Мембранный липид
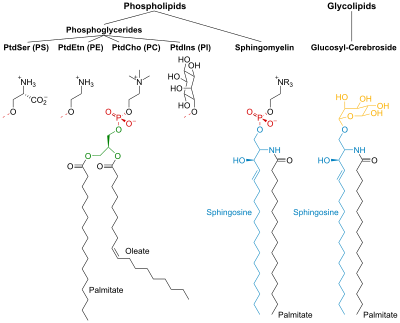
PtdCho – фосфатидилхолин ; PtdEtn – фосфатидилэтаноламин ; PtdIns - Фосфатидилинозитол ; PtdSer – фосфатидилсерин .
Мембранные липиды представляют собой группу соединений (структурно подобных жирам и маслам), которые образуют липидный бислой клеточной мембраны . Тремя основными классами мембранных липидов являются фосфолипиды , гликолипиды и холестерин . Липиды амфифильны : у них есть один конец, растворимый в воде («полярный»), и конец, растворимый в жире («неполярный»). Образуя двойной слой с полярными концами, направленными наружу, и неполярными концами, направленными внутрь, мембранные липиды могут образовывать «липидный бислой», который удерживает водную внутреннюю часть клетки отдельно от водянистой внешней части. Расположение липидов и различных белков, действующих как рецепторы и поры каналов в мембране, контролирует вход и выход других молекул и ионов в рамках клеточного метаболизма. Для выполнения физиологических функций мембранным белкам позволяется вращаться и диффундировать латерально в двумерном пространстве липидного бислоя благодаря наличию липидной оболочки, тесно прикрепленной к поверхности белка, называемой кольцевая липидная оболочка .
Биологические роли
[ редактировать ]Бислой, образованный мембранными липидами, служит удерживающей единицей живой клетки. Мембранные липиды также образуют матрицу, в которой находятся мембранные белки . Исторически считалось, что липиды выполняют лишь структурную роль. Функциональные роли липидов на самом деле многочисленны: они служат регуляторными агентами клеток роста и адгезии . Они участвуют в биосинтезе других биомолекул . Они могут служить для повышения ферментативной активности ферментов . [ 1 ]
Липид , не образующий бислой, такой как моногалактозилдиглицерид (МГДГ), преобладает среди липидов в тилакоидных мембранах, которые при гидратации в отдельности образуют обращенную гексагональную цилиндрическую фазу. Однако в сочетании с другими липидами и каротиноидами / хлорофиллами тилакоидных мембран они тоже образуют липидные бислои. [ 2 ]
Основные классы
[ редактировать ]фосфолипиды
[ редактировать ]Фосфолипиды и гликолипиды состоят из двух длинных неполярных ( гидрофобных ) углеводородных цепей, связанных с гидрофильной головной группой.
Головки фосфолипидов фосфорилированы и состоят из:
- Глицерин (отсюда и название фосфоглицериды, данное этой группе липидов), или
- Сфингозин (например, сфингомиелин и церамид ).
Тетраэфир диалкилглицерина (GDGT) помогает изучать древние факторы окружающей среды . [ 3 ]
Гликолипиды
[ редактировать ]Головки гликолипидов (глико-сахар) содержат сфингозин с присоединенными к нему одной или несколькими сахарными единицами . Гидрофобные цепи принадлежат либо:
- две жирные кислоты (ЖК) – в случае фосфоглицеридов, или
- одна ЖК и углеводородный хвост сфингозина – в случае сфингомиелина и гликолипидов.
Галактолипиды – моногалактозилдиглицерид (МГДГ) и дигалактозилдиглицерид (ДГДГ) образуют преобладающие липиды в мембранах тилакоидов хлоропластов высших растений; липосомальные структуры, образованные общим липидным экстрактом тилакоидных мембран, оказались чувствительными к сахарозе, поскольку она превращает бислои в мицеллярные структуры. [ 4 ]
Жирные кислоты
[ редактировать ]Жирные кислоты в фосфо- и гликолипидах обычно содержат четное число атомов углерода , обычно от 14 до 24 , причем наиболее распространенными являются 16- и 18-углеродные атомы. ЖК могут быть насыщенными и ненасыщенными, причем конфигурация двойных связей почти всегда цис . Длина и степень ненасыщенности цепей ЖК оказывают глубокое влияние на текучесть мембран . растений Мембраны тилакоидов сохраняют высокую текучесть даже при относительно низких температурах окружающей среды благодаря обилию 18-углеродных жирных ацильных цепей с тремя двойными связями - линоленовой кислоты , как было выявлено методами ЯМР 13-С. [ 5 ]
Фосфоглицериды
[ редактировать ]В фосфоглицеридах гидроксильные группы при С-1 и С-2 глицерина этерифицированы карбоксильными группами . жирных кислот Гидроксильная группа С-3 этерифицирована до фосфорной кислоты. Полученное соединение, называемое фосфатидатом , представляет собой простейший фосфоглицерат . В мембранах присутствуют лишь небольшие количества фосфатидата. Однако он является ключевым промежуточным продуктом в биосинтезе других фосфоглицеридов.
Сфинголипиды
[ редактировать ]Сфингозин — аминоспирт , содержащий длинную ненасыщенную углеводородную цепь. В сфингомиелине и гликолипидах аминогруппа сфингозина связана с ЖК амидной связью. В сфингомиелине первичная гидроксильная группа сфингозина этерифицирована до фосфорилхолина .

В гликолипидах к этой группе присоединяется сахарный компонент. Простейший гликолипид — цереброзид , в котором имеется только один сахарный остаток — Glc или Gal . Более сложные гликолипиды, такие как ганглиозиды , содержат разветвленную цепь, состоящую до семи остатков сахаров.
Стеролы
[ редактировать ]Самый известный стерол — холестерин , который содержится в организме человека. Холестерин также содержится в мембранах других клеток эукариот . Стеролы имеют гидрофобную четырехчленную жесткую структуру слитого кольца и небольшую полярную головную группу.
Холестерин биосинтезируется из мевалоната посредством скваленовой циклизации терпеноидов . Клеточные мембраны требуют высокого уровня холестерина – обычно в среднем 20% холестерина во всей мембране, локально увеличиваясь в участках плота до 50% холестерина (-% — молекулярное соотношение). [ 6 ] Он преимущественно связывается со сфинголипидами (см. Диаграмму) в богатых холестерином липидных плотах участков мембран эукариотических клеток. [ 7 ] Формирование липидных рафтов способствует агрегации периферических и трансмембранных белков, включая стыковку белков SNARE и VAMP . [ 8 ] Фитостерины , такие как ситостерин и стигмастерин, а также гопаноиды выполняют аналогичную функцию у растений и прокариот .
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ РБ Геннис. Биомембраны - молекулярная структура и функции. Спрингер-Верлаг, Нью-Йорк (1989).
- ^ YashRoy RC (1990)Ламеллярная дисперсия и фазовое разделение липидов мембран хлоропластов с помощью электронной микроскопии с отрицательным окрашиванием. Журнал биологических наук , том. 15(2), стр. 93-98. https://www.researchgate.net/publication/230820037_Lamellar_dispersion_and_phase_separation_of_chromoplast_membrane_lipids_by_negative_staining_electron_microscope?ev=prf_pub
- ^ Дальше; и др. (2007). «Экологический контроль распределения липидов бактериальных тетраэфирных мембран в почвах». Geochimica et Cosmochimica Acta . 71 (3): 703–713. Бибкод : 2007GeCoA..71..703W . дои : 10.1016/j.gca.2006.10.003 .
- ^ YashRoy RC (1994)Дестабилизация ламеллярной дисперсии липидов тилакоидных мембран сахарозой. Biochimica et Biophysical Acta , vol. 1212, стр. 129-133. https://www.researchgate.net/publication/15042978_Destabilisation_of_lamellar_dispersion_of_thylakoid_membrane_lipids_by_sucrose?ev=prf_pub
- ^ YashRoy RC (1987) 13-C ЯМР-исследования липидных жирно-ацильных цепей мембран хлоропластов. Индийский журнал биохимии и биофизики , том. 24(6), стр. 177-178. https://www.researchgate.net/publication/230822408_13-C_NMR_studies_of_lipid_fatty_acyl_chains_of_хлоропласты_мембраны?ev=prf_pub
- ^ де Мейер Ф., Смит Б. Влияние холестерина на структуру фосфолипидного бислоя. Проц Натл Академия наук США, 2009 г.; 106:3654-8.
- ^ Чен, Хэшунь; Родилась, Элла; Матур, Сатья Н.; Филд, Ф. Джеффри (1 декабря 1993 г.). «Синтез холестерина и сфингомиелина регулируется независимо в культивируемых клетках кишечника человека, CaCo-2: роль мембранного холестерина и содержания сфингомиелина» (PDF) . Журнал исследований липидов . 34 (12). Американское общество биохимии и молекулярной биологии: 2159–67. дои : 10.1016/S0022-2275(20)35356-6 . ISSN 0022-2275 . ПМИД 8301234 .
- ^ Ланг Т., Брунс Д., Венцель Д., Ридель Д., Холройд П., Тиле С., Ян Р. SNARE концентрируются в холестерин-зависимых кластерах, которые определяют места стыковки и слияния для экзоцитоза EMBO J 2001;20:2202-13.
Внешние ссылки
[ редактировать ]- Мембрана + липиды Национальной медицинской библиотеки США по медицинским предметным рубрикам (MeSH)