Префронтальная кора
| Префронтальная кора | |
|---|---|
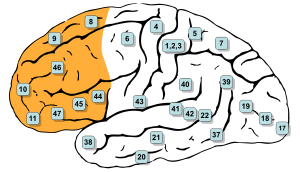 Области Бродмана 8, 9, 10, 11, 12, 13, 14, 24, 25, 32, 44, 45, 46 и 47 расположены в префронтальной коре. [1] | |
| Подробности | |
| Часть | Лобная доля |
| Части | Верхняя лобная извилина Средняя лобная извилина Нижняя лобная извилина |
| Артерия | Передняя мозговая Средний мозговой |
| вена | Верхний сагиттальный синус |
| Идентификаторы | |
| латинский | префронтальная кора |
| МеШ | D017397 |
| Нейроимена | 2429 |
| НейроЛекс ID | nlx_anat_090801 , ilx_0109209 |
| ФМА | 224850 |
| Анатомические термины нейроанатомии | |
В мозга млекопитающих анатомии префронтальная кора ( ПФК ) покрывает переднюю часть лобной доли коры головного мозга . Это ассоциативная кора лобной доли. [2] PFC содержит области Бродмана BA8 , BA9 , BA10 , BA11 , BA12 , BA13 , BA14 , BA24 , BA25 , BA32 , BA44 , BA45 , BA46 и BA47 . [1]
Эта область мозга участвует в широком спектре когнитивных функций более высокого порядка, включая формирование речи ( зона Брока ), взгляд ( лобные поля глаз ), рабочую память ( дорсолатеральная префронтальная кора ) и обработку рисков (например, вентромедиальная префронтальная кора ). Основной деятельностью этого отдела мозга считается оркестровка мыслей и действий в соответствии с внутренними целями. [3] Многие авторы указывают на неразрывную связь между волей человека к жизни, личностью и функциями префронтальной коры. [4]
Эта область мозга участвует в исполнительных функциях , таких как планирование , принятие решений , рабочая память , выражение личности, управление социальным поведением и контроль определенных аспектов речи и языка. [5] [6] [7] [8] Исполнительная функция связана со способностями различать конфликтующие мысли, определять хорошие и плохие, лучшие и лучшие, одинаковые и разные, будущие последствия текущей деятельности, работу над достижением определенной цели, прогнозирование результатов, ожидания, основанные на действиях, и социальный «контроль». (способность подавлять побуждения, которые, если их не подавить, могут привести к социально неприемлемым результатам).
Фронтальная кора поддерживает изучение конкретных правил, при этом большее количество передних областей поддерживает изучение правил на более высоких уровнях абстракции. [9]
Структура
[ редактировать ]Определение
[ редактировать ]Этот раздел необходимо обновить . ( март 2017 г. ) |
Есть три возможных способа определения префронтальной коры:
- как зернистая лобная кора
- зона проекции медиального дорсального ядра таламуса как
- как часть лобной коры, электрическая стимуляция которой не вызывает движений
Зернистая лобная кора
[ редактировать ]Префронтальная кора определяется на основании цитоархитектоники по наличию коркового зернистого слоя IV . Не совсем ясно, кто первым использовал этот критерий. Многие из ранних исследователей цитоархитектоники ограничивали использование термина «префронтальная» областью коры гораздо меньшего размера, включая прямую извилину и ростральную извилину ( Campbell , 1905; GE Smith , 1907; Brodmann , 1909; von Economo and Koskinas , 1925). Однако в 1935 году Якобсен использовал термин «префронтальная», чтобы отличить гранулярные префронтальные области от агранулярных моторных и премоторных областей. [10] Что касается областей Бродмана, префронтальная кора традиционно включает области 8, 9, 10, 11, 12, 13, 14, 24, 25, 32, 44, 45, 46 и 47. [1] однако не все эти области строго зернистые: 44 — дисгранулярные, каудальные 11 и орбитальные 47 — агранулярные. [11] Основная проблема с этим определением заключается в том, что оно хорошо работает только у приматов , но не у неприматов, поскольку у последних отсутствует зернистый слой IV. [12]
Зона проекции
[ редактировать ]Определение префронтальной коры как проекционной зоны ядра таламуса медиодорсального основано на работе Роуза и Вулси. [13] которые показали, что это ядро проецируется в переднюю и вентральную части мозга у неприматов, однако Роуз и Вулси назвали эту зону проекции «орбитофронтальной». Похоже, именно Акерт в 1964 году впервые прямо предположил, что этот критерий можно использовать для определения гомологов префронтальной коры у приматов и неприматов. [14] Это позволило установить гомологии, несмотря на отсутствие зернистой лобной коры у неприматов.
Определение проекционной зоны до сих пор широко распространено (например, Fuster [15] ), хотя его полезность подвергается сомнению. [11] [16] Современные исследования по отслеживанию путей показали, что проекции медиодорсального ядра таламуса не ограничиваются зернистой лобной корой у приматов. В результате было предложено определять префронтальную кору как область коры, имеющую более сильные реципрокные связи с медиодорсальным ядром, чем с любым другим ядром таламуса. [12] Уйлингс и др. [12] признаем, однако, что даже с применением этого критерия однозначно определить префронтальную кору может быть довольно сложно.
Электрически тихая область лобной коры
[ редактировать ]Третье определение префронтальной коры — это область лобной коры, электрическая стимуляция которой не приводит к заметным движениям. Например, в 1890 году Дэвид Феррье [17] использовал этот термин в этом смысле. Одна из сложностей с этим определением заключается в том, что электрически «тихая» лобная кора включает как зернистые, так и незернистые области. [11]
Подразделения
[ редактировать ]Этот раздел требует внимания эксперта в области нейробиологии . смотрите на странице обсуждения Подробности ( май 2019 г. ) |

По словам Стридтера, [18] ПФК человека можно разделить на две функционально, морфологически и эволюционно разные области: вентромедиальную ПФК (вмПФК), состоящую из:
- вентральная префронтальная кора (VPFC)
- медиальная префронтальная кора присутствует у всех млекопитающих (MPFC)
и латеральная префронтальная кора (LPFC), состоящая из:
- дорсолатеральная префронтальная кора (DLPFC)
- Вентролатеральная префронтальная кора (VLPFC) имеется только у приматов .
LPFC содержит области Бродмана BA8 , BA9 , BA10 , BA45 , BA46 и BA47 . Некоторые исследователи также включают BA44 . vmPFC содержит области Бродмана BA12 , BA25 , BA32 , BA33 , BA24 , BA11 , BA13 и BA14 .
В таблице ниже показаны различные способы разделения частей префронтальной коры человека на основе областей Бродмана. [1]
| 8 | 9 | 10 | 46 | 45 | 47 | 44 | 12 | 25 | 32 | 33 | 24 | 11 | 13 | 14 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| боковой | вентромедиальный | |||||||||||||
| дорсолатеральный | вентролатеральный | медиальный | вентральный | |||||||||||
- Дорсолатеральная префронтальная кора состоит из BA8 , BA9 , BA10 и BA46 . [1]
- Вентролатеральная префронтальная кора состоит из областей ВА45 , ВА47 и ВА44 . [1]
- Медиальная префронтальная кора (mPFC) состоит из BA12 , BA25 и передней поясной извилины : BA32 , BA33 , BA24 . [1] В этой области находится дорсальная связь , которая соединяет между собой многие части мозга. [19]
- Вентральная префронтальная кора состоит из областей ВА11 , ВА13 и ВА14 . [1] (См. также определение орбитофронтальной коры .)
- Дорсолатеральная префронтальная кора содержит ВА8 , включая лобные поля глаз . [1]
- Вентролатеральная префронтальная кора содержит BA45, который является частью зоны Брока . [20] Некоторые исследователи также относят BA44 к другой части зоны Брока.
Взаимосвязь
[ редактировать ]Префронтальная кора тесно связана с большей частью мозга, включая обширные связи с другими корковыми, подкорковыми участками и стволом мозга. [21] Дорсальная префронтальная кора особенно связана с областями мозга, отвечающими за внимание, познание и действие. [22] в то время как вентральная префронтальная кора связана с областями мозга, связанными с эмоциями. [23] Префронтальная кора также получает сигналы от систем возбуждения ствола мозга, и ее функция особенно зависит от нейрохимического окружения. [24] Таким образом, существует координация между состоянием возбуждения и психическим состоянием. [25] Взаимодействие между префронтальной корой и социально-эмоциональной системой мозга актуально для развития подростков, как предполагает модель дуальных систем .
Медиальная префронтальная кора участвует в генерации медленноволнового сна (SWS), а префронтальная атрофия связана с уменьшением SWS. [26] Префронтальная атрофия возникает естественным образом с возрастом человека, и было продемонстрировано, что у пожилых людей наблюдаются нарушения консолидации памяти по мере деградации их медиальной префронтальной коры. [26] У пожилых людей, вместо того, чтобы передаваться и храниться в неокортексе во время SWS, воспоминания начинают оставаться в гиппокампе, где они были закодированы , о чем свидетельствует повышенная активация гиппокампа по сравнению с более молодыми взрослыми во время задач по запоминанию , когда испытуемые учили словесные ассоциации, спали, а затем их попросили вспомнить выученные слова. [26]
Вентролатеральная префронтальная кора (VLPFC) участвует в различных аспектах производства речи и понимания языка. VLPFC тесно связана с различными областями мозга, включая латеральную и медиальную височную долю, верхнюю височную кору, подвисочную кору, периринальную кору и парагиппоккампальную кору. [27] Эти области мозга участвуют в восстановлении и консолидации памяти, обработке речи и ассоциации эмоций. Эти связи позволяют VLPFC опосредовать явное и неявное извлечение памяти и интегрировать его с языковыми стимулами, помогая планировать связную речь. [28] Другими словами, выбор правильных слов и умение оставаться «в теме» во время разговора исходит от VLPFC.
Функция
[ редактировать ]Исполнительная функция
[ редактировать ]Оригинальные исследования Фустера и Гольдмана-Ракича подчеркнули фундаментальную способность префронтальной коры представлять информацию, которой в данный момент нет в окружающей среде, и центральную роль этой функции в создании «мысленного блокнота для рисования». Гольдман-Ракич рассказала о том, как эти репрезентативные знания использовались для разумного управления мыслями, действиями и эмоциями, включая подавление неуместных мыслей, отвлекающих факторов, действий и чувств. [29] Таким образом, рабочую память можно рассматривать как фундаментальную основу внимания и поведенческого торможения. Фустер говорит о том, как эта префронтальная способность позволяет объединить прошлое с будущим, допуская как кросс-временные, так и кросс-модальные ассоциации в создании целенаправленных циклов восприятия-действия. [30] Эта способность к представительству лежит в основе всех остальных высших исполнительных функций.
Шимамура предложил теорию динамической фильтрации, чтобы описать роль префронтальной коры в управляющих функциях . Предполагается, что префронтальная кора действует как механизм пропускания или фильтрации высокого уровня, который усиливает целенаправленные активации и подавляет нерелевантные активации. Этот механизм фильтрации обеспечивает исполнительный контроль на различных уровнях обработки, включая выбор, поддержание, обновление и перенаправление активаций. Его также использовали для объяснения эмоциональной регуляции. [31]
Миллер и Коэн предложили интегративную теорию функции префронтальной коры, основанную на оригинальной работе Гольдмана-Ракича и Фустера. Они предполагают, что «когнитивный контроль возникает в результате активного поддержания паттернов активности в префронтальной коре, которая представляет цели и средства их достижения. Они подают предвзятые сигналы в другие структуры мозга, конечным эффектом которых является направление потока активности по нервным путям». которые устанавливают правильные сопоставления между входами, внутренними состояниями и выходами, необходимые для выполнения данной задачи». [32] По сути, они предполагают, что префронтальная кора управляет входными данными и связями, что позволяет когнитивно контролировать наши действия.
Префронтальная кора имеет большое значение, когда обработка информации сверху вниз необходима . Обработка сверху вниз по определению – это когда поведение определяется внутренними состояниями или намерениями. По их мнению, «ПФК имеет решающее значение в ситуациях, когда сопоставления между сенсорными входами, мыслями и действиями либо слабо установлены по сравнению с другими существующими, либо быстро меняются». [32] Примером этого может служить тест сортировки карточек штата Висконсин (WCST) . Испытуемым, выполняющим это задание, предлагается сортировать карточки по форме, цвету или количеству появляющихся на них символов. Идея состоит в том, что любая карта может быть связана с рядом действий, и ни одно сопоставление стимулов и реакций не будет работать. Люди с повреждением PFC могут сортировать карту при выполнении первоначальных простых задач, но не могут этого сделать, поскольку правила классификации меняются.
Миллер и Коэн приходят к выводу, что последствия их теории могут объяснить, какую роль играет ПФК в управлении когнитивными действиями. По словам самих исследователей, они утверждают, что «в зависимости от цели воздействия репрезентации в ПФК могут функционировать по-разному в качестве шаблонов, правил или целей внимания, передавая нисходящие сигналы предвзятости другим частям мозга, которые направляют процесс». поток активности по путям, необходимым для выполнения задачи». [32]
Экспериментальные данные указывают на роль префронтальной коры в обеспечении нормальной физиологии сна, сновидений и явлений депривации сна. [33]
При анализе и размышлении о признаках других людей активируется медиальная префронтальная кора, однако она не активируется при созерцании характеристик неодушевленных предметов. [34]
Исследования с использованием фМРТ показали, что медиальная префронтальная кора (mPFC), особенно передняя медиальная префронтальная кора (amPFC), может модулировать мимикрию. Нейробиологи предполагают, что социальный прайминг влияет на активность и обработку данных в amPFC и что эта область префронтальной коры модулирует мимикриальные реакции и поведение. [35]
Недавно исследователи использовали методы нейровизуализации и обнаружили, что наряду с базальными ганглиями префронтальная кора участвует в обучении образцов, что является частью теории образцов — одного из трех основных способов, которыми наш разум классифицирует вещи. Теория образцов утверждает, что мы классифицируем суждения, сравнивая их с аналогичным прошлым опытом в наших сохраненных воспоминаниях. [36]
Мета-анализ, проведенный в 2014 году профессором Николь П.Юань из Университета Аризоны, показал, что больший объем префронтальной коры и большая толщина коры ПФК связаны с лучшими исполнительными способностями. [37]
Внимание и память
[ редактировать ]
Широко распространенная теория о функции префронтальной коры головного мозга заключается в том, что она служит хранилищем кратковременной памяти . Эту идею впервые сформулировал Якобсен, который в 1936 году сообщил, что повреждение префронтальной коры приматов вызывает дефицит кратковременной памяти. [39] Карл Прибрам и коллеги (1952) определили часть префронтальной коры, ответственную за этот дефицит, как область 46 , также известную как дорсолатеральная префронтальная кора (dlPFC). [40] Совсем недавно Goldman-Rakic и коллеги (1993) вызвали кратковременную потерю памяти в локализованных областях пространства путем временной инактивации частей dlPFC. [41] концепцию рабочей памяти (см. также модель рабочей памяти Бэддели (1986) утвердил в современной нейробиологии После того, как Алан Бэддели ) , эти нейропсихологические открытия способствовали появлению теории о том, что префронтальная кора реализует рабочую память, а в некоторых крайних формулировках — только рабочую память. память. [42] В 1990-х годах у этой теории появилось множество последователей, и она стала преобладающей теорией функции PF, особенно у нечеловекообразных приматов. Концепция рабочей памяти, используемая сторонниками этой теории, сосредоточена в основном на кратковременном сохранении информации и в меньшей степени на манипулировании или мониторинге такой информации или на использовании этой информации для принятия решений. В соответствии с идеей о том, что префронтальная кора функционирует преимущественно в режиме поддержания памяти, активность периода задержки в ПФ часто интерпретируется как след памяти. (Фраза «активность периода задержки» применяется к активности нейронов, которая следует за временным представлением командного сигнала и сохраняется до последующего сигнала «начать» или «триггер».)
Чтобы изучить альтернативные интерпретации активности периода задержки в префронтальной коре, Лебедев и др. (2004) исследовали частоту разрядов одиночных префронтальных нейронов, когда обезьяны обращали внимание на стимул, отмечающий одно место, и запоминали другое, немаркированное место. [38] Оба места служили потенциальными объектами саккадического движения глаз . Хотя задача предъявляла интенсивные требования к кратковременной памяти, наибольшая часть префронтальных нейронов представляла собой посещаемые, а не запомнившиеся места. Эти результаты показали, что функции кратковременной памяти не могут объяснить всю или даже большую часть активности периода задержки в исследованной части префронтальной коры. Авторы предположили, что префронтальная активность в период задержки больше способствует процессу отбора внимания (и избирательного внимания ), чем сохранению памяти. [8] [43]
Речевое производство и язык
[ редактировать ]Различные области префронтальной коры задействованы во множестве важнейших функций, связанных с производством речи, пониманием языка и планированием реакции перед речью. [7] Когнитивная нейробиология показала, что левая вентролатеральная префронтальная кора жизненно важна для обработки слов и предложений.
Было обнаружено, что правая префронтальная кора отвечает за координацию извлечения явных воспоминаний для использования в речи, тогда как деактивация левой отвечает за опосредование извлечения неявных воспоминаний, которые будут использоваться при генерации глаголов. [7] Запоминание существительных (явная память) нарушено у некоторых пациентов с амнезией и повреждением правой префронтальной коры, но генерация глаголов остается неизменной, поскольку она зависит от дезактивации левой префронтальной коры. [28]
Многие исследователи теперь включают BA45 в префронтальную кору, поскольку вместе с BA44 он составляет область лобной доли, называемую зоной Брока . [20] Зона Брока широко считается выходной областью пути производства речи в мозге (в отличие от зоны Вернике в медиальной височной доле, которая рассматривается как область языкового ввода). Было показано, что BA45 участвует в извлечении соответствующих семантических знаний, которые будут использоваться в разговоре/речи. [7] Правая латеральная префронтальная кора (RLPFC) участвует в планировании сложного поведения и вместе с двусторонней BA45 поддерживает концентрацию и связность во время речи. [28] Однако было показано, что левый BA45 значительно активируется при сохранении связности речи у молодых людей. Было показано, что пожилые люди вербуют правильный BA45 чаще, чем их более молодые коллеги. [28] Это согласуется с данными об уменьшении латерализации в других системах мозга во время старения.
Кроме того, было показано, что это увеличение активности BA45 и RLPFC в сочетании с BA47 у пожилых пациентов способствует «высказываниям не по теме». Область BA47 в префронтальной коре участвует в «стимулированном» извлечении менее важных знаний, чем требуется для участия в разговоре. [28] Другими словами, было показано, что повышенная активация BA47 вместе с измененной активностью BA45 и более широкого RLPFC способствует включению менее релевантной информации и нерелевантных тангенциальных моделей разговорной речи у пожилых людей.
Клиническое значение
[ редактировать ]В последние несколько десятилетий системы визуализации мозга использовались для определения объемов областей мозга и нервных связей. В ряде исследований показано, что уменьшение объема и связей лобных долей с другими отделами мозга наблюдается у пациентов с психическими расстройствами ; те, кто подвергается повторяющимся стрессовым факторам ; [44] те, кто чрезмерно потребляет материалы откровенно сексуального характера; [45] самоубийства ; [46] преступники ; социопаты ; те, кто пострадал от отравления свинцом ; [47] Считается, что по крайней мере некоторые человеческие способности чувствовать вину или раскаяние, а также интерпретировать реальность зависят от хорошо функционирующей префронтальной коры. [48] Развитая нейросхема и функция саморегуляции префронтальной коры человека также связаны с более высокой чувствительностью у людей. [49] поскольку префронтальная кора у человека занимает гораздо больший процент мозга, чем у любого другого животного. Предполагается, что, поскольку за пять миллионов лет человеческой эволюции мозг увеличился втрое, [50] префронтальная кора увеличилась в размерах в шесть раз. [51]
Обзор исполнительных функций у здоровых людей, занимающихся спортом, показал, что левая и правая половины префронтальной коры, разделенной медиальной продольной щелью , становятся более взаимосвязанными в ответ на постоянные аэробные упражнения. [52] Два обзора исследований структурной нейровизуализации показывают, что заметное улучшение объема серого вещества префронтальной области и гиппокампа происходит у здоровых взрослых, которые выполняют упражнения средней интенсивности в течение нескольких месяцев. [53] [54]
Хроническое употребление алкоголя приводит к стойким изменениям в работе мозга, включая изменение способности принимать решения. Было показано, что префронтальная кора хронических алкоголиков уязвима к окислительному повреждению ДНК и гибели нейронов . [55]
История
[ редактировать ]Возможно, ярким примером нарушения функции префронтальной коры является случай Финеаса Гейджа , левая лобная доля которого была разрушена, когда в результате несчастного случая в 1848 году ему в голову проткнули большой железный стержень. Стандартное представление состоит в том, что, хотя Гейдж сохранил нормальную память, речь и моторику, его личность радикально изменилась: он стал раздражительным, вспыльчивым и нетерпеливым (характеристики, которые он раньше не проявлял), так что друзья описывали его как «больше не Гейджа». "; и, хотя раньше он был способным и эффективным работником, впоследствии он не смог завершить работу. [56] изменений Гейджа Однако тщательный анализ первичных данных показывает, что описания психологических обычно преувеличены, если сравнивать их с описанием, данным врачом Гейджа, причем наиболее поразительной особенностью является то, что изменения, описанные спустя годы после смерти Гейджа, гораздо более драматичны, чем все, что сообщалось при его жизни. . [57] [58]
Последующие исследования пациентов с префронтальными травмами показали, что пациенты вербально выражали наиболее подходящие социальные реакции в определенных обстоятельствах. Тем не менее, на самом деле, они вместо этого следовали поведению, направленному на немедленное удовлетворение, несмотря на то, что знали, что долгосрочные результаты будут обречены на провал.
Интерпретация этих данных показывает, что не только навыки сравнения и понимания конечных результатов заложены в префронтальной коре, но и префронтальная кора (при правильном функционировании) контролирует умственную возможность отложить немедленное удовлетворение ради лучшего или более полезного долгосрочного удовлетворения. результат. Эта способность ждать вознаграждения является одним из ключевых моментов, определяющих оптимальную исполнительную функцию человеческого мозга. [ нужна ссылка ]
В настоящее время проводится множество исследований, посвященных пониманию роли префронтальной коры при неврологических расстройствах. Начались клинические испытания некоторых препаратов, которые, как было доказано, улучшают функцию префронтальной коры, включая гуанфацин , который действует через адренергический рецептор альфа-2А . Последующая мишень этого препарата, канал HCN , является одной из новейших областей исследований в фармакологии префронтальной коры. [59]
Этимология
[ редактировать ]Термин «префронтальный отдел мозга», по-видимому, был введен Ричардом Оуэном в 1868 году. [10] По его мнению, префронтальная область ограничивалась самой передней частью лобной доли (приблизительно соответствующей лобному полюсу). Было высказано предположение, что его выбор термина был основан на префронтальной кости , присутствующей у большинства амфибий и рептилий. [10]
Дополнительные изображения
[ редактировать ]-
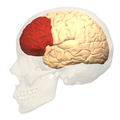
 Анимация, префронтальная кора левого полушария головного мозга (показана красным)
Анимация, префронтальная кора левого полушария головного мозга (показана красным) -
 Вид спереди
Вид спереди -
 Боковой вид
Боковой вид -
 Медиальная перспектива
Медиальная перспектива




См. также
[ редактировать ]- Лоботомия
- Теория интерференции
- Теория субъективности самомодели
- Дорсомедиальная префронтальная кора
- Вентромедиальная префронтальная кора
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я Мюррей Э., Уайз С., Грахатл К. (2016). «Глава 1: История систем памяти» . Эволюция систем памяти: предки, анатомия и адаптации (1-е изд.). Издательство Оксфордского университета. стр. 22–24. ISBN 978-0-19-150995-7 . Проверено 12 марта 2017 г.
- ^ «Префронтальная кора — обзор | Темы ScienceDirect» . www.sciencedirect.com . Проверено 8 февраля 2024 г.
- ^ Миллер Э.К., Фридман Д.Д., Уоллис Дж.Д. (август 2002 г.). «Префронтальная кора: категории, понятия и познание» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 357 (1424): 1123–1136. дои : 10.1098/rstb.2002.1099 . ПМК 1693009 . ПМИД 12217179 .
- ^ ДеЯнг К.Г., Хирш Дж.Б., Шейн М.С., Пападеметрис X, Радживан Н., Грей JR (июнь 2010 г.). «Проверка предсказаний нейробиологии личности. Структура мозга и большая пятерка» . Психологическая наука . 21 (6): 820–828. дои : 10.1177/0956797610370159 . ПМК 3049165 . ПМИД 20435951 .
- ^ Жоао, Рафаэль Батиста; Филгейрас, Ракель Маттос (03 октября 2018 г.), Старчевич, Ана; Филипович, Бранислав (ред.), «Лобная доля: функциональная нейроанатомия ее схем и связанные с ней синдромы отключения» , Префронтальная кора головного мозга , InTech, doi : 10.5772/intechopen.79571 , ISBN 978-1-78923-903-4 , получено 24 июня 2023 г.
- ^ Ян Ю, Рейн А. (ноябрь 2009 г.). «Результаты префронтальной структурной и функциональной визуализации мозга у антисоциальных, жестоких и психопатических людей: метаанализ» . Психиатрические исследования . 174 (2): 81–88. doi : 10.1016/j.pscychresns.2009.03.012 . ПМК 2784035 . ПМИД 19833485 .
- ^ Перейти обратно: а б с д Габриэли Дж. Д., Полдрак Р. А., Десмонд Дж. Э. (февраль 1998 г.). «Роль левой префронтальной коры в языке и памяти» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (3): 906–913. Бибкод : 1998PNAS...95..906G . дои : 10.1073/pnas.95.3.906 . ПМК 33815 . ПМИД 9448258 .
- ^ Перейти обратно: а б Балдауф Д., Дезимона Р. (апрель 2014 г.). «Нейронные механизмы объектного внимания» . Наука . 344 (6182): 424–427. Бибкод : 2014Sci...344..424B . дои : 10.1126/science.1247003 . ПМИД 24763592 . S2CID 34728448 .
- ^ Бадре Д., Кайзер А.С., Д'Эспозито М. (апрель 2010 г.). «Фронтальная кора и открытие абстрактных правил действия» . Нейрон . 66 (2): 315–326. дои : 10.1016/j.neuron.2010.03.025 . ПМК 2990347 . ПМИД 20435006 .
- ^ Перейти обратно: а б с Палец С (1994). Истоки нейробиологии: история исследований функций мозга . Оксфорд [Оксфордшир]: Издательство Оксфордского университета. ISBN 978-0-19-514694-3 . [ нужна страница ]
- ^ Перейти обратно: а б с Прейсс ТМ (1995). «Есть ли у крыс префронтальная кора? Программа Роуз-Вулси-Акерт пересмотрена». Журнал когнитивной нейронауки . 7 (1): 1–24. дои : 10.1162/jocn.1995.7.1.1 . ПМИД 23961750 . S2CID 2856619 .
- ^ Перейти обратно: а б с Уилингс Х.Б., Гроеневеген Х.Дж., Колб Б. (ноябрь 2003 г.). «Есть ли у крыс префронтальная кора?». Поведенческие исследования мозга . 146 (1–2): 3–17. дои : 10.1016/j.bbr.2003.09.028 . ПМИД 14643455 . S2CID 32136463 .
- ^ Роуз Дж. Э., Вулси CN (1948). «Орбитофронтальная кора и ее связи с медиодорсальным ядром у кролика, овцы и кошки». Научные публикации – Ассоциация исследований нервных и психических заболеваний . 27 : 210–232. ПМИД 18106857 .
- ^ Пройсс Т.М., Гольдман-Ракич П.С. (август 1991 г.). «Миело- и цитоархитектура зернистой лобной коры и окружающих областей у стрепсиринового примата Галаго и человекообразного примата Макака». Журнал сравнительной неврологии . 310 (4): 429–474. дои : 10.1002/cne.903100402 . ПМИД 1939732 . S2CID 34575725 .
- ^ Фустер Дж. М. (2008). Префронтальная кора (4-е изд.). Бостон: Академическая пресса. ISBN 978-0-12-373644-4 . [ нужна страница ]
- ^ Маркович Х.Дж. , Притцель М. (1979). «Префронтальная кора: область проекции медиодорсального ядра таламуса?» . Физиологическая психология . 7 (1): 1–6. дои : 10.3758/bf03326611 .
- ^ Ферье Д. (июнь 1890 г.). «Крунские лекции по церебральной локализации» . Британский медицинский журнал . 1 (1537): 1349–1355. дои : 10.1136/bmj.1.1537.1349 . ПМК 2207859 . ПМИД 20753055 .
- ^ Стридтер Г.Ф. (2005). Принципы эволюции мозга . Синауэр Ассошиэйтс. ISBN 978-0-87893-820-9 .
- ^ Шелин, Иветт; Ян, Шизи (2010). «Функциональная МРТ в состоянии покоя при депрессии выявляет усиление связи между сетями через дорсальную связь» . ПНАС . 107 (24): 11020–11025. Бибкод : 2010PNAS..10711020S . дои : 10.1073/pnas.1000446107 . ПМЦ 2890754 . ПМИД 20534464 .
- ^ Перейти обратно: а б «Зона Брока | анатомия» . Британская энциклопедия . Проверено 12 декабря 2019 г.
- ^ Альварес Х.А., Эмори Э. (март 2006 г.). «Исполнительная функция и лобные доли: метааналитический обзор». Обзор нейропсихологии . 16 (1): 17–42. дои : 10.1007/s11065-006-9002-x . ПМИД 16794878 . S2CID 207222975 .
- ^ Гольдман-Ракич П.С. (1988). «Топография познания: параллельные распределенные сети в ассоциативной коре приматов». Ежегодный обзор неврологии . 11 : 137–156. дои : 10.1146/annurev.ne.11.030188.001033 . ПМИД 3284439 .
- ^ Прайс JL (июнь 1999 г.). «Префронтальные корковые сети, связанные с висцеральными функциями и настроением». Анналы Нью-Йоркской академии наук . 877 (1): 383–396. Бибкод : 1999NYASA.877..383P . дои : 10.1111/j.1749-6632.1999.tb09278.x . ПМИД 10415660 . S2CID 37564764 .
- ^ Роббинс Т.В., Арнстен А.Ф. (2009). «Нейропсихофармакология лобно-исполнительной функции: моноаминергическая модуляция» . Ежегодный обзор неврологии . 32 : 267–287. дои : 10.1146/annurev.neuro.051508.135535 . ПМЦ 2863127 . ПМИД 19555290 .
- ^ Арнстен А.Ф., Паспалас К.Д., Гамо Нью-Джерси, Ян Ю, Ван М. (август 2010 г.). «Динамическая сетевая связь: новая форма нейропластичности» . Тенденции в когнитивных науках . 14 (8): 365–375. дои : 10.1016/j.tics.2010.05.003 . ПМК 2914830 . ПМИД 20554470 .
- ^ Перейти обратно: а б с Мандер Б.А., Рао В., Лу Б., Салетин Дж.М., Линдквист Дж.Р., Анколи-Исраэль С. и др. (март 2013 г.). «Префронтальная атрофия, нарушение медленных волн медленного сна и нарушение памяти, зависящей от гиппокампа, при старении» . Природная неврология . 16 (3): 357–364. дои : 10.1038/nn.3324 . ПМК 4286370 . ПМИД 23354332 .
- ^ Куль Б.А., Вагнер А.Д. (1 января 2009 г.). «Стратегический контроль памяти». В Squire LR (ред.). Энциклопедия неврологии . Академическая пресса. стр. 437–444. дои : 10.1016/b978-008045046-9.00424-1 . ISBN 978-0-08-045046-9 .
- ^ Перейти обратно: а б с д и Хоффман П. (январь 2019 г.). «Снижение активности префронтальных отделов предсказывает появление не по теме высказываний во время произнесения речи» . Природные коммуникации . 10 (1): 515. Бибкод : 2019NatCo..10..515H . дои : 10.1038/s41467-019-08519-0 . ПМК 6355898 . ПМИД 30705284 .
- ^ Гольдман-Ракич П.С. (октябрь 1996 г.). «Префронтальный ландшафт: значение функциональной архитектуры для понимания человеческого мышления и центрального исполнительного органа». Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 351 (1346): 1445–1453. дои : 10.1098/rstb.1996.0129 . JSTOR 3069191 . ПМИД 8941956 .
- ^ Фустер Дж. М., Боднер М., Крогер Дж. К. (май 2000 г.). «Кросс-модальная и кросс-временная ассоциация в нейронах лобной коры». Природа . 405 (6784): 347–351. Бибкод : 2000Natur.405..347F . дои : 10.1038/35012613 . ПМИД 10830963 . S2CID 4421762 .
- ^ Шимамура А.П. (2000). «Роль префронтальной коры в динамической фильтрации» . Психобиология . 28 (2): 207–218. дои : 10.3758/BF03331979 . S2CID 140274181 .
- ^ Перейти обратно: а б с Миллер Э.К., Коэн Дж.Д. (2001). «Интегративная теория функции префронтальной коры». Ежегодный обзор неврологии . 24 : 167–202. дои : 10.1146/annurev.neuro.24.1.167 . ПМИД 11283309 . S2CID 7301474 .
- ^ Музур А., Пейс-Шотт Э.Ф., Хобсон Дж.А. (ноябрь 2002 г.). «Префронтальная кора во сне». Тенденции в когнитивных науках . 6 (11): 475–481. дои : 10.1016/S1364-6613(02)01992-7 . ПМИД 12457899 . S2CID 5530174 .
- ^ Митчелл Дж.П., Хизертон Т.Ф., Макрей К.Н. (ноябрь 2002 г.). «Различные нейронные системы служат знанием человека и объекта» . Труды Национальной академии наук Соединенных Штатов Америки . 99 (23): 15238–15243. Бибкод : 2002PNAS...9915238M . дои : 10.1073/pnas.232395699 . ПМЦ 137574 . ПМИД 12417766 .
- ^ Ван Ю, Hamilton AF (апрель 2015 г.). «Передняя медиальная префронтальная кора реализует социальную настройку мимикрии» . Социальная когнитивная и аффективная нейронаука . 10 (4): 486–493. дои : 10.1093/scan/nsu076 . ПМЦ 4381231 . ПМИД 25009194 .
- ^ Шактер, Дэниел Л., Дэниел Тодд Гилберт и Дэниел М. Вегнер. Психология. 2-е изд., страницы 364–366. Нью-Йорк, штат Нью-Йорк: Worth Publishers, 2011. Печать.
- ^ Юань П., Раз Н. (май 2014 г.). «Префронтальная кора и исполнительные функции у здоровых взрослых: метаанализ структурных исследований нейровизуализации» . Неврологические и биоповеденческие обзоры . 42 : 180–192. doi : 10.1016/j.neubiorev.2014.02.005 . ПМК 4011981 . ПМИД 24568942 .
- ^ Перейти обратно: а б Лебедев М.А., Мессингер А, Кралик Дж.Д., Уайз С.П. (ноябрь 2004 г.). «Представление посещенных и запомненных мест в префронтальной коре» . ПЛОС Биология . 2 (11): е365. дои : 10.1371/journal.pbio.0020365 . ПМЦ 524249 . ПМИД 15510225 .

- ^ Якобсен CF (1936) Исследования мозговой функции у приматов. I. Функции лобных ассоциативных зон у обезьян. Comp Psychol Monogr 13: 3–60.
- ^ Прибрам К.Х., Мишкин М., Росволд Х.Э., Каплан С.Дж. (декабрь 1952 г.). «Влияние на эффективность отсроченного ответа при поражениях дорсолатеральной и вентромедиальной лобной коры бабуинов». Журнал сравнительной и физиологической психологии . 45 (6): 565–575. дои : 10.1037/h0061240 . ПМИД 13000029 .
- ^ Фунахаши С., Брюс С.Дж., Голдман-Ракич П.С. (апрель 1993 г.). «Дорсолатеральные префронтальные поражения и глазодвигательная реакция с задержкой реакции: свидетельства мнемонических «скотом» » . Журнал неврологии . 13 (4): 1479–1497. doi : 10.1523/JNEUROSCI.13-04-01479.1993 . ПМК 6576716 . ПМИД 8463830 .
- ^ Баддели А. (1986) Рабочая память. Оксфорд: Издательство Оксфордского университета. стр.289
- ^ Бедини М., Балдауф Д. (август 2021 г.). «Отпечатки структуры, функций и связей лобного поля глаза по сравнению с нижним лобным соединением: всестороннее сравнение» . Европейский журнал неврологии . 54 (4): 5462–5506. дои : 10.1111/ejn.15393 . ПМЦ 9291791 . ПМИД 34273134 . S2CID 235999643 .
- ^ Листон С., Миллер М.М., Голдуотер Д.С., Рэдли Дж.Дж., Рошер А.Б., Хоф П.Р. и др. (июль 2006 г.). «Вызванные стрессом изменения в морфологии дендритов префронтальной коры предсказывают избирательные нарушения перцептивного переключения внимания» . Журнал неврологии . 26 (30): 7870–7874. doi : 10.1523/JNEUROSCI.1184-06.2006 . ПМК 6674229 . ПМИД 16870732 .
- ^ «У зрителей порнографии меньшая система вознаграждений» . МАКС-ПЛАНК-ГЕСЕЛЬШАФТ . 2 июня 2014 года . Проверено 2 июля 2018 г.
- ^ Райковска Г. (декабрь 1997 г.). «Морфометрические методы исследования префронтальной коры у суицидников и психиатрических больных». Анналы Нью-Йоркской академии наук . 836 (1): 253–268. Бибкод : 1997NYASA.836..253R . дои : 10.1111/j.1749-6632.1997.tb52364.x . ПМИД 9616803 . S2CID 32947726 .
- ^ Сесил К.М., Брубейкер С.Дж., Адлер С.М., Дитрих К.Н., Алтай М., Эгельхофф Дж.К. и др. (май 2008 г.). Бальмес Дж. (ред.). «Уменьшение объема мозга у взрослых, подвергшихся воздействию свинца в детстве» . ПЛОС Медицина . 5 (5): е112. doi : 10.1371/journal.pmed.0050112 . ПМЦ 2689675 . ПМИД 18507499 .
- ^ Андерсон С.В., Бечара А., Дамасио Х., Транел Д., Дамасио А.Р. (ноябрь 1999 г.). «Нарушение социального и морального поведения, связанное с ранним повреждением префронтальной коры человека». Природная неврология . 2 (11): 1032–1037. дои : 10.1038/14833 . ПМИД 10526345 . S2CID 204990285 .
- ^ Фариба К., Гокараконда С.Б. (2021). «Расстройства контроля над импульсами» . СтатПерлс . Остров сокровищ (Флорида): StatPearls Publishing. ПМИД 32965950 . Проверено 4 мая 2021 г.
- ^ Шенеманн П.Т., Будингер Т.Ф., Сарич В.М., Ван В.С. (апрель 2000 г.). «Размер мозга не влияет на общие когнитивные способности в семье» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (9): 4932–4937. Бибкод : 2000PNAS...97.4932S . дои : 10.1073/pnas.97.9.4932 . ЧВК 18335 . ПМИД 10781101 .
- ^ Касцио Т. «Дом и психология, серия 14» . Психология сегодня . Архивировано из оригинала 8 апреля 2013 г. Проверено 15 ноября 2011 г.
- ^ Гини Х., Мачадо Л. (февраль 2013 г.). «Польза регулярных аэробных упражнений для исполнительной деятельности у здорового населения» . Психономический бюллетень и обзор . 20 (1): 73–86. дои : 10.3758/s13423-012-0345-4 . ПМИД 23229442 . S2CID 24190840 .
- ^ Эриксон К.И., Леки Р.Л., Вайнштейн А.М. (сентябрь 2014 г.). «Физическая активность, физическая форма и объем серого вещества» . Нейробиология старения . 35 (Приложение 2): С20–С28. doi : 10.1016/j.neurobiolaging.2014.03.034 . ПМК 4094356 . ПМИД 24952993 .
- ^ Валканова В., Эгия Родригес Р., Эбмайер К.П. (июнь 2014 г.). «Разум превыше материи – что мы знаем о нейропластичности у взрослых?». Международная психогериатрия . 26 (6): 891–909. дои : 10.1017/S1041610213002482 . ПМИД 24382194 . S2CID 20765865 .
- ^ Фаулер А.К., Томпсон Дж., Чен Л., Дагда М., Дертьен Дж., Доссу К.С. и др. (2014). «Дифференциальная чувствительность префронтальной коры и гиппокампа к алкогольной токсичности» . ПЛОС ОДИН . 9 (9): e106945. Бибкод : 2014PLoSO...9j6945F . дои : 10.1371/journal.pone.0106945 . ПМЦ 4154772 . ПМИД 25188266 .
- ^ Антонио Дамасио , Ошибка Декарта . Паб «Пингвин Путман» , 1994 год . [ нужна страница ]
- ^ Малкольм Макмиллан, Странная слава: истории Финеаса Гейджа (MIT Press, 2000), стр. 116–119, 307–333, особенно. стр. 11,333.
- ^ Макмиллан М (2008). «Финес Гейдж – Разгадывая миф» . Психолог . 21 (9). Британское психологическое общество : 828–831. Архивировано из оригинала 3 сентября 2010 г. Проверено 21 июня 2014 г.
- ^ Ван М., Рамос Б.П., Паспалас К.Д., Шу Ю, Симен А., Дуке А. и др. (апрель 2007 г.). «Альфа2А-адренорецепторы укрепляют сети рабочей памяти, ингибируя передачу сигналов канала цАМФ-HCN в префронтальной коре» . Клетка . 129 (2): 397–410. дои : 10.1016/j.cell.2007.03.015 . ПМИД 17448997 . S2CID 741677 .