологии фагового дисплея антител и ее применения». Фаговый дисплей антител . Методы молекулярной биологии. Том. 178. Хумана Пресс. стр. 1–37. дои : 10.1385/1-59259-240-6:001 . ISBN 978-1-59259-240-1 . ПМИД 11968478 .
- ^ Фурка А. Исследование возможности систематического поиска пептидов, пригодных для медицинского применения. Исследование возможности систематического поиска фармацевтически полезных пептидов) https://mersz.hu/mod/object.php?objazonosito=matud202006_f42772_i2
- ^ Фурка А (2002). Комбинаторная химия 20 лет спустя.., Drug DiscovToday 7; 1-4.
- ^ Гуднау, Роберт А. младший (2014). РУКОВОДСТВО ПО ХИМИИ, ЗАКОДИРОВАННОЙ FORDNA. Теория и приложения для исследования химического космоса и открытия лекарств . Уолтем, Массачусетс, США: John Wiley & Sons, Inc., Хобокен, Нью-Джерси. стр. 2–13.
- ^ Бреннер С., Лернер Р.А. (июнь 1992 г.). «Закодированная комбинаторная химия» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (12): 5381–3. Бибкод : 1992PNAS...89.5381B . дои : 10.1073/pnas.89.12.5381 . ПМК 49295 . ПМИД 1608946 .
- ^ Перейти обратно: а б с Нильсен Дж., Бреннер С., Янда К.Д. (1993). «Синтетические методы реализации закодированной комбинаторной химии». Журнал Американского химического общества . 115 (21): 9812–9813. дои : 10.1021/ja00074a063 .
- ^ Перейти обратно: а б Ниделс М.К., Джонс Д.Г., Тейт Э.Х., Хейнкель Г.Л., Кочерспергер Л.М., Дауэр В.Дж., Барретт Р.В., Галлоп М.А. (ноябрь 1993 г.). «Создание и скрининг библиотеки синтетических пептидов, кодируемых олигонуклеотидами» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (22): 10700–4. Бибкод : 1993PNAS...9010700N . дои : 10.1073/pnas.90.22.10700 . ПМЦ 47845 . ПМИД 7504279 .
- ^ Мукунд С. Чоргаде (2006). Открытие и разработка лекарств . Нью-Йорк: Wiley-Interscience. стр. 129–167. ISBN 978-0-471-39848-6 .
- ^ Хейтнер Т.Р., Хансен, штат Нью-Джерси (ноябрь 2009 г.). «Упрощение обнаружения и оптимизации хитов с помощью ДНК-реактора масштаба йоктолитров». Мнение экспертов об открытии лекарств . 4 (11): 1201–13. дои : 10.1517/17460440903206940 . ПМИД 23480437 . S2CID 20881306 .
- ^ Перейти обратно: а б А. Фурка, Ф. Себастьен, М. Асгедом, Г. Дибо, «Рог изобилия пептидов путем синтеза в основных моментах современной биохимии», материалы 14-го Международного биохимического конгресса, ВСП. Утрехт, Нидерланды, 1988 г., Том. 5, стр. 47.
- ^ Перейти обратно: а б Фурка А., Себастьен Ф., Асгедом М., Дибо Г. (1991) Общий метод быстрого синтеза многокомпонентных пептидных смесей. Int J пептидный белок Res 37; 487-93.
- ^ Харбери Д.Р., Халпин Д.Р. (2000) WO 00/23458.
- ^ Винсингер Н., Дамуазо Р., Талли Д.С., Гейерстангер Б.Х., Бердик К., Харрис Дж.Л. (октябрь 2004 г.). «Микрочипы с протеазным субстратом, кодируемым PNA» . Химия и биология . 11 (10): 1351–60. doi : 10.1016/j.chembiol.2004.07.015 . ПМИД 15489162 .
- ^ Замбальдо К., Барлуэнга С., Винсингер Н. (июнь 2015 г.). «Химические библиотеки, закодированные в PNA». Современное мнение в области химической биологии . 26 :8–15. дои : 10.1016/j.cbpa.2015.01.005 . ПМИД 25621730 .
- ^ Перейти обратно: а б с д Манноччи Л., Чжан Й., Шойерманн Дж., Леймбахер М., Де Беллис Г., Рицци Е., Думелин С., Мелькко С., Нери Д. (ноябрь 2008 г.). «Высокопроизводительное секвенирование позволяет идентифицировать связывающие молекулы, выделенные из химических библиотек, закодированных ДНК» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (46): 17670–5. Бибкод : 2008PNAS..10517670M . дои : 10.1073/pnas.0805130105 . ПМЦ 2584757 . ПМИД 19001273 .
- ^ Буллер Ф., Манноччи Л., Чжан Й., Дюмелин С.Э., Шойерманн Дж., Нери Д. (ноябрь 2008 г.). «Разработка и синтез новой химической библиотеки, кодируемой ДНК, с использованием циклоприсоединений Дильса-Альдера». Письма по биоорганической и медицинской химии . 18 (22): 5926–31. дои : 10.1016/j.bmcl.2008.07.038 . ПМИД 18674904 .
- ^ Перейти обратно: а б с д и ж г час я Мелькко С., Шойерманн Дж., Дюмелин С.Э., Нери Д. (май 2004 г.). «Закодированные самособирающиеся химические библиотеки». Природная биотехнология . 22 (5): 568–74. дои : 10.1038/nbt961 . ПМИД 15097996 . S2CID 24806778 .
- ^ Ловринович М., Нимейер К.М. (май 2005 г.). «Микрочипы ДНК как инструменты декодирования в комбинаторной химии и химической биологии». Ангеванде Хеми . 44 (21): 3179–83. дои : 10.1002/anie.200500645 . ПМИД 15861437 .
- ^ Дюмелен С.Э., Трюссель С., Буллер Ф., Траксель Е., Бутц Ф., Чжан Й., Манноччи Л., Бек С.К., Друмеа-Миранча М., Зеелигер М.В., Балтес К., Мюгглер Т., Кранц Ф., Рудин М., Мелькко С., Шойерманн Дж., Нери Д (2008). «Портативное связующее вещество альбумина из химической библиотеки, закодированной ДНК». Прикладная химия . 47 (17): 3196–201. дои : 10.1002/anie.200704936 . ПМИД 18366035 .
- ^ Мелькко С., Чжан Й., Дюмелин С.Э., Шойерманн Дж., Нери Д. (2007). «Выделение высокоаффинных ингибиторов трипсина из химической библиотеки, кодируемой ДНК». Ангеванде Хеми . 46 (25): 4671–4. дои : 10.1002/anie.200700654 . ПМИД 17497616 .
- ^ Перейти обратно: а б Халпин Д.Р., Харбери П.Б. (июль 2004 г.). «ДНК-дисплей I. Маршрутизация популяций ДНК, закодированная последовательностями» . ПЛОС Биология . 2 (7): Е173. дои : 10.1371/journal.pbio.0020173 . ПМК 434148 . ПМИД 15221027 .

- ^ Перейти обратно: а б Халпин Д.Р., Харбери П.Б. (июль 2004 г.). «Дисплей ДНК II. Генетические манипуляции с библиотеками комбинаторной химии для эволюции малых молекул» . ПЛОС Биология . 2 (7): Е174. дои : 10.1371/journal.pbio.0020174 . ПМЦ 434149 . ПМИД 15221028 .
- ^ Перейти обратно: а б Gartner ZJ, Лю DR (июль 2001 г.). «Общность синтеза по шаблону ДНК как основа для эволюции неприродных малых молекул» . Журнал Американского химического общества . 123 (28): 6961–3. дои : 10.1021/ja015873n . ПМЦ 2820563 . ПМИД 11448217 .
- ^ Перейти обратно: а б Кальдероне К.Т., Пакетт Дж.В., Гартнер З.Дж., Лю Д.Р. (ноябрь 2002 г.). «Направление несовместимых в противном случае реакций в одном растворе с использованием органического синтеза по шаблону ДНК». Ангеванде Хеми . 41 (21): 4104–8. doi : 10.1002/1521-3773(20021104)41:21<4104::AID-ANIE4104>3.0.CO;2-O . ПМИД 12412096 .
- ^ Ли Х, Лю Д.Р. (сентябрь 2004 г.). «Органический синтез по шаблону ДНК: стратегия природы по контролю химической реактивности применительно к синтетическим молекулам». Ангеванде Хеми . 43 (37): 4848–70. дои : 10.1002/anie.200400656 . ПМИД 15372570 .
- ^ Перейти обратно: а б Гартнер З.Дж., Це Б.Н., Грубина Р., Дойон Дж.Б., Снайдер Т.М., Лю Д.Р. (сентябрь 2004 г.). «Органический синтез по шаблону ДНК и подбор библиотеки макроциклов» . Наука . 305 (5690): 1601–5. Бибкод : 2004Sci...305.1601G . дои : 10.1126/science.1102629 . ПМК 2814051 . ПМИД 15319493 .
- ^ Харрис Дж., Мейсон Д.Э., Ли Дж., Бердик К.В., Бэкес Б.Дж., Чен Т., Шипуэй А, Ван Хик Г., Гоф Л., Гаеммагами А., Шакиб Ф., Дебен Ф., Винсингер Н. (октябрь 2004 г.). «Профиль активности экстракта аллергенов пылевых клещей с использованием библиотек субстратов и функциональных протеомных микроматриц» . Химия и биология . 11 (10): 1361–72. doi : 10.1016/j.chembiol.2004.08.008 . ПМИД 15489163 .
- ^ Сэнгер Ф., Никлен С., Коулсон А.Р. (декабрь 1977 г.). «Секвенирование ДНК с ингибиторами обрыва цепи» . Труды Национальной академии наук Соединенных Штатов Америки . 74 (12): 5463–7. Бибкод : 1977PNAS...74.5463S . дои : 10.1073/pnas.74.12.5463 . ПМК 431765 . ПМИД 271968 .