Рибосомальный съемник кадров
Рибосомальное рамки , также известное как трансляционное кадрирование или перекодирование трансляции , является биологическим явлением, которое происходит во время трансляции , которое приводит к производству множества уникальных белков из одной мРНК . [ 1 ] Процесс может быть запрограммирован с помощью нуклеотидной последовательности мРНК и иногда влияет вторичная трехмерная структура мРНК . [ 2 ] Он был описан в основном в вирусах (особенно ретровирусах ), ретротранспозонах и элементах бактериальной вставки, а также в некоторых клеточных генах . [ 3 ]
Также было обнаружено, что мелкие молекулы, белки и нуклеиновые кислоты стимулируют уровни фрейм -съемки. В декабре 2023 года сообщалось, что , транскрибированные in vitro (IVT) мРНК в ответ на BNT162B2 (Pfizer-Biontech) против ков-19 вакцину, вызвала рибосомальный фрейм. [ 4 ]
Обзор процесса
[ редактировать ]Белки транслируются с чтением тринуклеотидов на цепи мРНК, также известной как кодоны , от одного конца мРНК до другой (от 5 ' до 3' конца ), начиная с аминокислотного метионина в качестве начала (инициация) Кодон август. Каждый кодон переводится в одну аминокислоту . Сам код считается вырожденным , что означает, что конкретная аминокислота может быть указана более чем одним кодоном. Однако сдвиг любого количества нуклеотидов, который не делится на 3 в рамке считывания, приведет к чтению последующих кодонов по -разному. [ 5 ] Это эффективно меняет рибосомальный кадр считывания .
Пример предложения
[ редактировать ]В этом примере следующее предложение трехбуквенных слов имеет смысл при чтении с самого начала:
|Start|THE CAT AND THE MAN ARE FAT ... |Start|123 123 123 123 123 123 123 ...
Однако, если кадр считывания смещается на одну букву на между T и H первого слова (эффективно FrameShift +1 при рассмотрении положения 0 является начальной позицией T ),
T|Start|HEC ATA NDT HEM ANA REF AT... -|Start|123 123 123 123 123 123 12...
Затем предложение читается по -другому, не имеет смысла.
ДНК -пример
[ редактировать ]В этом примере следующая последовательность представляет собой область митохондриального генома человека с двумя перекрывающимися генами MT-ATP8 и MT-ATP6 . При чтении с самого начала эти кодоны имеют смысл для рибосомы и могут быть переведены в аминокислоты (АА) под кодом митохондриального кода позвоночных :
|Start|AAC GAA AAT CTG TTC GCT TCA ... |Start|123 123 123 123 123 123 123 ... | AA | N E N L F A S ...
Тем не менее, давайте изменим кадр считывания, запустив один нуклеотид вниз по течению (фактически «+1 FrameShift» при рассмотрении положения 0 является начальной позицией A )::
A|Start|ACG AAA ATC TGT TCG CTT CA... -|Start|123 123 123 123 123 123 12... | AA | T K I C S L ...
Из -за этого +1 -фраместика последовательность ДНК читается по -разному. Поэтому различные кадры считывания кодонов дают разные аминокислоты.
Эффект
[ редактировать ]В случае транспортировки рибосомы, смягчение кадров может привести к бессмысленной мутации , преждевременному стоп -кодону после кадрирования, либо к созданию совершенно нового белка после фрейм. В случае, когда фрейм-съемка приводит к бессмысленной бессмысленности, путь, опосредованный бессмысленностью фреймэрией служит методом регуляции уровня экспрессии , может разрушить транскрипт мРНК, поэтому сэмпение с связанного гена. [ 6 ]
Если производится новый или нецелевой белок, он может вызвать другие неизвестные последствия. [ 4 ]
Функционировать в вирусах и эукариотах
[ редактировать ]В вирусах это явление может быть запрограммирован на наличие на определенных участках и позволяет вирусу кодировать несколько типов белков из одной и той же мРНК. Примечательные примеры включают ВИЧ-1 (вирус иммунодефицита человека), [ 7 ] RSV ( вирус саркомы ) [ 8 ] и вирус гриппа (грипп), [ 9 ] которые все полагаются на кадрирование, чтобы создать правильное соотношение белков с рамой (нормальное трансляция) и «транс-рама» (кодируется фрейм-съемной последовательности). Его использование в вирусах в первую очередь для уплотнения более генетической информации в более короткое количество генетического материала.
У эукариот он, по -видимому, играет роль в регуляции уровней экспрессии генов путем создания преждевременных остановок и создания нефункциональных транскриптов. [ 3 ] [ 10 ]
Типы фреймэфира
[ редактировать ]Наиболее распространенным типом кадрирования является -1 фрейм -съемки или запрограммирован -1 рибосомальный фрамезирование (-1 PRF) . Другие, более редкие типы фрейм -съемки включают в себя +1 и -2. [ 2 ] Считается, что −1 и +1 -сэмпение контролируются различными механизмами, которые обсуждаются ниже. Оба механизма связаны с кинетически .
Запрограммирован -1 рибосомный кадр
[ редактировать ]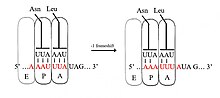
В −1 рамки рибосома проскальзывает один нуклеотид и продолжает трансляцию в рамке -1. Обычно существует три элемента, которые включают сигнал сдвига кадров -1: скользкая последовательность , область проставки и вторичная структура РНК. Скользкая последовательность соответствует мотиву x_xxy_yyh, где XXX-это три идентичных нуклеотида (хотя некоторые исключения происходят), Yyy обычно представляет UUU или AAA, а H-C или U. Поскольку структура этого мотива содержит 2 соседнего 3-нуклеотида. Повторения считается, что -1 фрамезирование описывается моделью тандемного проскальзывания, в которой рибосомальная P-сайта TRNA Anticodon переосмысливает от XXY в XXX и анти-сайт-анти-анти-картины от YYH до YYY одновременно. Эти новые пары идентичны парам 0-рамы, за исключением их третьих позиций. Эта разница не имеет существенного различия антикодонского связывания, потому что третий нуклеотид в кодоне, известный как положение колебания , имеет более слабую специфичность связывания антикодона тРНК, чем у первого и второго нуклеотидов. [ 2 ] [ 11 ] В этой модели структура мотива объясняется тем фактом, что первое и второе положение антикодонов должно быть в состоянии идеально сочетаться как в кадрах 0, так и в -1. Следовательно, нуклеотиды 2 и 1 должны быть идентичными, а нуклеотиды 3 и 2 также должны быть идентичны, что приводит к необходимой последовательности из 3 идентичных нуклеотидов для каждой тРНК, которая проскальзывает. [ 12 ]
+1 рибосомальный кадр
[ редактировать ]Слительная последовательность для сигнала +1 FrameShift не имеет одинакового мотива и вместо этого, по -видимому, функционирует, приостанавливая рибосому в последовательности, кодирующей редкую аминокислоту. [ 13 ] Рибосомы не транслируют белки с устойчивой скоростью, независимо от последовательности. нет равных количеств тРНК этого конкретного кодона Для перевода определенных кодонов требуется больше времени, потому что в цитозоле . [ 14 ] Из -за этого отставания существует в небольших участках последовательностей кодонов, которые контролируют скорость рибосомального кадрирования. В частности, рибосома должна остановиться, чтобы дождаться прибытия редкой тРНК, и это увеличивает кинетическую благоприятность рибосомы и связанной с ними тРНК, скользящей в новой раме. [ 13 ] [ 15 ] В этой модели изменение в рамке считывания вызвано одним скольжением тРНК, а не двумя.
Контролирующие механизмы
[ редактировать ]Рибосомальный фрейм-съемник может контролироваться механизмами, обнаруженными в последовательности мРНК (цис-действие). Обычно это относится к скользкой последовательности, вторичной структуре РНК или обоим. Сигнал сдвига кадров состоит из обоих элементов, разделенных областью проставки, как правило, длиной 5–9 нуклеотидов. [ 2 ] Фрамерирование также может быть индуцировано другими молекулами, которые взаимодействуют с рибосомой или мРНК (транс-действия).
Элементы сигнала FrameShift
[ редактировать ]
Скользкая последовательность
[ редактировать ]Слитель -последовательности потенциально могут сделать рибосому для чтения «скольжение» и пропустить несколько нуклеотидов (обычно только 1) и после этого считать совершенно другую кадр. В запрограммированном −1 рибосомном рамке скользкая последовательность соответствует мотиву x_xxy_yyh, где XXX - это три идентичных нуклеотида (хотя некоторые исключения происходят), yyy обычно представляет UUU или AAA, а H - A, C или U. в случае +. 1 Фрамерирование, скользкая последовательность содержит кодоны, для которых соответствующая тРНК встречается более редко, а смягчение кадров предпочтительнее, потому что кодон в новой кадре имеет более распространенную ассоциированную тРНК. [ 13 ] Одним из примеров скользкой последовательности является полиа на мРНК, которая, как известно, индуцирует проскальзывание рибосомы даже при отсутствии каких -либо других элементов. [ 16 ]
РНК вторичная структура
[ редактировать ]Эффективное рибосомальное рамки, как правило, требует присутствия вторичной структуры РНК, чтобы усилить эффекты скользкой последовательности. [ 12 ] структура РНК (которая может быть петлей ствола или псевдокнота Считается, что ), приостанавливает рибосому на скользком месте во время трансляции, заставляя ее переехать и продолжать репликацию из положения -1. Считается, что это происходит потому, что структура физически блокирует движение рибосомы, застряв в туннеле мРНК рибосомы. [ 2 ] Эта модель подтверждается тем фактом, что сила псевдокнота положительно коррелировала с уровнем рамки для ассоциированной мРНК. [ 3 ] [ 17 ]
Ниже приведены примеры прогнозируемых вторичных структур для элементов сдвига кадров, которые, как показано, стимулируют кадрирование в различных организмах. Большинство показанных структур представляют собой петли стеблей, за исключением структуры псевдоплики Alil (апикальная петля-петля). На этих изображениях более крупные и неполные круги мРНК представляют линейные области. Вторичные структуры «петли ствола», где «стебли» образуются в области спаривания основания мРНК с другой областью на той же цепи, показаны выступающими из линейной ДНК. Линейная область сигнала рибосомального рибосом ВИЧ содержит высококонсервативную uuu Uuu скользкую последовательность; Многие другие прогнозируемые структуры также содержат кандидатов для скользких последовательностей.
Последовательности мРНК на изображениях могут быть прочитаны в соответствии с набором руководящих принципов. В то время как A, T, C и G представляют собой конкретный нуклеотид в положении, есть также буквы, которые представляют собой двусмысленность, которые используются, когда в этом положении может возникнуть более одного вида нуклеотида. Правила Международного союза чистой и прикладной химии ( IUPAC ) следующие: [ 18 ]
| Символ [ 18 ] | Описание | Базы представлены | Дополнение | ||||
|---|---|---|---|---|---|---|---|
| А | Денин | А | 1 | Т | |||
| В | C Ytosine | В | Глин | ||||
| Глин | Гуанин | Глин | В | ||||
| Т | T HYMINE | Т | А | ||||
| В | U racil | В | А | ||||
| В | Слабый | А | Т | 2 | В | ||
| С | Сильный | В | Глин | С | |||
| М | M INO | А | В | K | |||
| K | K Eto | Глин | Т | М | |||
| Ведущий | pu r ine | А | Глин | Ведущий | |||
| И | P y rimidine | В | Т | И | |||
| Беременный | Не ( B приходит после а) | В | Глин | Т | 3 | V | |
| Дюймовый | не C ( D приходит после C) | А | Глин | Т | ЧАС | ||
| ЧАС | не г ( ч приходит после г) | А | В | Т | Дюймовый | ||
| V | не t ( v приходит после t и u) | А | В | Глин | Беременный | ||
| Не | любой N Ucleotide (не разрыв) | А | В | Глин | Т | 4 | Не |
| С | Z ero | 0 | С | ||||
Эти символы также действительны для РНК, за исключением U (uracil), заменив T (тимин). [ 18 ]
Галерея изображений вторичной структуры | |
|---|---|
|




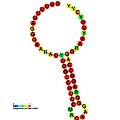


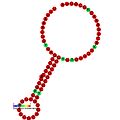




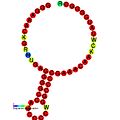

| Тип | Распределение | Рефери |
|---|---|---|
| АЛИЛ ПСЕУДОКНОТ | Бактерии | [ 19 ] |
| Элемент стимуляции РНК с антизимной РНК | Беспозвоночные | [ 20 ] |
| Коронавирус -каркасный элемент стимуляции | Coronavirus | [ 21 ] |
| DNAX Рибосомальный кадритный элемент | Эукариота , бактерии | [ 22 ] |
| Сигнал рибосомального сдвига ВИЧ | Вирусы | |
| Последовательность вставки IS1222 Рибосомальный элемент рамки | Эукариота , бактерии | |
| Рибосомальный съемник кадров | Вирусы |
Транс-действия элементы
[ редактировать ]Было обнаружено, что мелкие молекулы, белки и нуклеиновые кислоты стимулируют уровни фрейм -съемки. Например, механизм негативной петли обратной связи в пути синтеза полиаминов основан на уровнях полиаминов, стимулирующих увеличение +1 фреймэффирта, что приводит к производству ингибирующего фермента . Также было показано, что определенные белки, которые необходимы для распознавания кодона или которые связываются непосредственно с последовательности мРНК, модулируют уровни фреймэширования. Молекулы микроРНК (miRNA) могут гибридизоваться во вторичную структуру РНК и влиять на ее прочность. [ 6 ]
Смотрите также
[ редактировать ]- Элемент стимуляции РНК с антизимной РНК
- Коронавирус -каркасный элемент стимуляции
- DNAX Рибосомальный кадритный элемент
- Мутация сдвига кадров
- Сигнал рибосомального сдвига ВИЧ
- Последовательность вставки IS1222 Рибосомальный элемент рамки
- Recode Database
- Рибосомальная пауза
- Скользкая последовательность
Ссылки
[ редактировать ]- ^ Atkins JF, Loughran G, Bhatt PR, Firth AE, Baranov PV (сентябрь 2016 г.). «Рибосомальный фармэрией и транскрипционное проскальзывание: от генетической стеганографии и криптографии до Adventious использования» . Исследование нуклеиновых кислот . 44 (15): 7007–7078. doi : 10.1093/nar/gkw530 . PMC 5009743 . PMID 27436286 .
- ^ Jump up to: а беременный в дюймовый и Napthine S, Ling R, Finch LK, Jones JD, Bell S, Brierley I, Firth AE (июнь 2017 г.). «Белок-направленный рибосомальный фрейм-съемка временно регулирует экспрессию генов» . Природная связь . 8 : 15582. Bibcode : 2017natco ... 815582n . doi : 10.1038/ncomms15582 . PMC 5472766 . PMID 28593994 .
- ^ Jump up to: а беременный в Кетлер Р. (2012). «О запрограммированном рибосомном рамке: альтернативные протеомы» . Границы в генетике . 3 : 242. DOI : 10.3389/fgene.2012.00242 . PMC 3500957 . PMID 23181069 .
- ^ Jump up to: а беременный Mulroney, Thomas E.; Pöyry, Tuija; Ям-Пук, Хуан Карлос; Руст, Мария; Харви, Роберт Ф.; Калмар, Лахос; Хорнер, Эмили; Бут, Люси; Феррейра, Александр П.; Стонели, Марк; Савакар, Риттвик; Mentzer, Alexander J.; Лилли, Кэтрин С.; Smales, C. Марк; фон дер Хаар, Тобиас (6 декабря 2023 г.). «N1-метилпсеудуридилирование мРНК вызывает +1 рибосомальное кадрирование» . Природа . 625 (7993): 189–194. doi : 10.1038/s41586-023-06800-3 . ISSN 1476-4687 . PMC 10764286 . PMID 38057663 .
- ^ Иванов И.П., Аткинс Дж.Ф. (2007). «Рибосомальное рамение в декодирующих мРНК антизима от дрожжей и протистов до людей: около 300 случаев выявляют замечательное разнообразие, несмотря на основное сохранение» . Исследование нуклеиновых кислот . 35 (6): 1842–1858. doi : 10.1093/nar/gkm035 . PMC 1874602 . PMID 17332016 .
- ^ Jump up to: а беременный Dever TE, Dinman JD, Green R (август 2018 г.). «Удлинение перевода и перекодирование у эукариот» . Перспективы Cold Spring Harbor в биологии . 10 (8): A032649. doi : 10.1101/cshperspect.a032649 . PMC 6071482 . PMID 29610120 .
- ^ Jump up to: а беременный Jacks T, Power MD, Masiarz FR, Luciw PA, Barr PJ, Varmus He (январь 1988 г.). «Характеристика рибосомального фрейм-сшивания в экспрессии gag-pol ВИЧ-1». Природа . 331 (6153): 280–283. Bibcode : 1988natur.331..280j . doi : 10.1038/331280A0 . PMID 2447506 . S2CID 4242582 .
- ^ Jump up to: а беременный Джекс Т., Мадхани Х.Д., Масиарц Ф.Р., Вармус Хе (ноябрь 1988 г.). «Сигналы для рибосомального рамки в районе gag-pol вируса саркомы» . Клетка . 55 (3): 447–458. doi : 10.1016/0092-8674 (88) 90031-1 . PMC 7133365 . PMID 2846182 .
- ^ Jagger BW, Wise HM, Kash JC, Walters Ka, Wills NM, Xiao YL, Dunfee RL, Schwartzman LM, Ozinsky A, Bell GL, Dalton RM, Lo A, Efstathiou S, Atkins JF, Firth Ae, Taubenberg (Июль 2012 г.). «Перекрывающаяся область кодирования белка в сегменте 3 вируса гриппа A модулирует ответ хозяина» . Наука . 337 (6091): 199–204. Bibcode : 2012sci ... 337..199J . doi : 10.1126/science.12222213 . PMC 3552242 . PMID 22745253 .
- ^ Advani VM, Dinman JD (январь 2016 г.). «Перепрограммирование генетического кода: появляющаяся роль рибосомального фреймэфира в регуляции экспрессии клеточных генов» . Биологии . 38 (1): 21–26. doi : 10.1002/bies.201500131 . PMC 4749135 . PMID 26661048 .
- ^ Крик Ф.Х. (август 1966 г.). «Кодон - пары Антидона: гипотеза колебания». Журнал молекулярной биологии . 19 (2): 548–555. doi : 10.1016/s0022-2836 (66) 80022-0 . PMID 5969078 .
- ^ Jump up to: а беременный Бриерли I (август 1995 г.). «Рибосомальный кадр вирусных РНК» . Журнал общей вирусологии . 76 (Pt 8) (8): 1885–1892. doi : 10.1099/0022-1317-76-8-1885 . PMID 7636469 .
- ^ Jump up to: а беременный в дюймовый Harger JW, Meskauskas A, Dinman JD (сентябрь 2002 г.). «Интегрированная модель» запрограммированного рибосомального фарма -снимания » . Тенденции в биохимических науках . 27 (9): 448–454. doi : 10.1016/s0968-0004 (02) 02149-7 . PMID 12217519 .
- ^ Gurvich Oli, Baranv Pv, Gesteland RF, Atkins JF (июнь 2005 г.). «Уровни экспрессии уровни влияют на рибсомальный фреймэффициент на кодонах тандмананена . Журнал бактераологии 187 (12): 4023–4 Doi : 10.1128/ jb.187.12.4023-4 1151738PMC 15937165PMID
- ^ Калискан Н., Катунин В.И., Белардинелли Р., Песке Ф., Роднина М.В. (июнь 2014 г.). «Запрограммирован -1 рамки с помощью кинетического распределения во время препятствий транслокации» . Клетка . 157 (7): 1619–1631. doi : 10.1016/j.cell.2014.04.041 . PMC 7112342 . PMID 24949973 .
- ^ Артур Л., Павлович-Джуранович С., Смит-Кутму К., Грин Р., Zczesny P, Djuranovic S (июль 2015). «Трансляционный контроль с помощью A-богатых последовательностей A-кодирования лизина» . Наука достижения . 1 (6): E1500154. Bibcode : 2015scia .... 1e0154a . doi : 10.1126/sciadv.1500154 . PMC 4552401 . PMID 26322332 .
- ^ Хансен Т.М., Рейхани С.Н., Одершхед Л.Б., Сёренсен М.А. (апрель 2007 г.). «Корреляция между механической прочностью псевдокнотов РНК мессенджера и рибосомным кадром» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (14): 5830–5835. Bibcode : 2007pnas..104.5830h . doi : 10.1073/pnas.0608668104 . PMC 1838403 . PMID 17389398 .
- ^ Jump up to: а беременный в Комитет по номенклатуру Международного союза биохимии (NC-IUB) (1984). «Номенклатура для не полностью указанных оснований в последовательностях нуклеиновых кислот» . Получено 4 февраля 2008 года .
- ^ Mazauric MH, Licznar P, Prère MF, Canal I, Fayet O (июль 2008 г.). «Апикальная петля-внутренняя петля РНК псевдокноты: новый тип стимулятора -1 трансляционного кадрирования у бактерий» . Журнал биологической химии . 283 (29): 20421–20432. doi : 10.1074/jbc.m802829200 . PMID 18474594 .
- ^ Иванов IP, Андерсон CB, Gesteland RF, Atkins JF (июнь 2004 г.). «Идентификация новой мРНК -антизима +1, снимающая стимулирующие псевдокнот в подмножестве разнообразных беспозвоночных и его очевидное отсутствие у промежуточных видов» . Журнал молекулярной биологии . 339 (3): 495–504. doi : 10.1016/j.jmb.2004.03.082 . PMC 7125782 . PMID 15147837 .
- ^ Баранов П.В., Хендерсон С.М., Андерсон К.Б., Гестеланд Р.Ф., Аткинс Дж. Ф., Говард М.Т. (февраль 2005 г.). «Запрограммированное рибосомное фармаризацию в декодировании генома SARS-COV» . Вирусология . 332 (2): 498–510. doi : 10.1016/j.virol.2004.11.038 . PMC 7111862 . PMID 15680415 .
- ^ Larsen B, Gesteland RF, Atkins JF (август 1997 г.). «Структурное зондирование и мутагенный анализ петли ствола, необходимый для рибосомального фрамеирования Escherichia coli: запрограммированная эффективность 50%» . Журнал молекулярной биологии . 271 (1): 47–60. doi : 10.1006/jmbi.1997.1162 . PMC 7126992 . PMID 9300054 .
Внешние ссылки
[ редактировать ]- FrameShifting,+рибосомаль в Национальной библиотеке медицины Медицинской библиотеки США (Mesh)
- WISE2 - выравнивает белок против последовательности ДНК , позволяющих кадрировать и интроны
- Быстро - сравните последовательность ДНК с базой данных последовательностей белка , позволяя пробелы и кадры
- Путь архивировал 19 июля 2011 года в The Wayback Machine- инструмент, который сравнивает два FrameShift белка ( обратного перевода ) принцип
- Recode2 - база данных перекодированных генов, в том числе те, которые требуют запрограммированного трансляционного съемки кадров.