Сиаловая кислота
 N -ацетилнейраминовая кислота, наиболее распространенная из сиаловых кислот. |
Сиаловые кислоты представляют собой класс альфа- кетокислот сахаров с девятиуглеродным остовом . [ 1 ] Термин «сиаловая кислота» (от греческого σίαλον (сиалон) « слюна ») был впервые введен шведским биохимиком Гуннаром Бликсом в 1952 году. Наиболее распространенным представителем этой группы является N -ацетилнейраминовая кислота (Neu5Ac или NANA), обнаруженная у животных и некоторых прокариоты .
Сиаловые кислоты широко распространены в тканях животных, а родственные формы встречаются в меньшей степени в других организмах, например, в некоторых микроводорослях . [ 2 ] бактерии и археи . [ 3 ] [ 4 ] [ 5 ] [ 6 ] Сиаловые кислоты обычно входят в состав гликопротеинов , гликолипидов или ганглиозидов , где они украшают концы сахарных цепей на поверхности клеток или растворимых белков. [ 7 ] Однако сиаловые кислоты также наблюдались в эмбрионах дрозофилы и других насекомых. [ 8 ] Как правило, растения не содержат и не проявляют сиаловые кислоты. [ 9 ]
У людей мозг имеет самое высокое содержание сиаловых кислот, где эти кислоты играют важную роль в нервной передаче и структуре ганглиозидов в синаптогенезе . [ 7 ] Известно более 50 видов сиаловой кислоты, все из которых можно получить из молекулы нейраминовой кислоты путем замены ее аминогруппы или одной из ее гидроксильных групп. [ 1 ] Обычно аминогруппа содержит либо ацетильную, либо гликолильную группу, но были описаны и другие модификации. Было показано, что эти модификации наряду со связями являются тканеспецифичными и регулируемыми в процессе развития экспрессиями , поэтому некоторые из них обнаруживаются только в определенных типах гликоконъюгатов в конкретных клетках. [ 8 ] Гидроксильные ; заместители могут значительно различаться ацетильная , лактильная , метильная , сульфатная и фосфатная группы. Обнаружены [ 10 ]
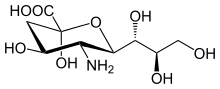
Структура
[ редактировать ]Семейство сиаловых кислот включает множество производных девятиуглеродного сахара нейраминовой кислоты , но эти кислоты редко встречаются в природе в свободном виде. В норме их можно обнаружить в составе олигосахаридных цепей муцинов, гликопротеинов и гликолипидов, занимающих концевые, невосстанавливающие положения сложных углеводов как на внешних, так и на внутренних участках мембраны, где они очень обнажены и выполняют важные функции. [ 7 ]

Нумерация атомов углерода начинается с карбоксилатного углерода и продолжается по всей цепи. Конфигурация, которая помещает карбоксилат в аксиальное положение, представляет собой альфа-аномер.

Альфа-аномер — это форма, которая возникает, когда сиаловая кислота связана с гликанами. Однако в растворе он находится преимущественно (более 90%) в бета-аномерной форме. Был обнаружен бактериальный фермент с мутаротазной активностью сиаловой кислоты, NanM, который способен быстро уравновешивать растворы сиаловой кислоты до положения равновесия покоя, составляющего около 90% бета/10% альфа. [ 11 ]
В отличие от других животных, люди генетически не способны производить вариант сиаловой кислоты N-гликолилнейраминовую кислоту (Neu5Gc). Однако небольшие количества Neu5Gc, обнаруженные в тканях человека, могут быть включены из экзогенных (питательных) источников. [ 12 ]
Биосинтез
[ редактировать ]Сиаловая кислота синтезируется глюкозамин-6-фосфатом и ацетил-КоА через трансферазу , в результате чего образуется N -ацетилглюкозамин-6-P. он превращается в N- ацетилманнозамин-6-P В результате эпимеризации , который реагирует с фосфоенолпируватом с образованием N -ацетилнейрамин-9-P (сиаловой кислоты). Чтобы он стал активным и включился в процесс биосинтеза олигосахаридов в клетке, к нему добавляется монофосфатный нуклеозид, который образуется из цитидинтрифосфата , превращая сиаловую кислоту в цитидинмонофосфат-сиаловую кислоту (CMP-сиаловая кислота). Это соединение синтезируется в ядре животной клетки. [ 13 ] [ 14 ]
В бактериальных системах сиаловые кислоты также могут биосинтезироваться альдолазой . Этот фермент использует, например, производное маннозы в качестве субстрата, встраивая три атома углерода из пирувата в полученную структуру сиаловой кислоты. Эти ферменты могут быть использованы для химиоферментативного синтеза производных сиаловой кислоты. [ 15 ]

Функция
[ редактировать ]Гликопротеины, содержащие сиаловую кислоту ( сиалогликопротеины ), связывают селектин у человека и других организмов. Метастатические раковые клетки часто экспрессируют высокую плотность гликопротеинов, богатых сиаловой кислотой. Эта сверхэкспрессия сиаловой кислоты на поверхностях создает отрицательный заряд на клеточных мембранах. Это создает отталкивание между клетками (клеточное противостояние). [ 16 ] и помогает этим раковым клеткам поздней стадии попасть в кровоток. Недавние эксперименты продемонстрировали присутствие сиаловой кислоты во внеклеточном матриксе, секретируемом раком . [ 17 ]
, богатые сиаловой кислотой, Олигосахариды на гликоконъюгатах (гликолипиды, гликопротеины, протеогликаны), обнаруженные на поверхностных мембранах, помогают удерживать воду на поверхности клеток. [ нужна ссылка ] . Области, богатые сиаловой кислотой, способствуют созданию отрицательного заряда на поверхности клеток. Поскольку вода представляет собой полярную молекулу с частичными положительными зарядами на обоих атомах водорода, она притягивается к клеточным поверхностям и мембранам. Это также способствует поглощению клеточной жидкости.
Остатки сиаловой кислоты присутствуют в муциновых гликопротеинах слизи. [ 18 ]
Сиаловая кислота может «прятать» маннозные антигены на поверхности клеток-хозяев или бактерий от маннозосвязывающего лектина. [ нужна ссылка ] Это предотвращает активацию комплемента .
Сиаловая кислота в форме полисиаловой кислоты представляет собой необычную посттрансляционную модификацию , которая возникает в молекулах адгезии нервных клеток (NCAM). В синапсе сильный отрицательный заряд полисиаловой кислоты предотвращает сшивание клеток NCAM.
Введение эстрогена кастрированным мышам приводит к дозозависимому снижению содержания сиаловой кислоты во влагалище. И наоборот, содержание сиаловой кислоты во влагалище мыши является мерой активности эстрогена. Эталонными веществами являются эстрадиол для подкожного применения и этинилэстрадиол для перорального применения. [ 19 ]
Иммунитет
[ редактировать ]Сиаловые кислоты обнаружены на всех поверхностях клеток позвоночных и некоторых беспозвоночных, а также у некоторых бактерий, взаимодействующих с позвоночными.
Многие вирусы, такие как Ad26 [ 20 ] серотипы аденовирусов ( Adenoviridae ), ротавирусов ( Reoviridae ) и вирусов гриппа ( Orthomyxoviridae ) могут использовать сиалилированные структуры хозяина для связывания с клеткой-мишенью-хозяином. Сиаловые кислоты являются хорошей мишенью для этих вирусов, поскольку они высококонсервативны и в большом количестве присутствуют практически во всех клетках. Неудивительно, что сиаловые кислоты также играют важную роль в некоторых вирусных инфекциях человека. Вирусы гриппа имеют на своей поверхности гликопротеины гемагглютининовой активности (НА), которые связываются с сиаловыми кислотами, обнаруженными на поверхности эритроцитов человека и на клеточных мембранах верхних дыхательных путей. На этом основана гемагглютинация, когда вирусы смешиваются с клетками крови, и проникновение вируса в клетки верхних дыхательных путей. Широко используемые противогриппозные препараты ( осельтамивир и занамивир ) представляют собой аналоги сиаловой кислоты, которые препятствуют высвобождению вновь созданных вирусов из инфицированных клеток путем ингибирования вирусного фермента нейраминидазы . [ 21 ]
Некоторые бактерии также используют сиалилированные структуры хозяина для связывания и распознавания. Например, данные показывают, что свободные сиаловые кислоты могут служить сигналом для некоторых конкретных бактерий, таких как пневмококк . Свободная сиаловая кислота, возможно, может помочь бактерии распознать, что она достигла среды позвоночных, подходящей для ее колонизации. Модификации Sias, такие как N -гликолильная группа в положении 5 или O -ацетильные группы в боковой цепи, могут снизить действие бактериальных сиалидаз. [ 21 ]
Метаболизм
[ редактировать ]Синтез и деградация сиаловой кислоты распределяются в разных компартментах клетки. Синтез начинается в цитозоле, где N -ацетилманнозамин- 6-фосфат и фосфоенолпируват дают начало сиаловой кислоте. Позже фосфат Neu5Ac 9 активируется в ядре остатком цитидинмонофосфата (CMP) через синтазу CMP-Neu5Ac. Хотя связь между сиаловой кислотой и другими соединениями, как правило, представляет собой α-связывание, эта конкретная связь является единственной, которая представляет собой β-связь. CMP-Neu5Ac затем транспортируется в эндоплазматический ретикулум или аппарат Гольджи, где он может быть перенесен на олигосахаридную цепь, становясь новым гликоконъюгатом. Эту связь можно модифицировать путем О- ацетилирования или О- метилирования . Когда гликоконъюгат созревает, он транспортируется на поверхность клетки.
Сиалидаза является одним из наиболее важных ферментов катаболизма сиаловой кислоты. Это может вызвать удаление остатков сиаловой кислоты с поверхности клеток или сывороточных сиалогликоконъюгатов. Обычно у высших животных гликоконъюгаты, склонные к деградации, захватываются путем эндоцитоза. После слияния поздней эндосомы с лизосомой лизосомальные сиалидазы удаляют остатки сиаловой кислоты. Действие этих сиалидаз основано на удалении О -ацетильных групп. Молекулы свободной сиаловой кислоты транспортируются в цитозоль через мембрану лизосомы. Там они могут быть переработаны и снова активированы для образования еще одной зарождающейся молекулы гликоконъюгата в аппарате Гольджи. Сиаловые кислоты также могут расщепляться до ацилманнозамина и пирувата с помощью цитозольного фермента ацилнейраминатиазы.
Некоторые тяжелые заболевания могут зависеть от присутствия или отсутствия некоторых ферментов, связанных с метаболизмом сиаловой кислоты. Сиалидоз и дефицит сиаловой кислоты с мутациями в гене NANS (см. ниже) могут быть примерами этого типа расстройства. [ 22 ]
Развитие мозга
[ редактировать ]Крысиные детеныши, получавшие сиаловую кислоту, продемонстрировали улучшение обучения и памяти во взрослом возрасте. [ 23 ] Взаимосвязь между добавлением сиаловой кислоты в рацион и когнитивной функцией наблюдалась у поросят, которых кормили высокими дозами сиаловой кислоты. [ 24 ]

Болезни
[ редактировать ]Сиаловые кислоты связаны с несколькими различными заболеваниями, наблюдаемыми у людей.
Дефицит сиаловой кислоты с мутациями NANS гена
[ редактировать ]Биаллельные рецессивные мутации в гене синтеза сиаловой кислоты, синтазе N-ацетилнейраминовой кислоты ( NANS ), у людей могут привести к тяжелому заболеванию, проявляющемуся умственной отсталостью и низким ростом, что подчеркивает важность сиаловой кислоты в развитии мозга. [ 25 ] Терапевтическое исследование с краткосрочным пероральным приемом сиаловой кислоты не выявило значительного положительного влияния на биохимические параметры. [ 26 ]
болезнь Саллы
[ редактировать ]Болезнь Салла — чрезвычайно редкое заболевание, которое считается самой легкой формой нарушения накопления свободной сиаловой кислоты. [ 27 ] хотя его детская форма считается агрессивным вариантом, и люди, страдающие от нее, страдают умственной отсталостью. [ 28 ] Это аутосомно-рецессивное заболевание, вызванное мутацией хромосомы 6 . [ 29 ] В основном поражает нервную систему [ 27 ] и это вызвано нарушением лизосомального накопления, которое возникает из-за дефицита специфического переносчика сиаловой кислоты, расположенного на лизосомальной мембране. [ 30 ] В настоящее время не существует лекарства от этого заболевания, и лечение носит поддерживающий характер, направленный на контроль симптомов. [ 27 ]
Атеросклероз
[ редактировать ]Подфракции холестерина ЛПНП, которые участвуют в возникновении атеросклероза, имеют пониженный уровень сиаловой кислоты. [ 31 ] К ним относятся небольшие частицы ЛПНП высокой плотности и электроотрицательные ЛПНП. [ 31 ] Снижение уровня сиаловой кислоты в небольших частицах ЛПНП высокой плотности увеличивает сродство этих частиц к протеогликанам в стенках артерий . [ 31 ]
Грипп
[ редактировать ]Всем штаммам вируса гриппа А для связи с клетками необходима сиаловая кислота. Существуют различные формы сиаловых кислот, которые имеют различное сродство к разновидностям вируса гриппа А. Это разнообразие является важным фактом, определяющим, какие виды могут быть заражены. [ 32 ] Когда определенный вирус гриппа А распознается рецептором сиаловой кислоты, клетка имеет тенденцию эндоцитировать вирус, поэтому клетка становится инфицированной.
Сиаловые кислоты и другие неулозоновые кислоты (NulOs) у прокариот
[ редактировать ]Сиаловые кислоты широко распространены в тканях позвоночных, где они участвуют во многих различных биологических процессах. Первоначально обнаруженные в линии вторичноротых животных, сиаловые кислоты на самом деле можно рассматривать как подмножество более древнего семейства моносахаридов с 9-углеродным остовом, называемых ноулозоновыми кислотами (NulOs), которые совсем недавно были также обнаружены у Eubacteria и Archaea. [ 33 ] Многие патогенные бактерии включают сиаловую кислоту в элементы клеточной поверхности, такие как липополисахарид или капсульные полисахариды, что помогает им уклоняться от врожденного иммунного ответа хозяина. [ 34 ] Недавнее исследование на уровне генома изучило большой набор секвенированных микробных геномов, которые показали, что пути биосинтеза для производства неулозоновых кислот (NulOs) гораздо более широко распространены на филогенетическом древе жизни, чем предполагалось ранее. [ 35 ] Более того, этот вывод подтверждается недавними исследованиями окрашивания лектинов и исследованием молекулярного уровня прокариотических неулозоновых кислот, показавшими, что также многие непатогенные и чисто экологические штаммы продуцируют бактериальные сиаловые кислоты (NulOs). [ 36 ] [ 37 ] Некоторые бактерии ( анаммокс ) производят NulO, которые в дополнение к очень кислой группе альфа-кетокислот также содержат (нейтрализующие) основные группы (свободные амины). [ 38 ] Сопоставимые сиаловые кислоты клеточной поверхности были получены путем химического ремоделирования для управления зарядом клеточной поверхности путем образования свободного амина в положении C5, который нейтрализует отрицательно заряженную карбоксильную группу в положении C1. [ 39 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б Варки, Аджит; Роланд Шауэр (2008). «Сиаловые кислоты» . в Основах гликобиологии . Пресса Колд-Спринг-Харбор. пп. гл. 14. ISBN 9780879697709 .
- ^ Вагстафф, Бен (2018). «Идентификация пути биосинтеза Kdn у гаптофита Prymnesium parvum предполагает широкое распространение биосинтеза сиаловой кислоты среди микроводорослей» . Журнал биологической химии . 293 (42): 16277–16290. дои : 10.1074/jbc.RA118.004921 . ПМК 6200933 . ПМИД 30171074 .
- ^ Аджит, Варки (2017). «Сиаловые кислоты и другие ненулозвуковые кислоты» . Сиаловые кислоты и другие неулосоновые кислоты. Основы гликобиологии . Cold Spring Harbor Laboratory Press. doi : 10.1101/glycobiology.3e.015 (неактивно с 31 января 2024 г.). PMID 28876847 .
{{cite book}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Клейкамп, Хьюго (2020). «Исследование химического разнообразия микробных нонулозвуковых кислот – универсальный подход к крупномасштабным исследованиям» . Химическая наука . 11 (11): 3074–3080. дои : 10.1039/c9sc06406k . ПМЦ 8157484 . ПМИД 34122812 .
- ^ Льюис, Аманда (2009). «Инновации в биосинтезе сиаловой кислоты у хозяина и микробов, выявленные путем филогеномного предсказания структуры неулозвуковой кислоты» . Труды Национальной академии наук . 106 (32): 13552–13557. Бибкод : 2009PNAS..10613552L . дои : 10.1073/pnas.0902431106 . ПМК 2726416 . ПМИД 19666579 .
- ^ Шауэр, Роланд (2018). «Исследование мира сиаловой кислоты» . Adv CarboHydraulic Chem Biochem . Достижения в химии и биохимии углеводов. 75 (75): 1–213. дои : 10.1016/bs.accb.2018.09.001 . ISBN 9780128152027 . ПМК 7112061 . ПМИД 30509400 .
- ^ Перейти обратно: а б с Ван, Б.; Бранд-Миллер, Дж. (2003). «Роль и потенциал сиаловой кислоты в питании человека» . Европейский журнал клинического питания . 57 (11): 1351–1369. дои : 10.1038/sj.ejcn.1601704 . ПМИД 14576748 .
- ^ Перейти обратно: а б Мандал, К. (1990). «Лектины, связывающие сиаловую кислоту». Эксперименты . 46 (5): 433–441. дои : 10.1007/BF01954221 . ПМИД 2189746 . S2CID 27075067 .
- ^ Варки, Аджит; Роланд Шауэр (2008). «Сиаловые кислоты» . в Основах гликобиологии . Пресса Колд-Спринг-Харбор. пп. гл. 14. ISBN 9780879697709 .
- ^ Шауэр Р. (2000). «Достижения и проблемы исследования сиаловых кислот» . Гликокондж. Дж . 17 (7–9): 485–499. дои : 10.1023/А:1011062223612 . ПМЦ 7087979 . ПМИД 11421344 .
- ^ Севери Э., Мюллер А., Поттс-младший, Лич А., Уильямсон Д., Уилсон К.С., Томас Г.Х. (2008). «Мутаротация сиаловой кислоты катализируется белком бета-пропеллера YjhT Escherichia coli» . J Биол Хим . 283 (8): 4841–91. дои : 10.1074/jbc.M707822200 . ПМИД 18063573 .
- ^ Тангворанунтакул, Пэм (14 октября 2003 г.). «Поглощение человеком и включение иммуногенной нечеловеческой пищевой сиаловой кислоты» . ПНАС . 100 (21): 12045–12050. Бибкод : 2003PNAS..10012045T . дои : 10.1073/pnas.2131556100 . ПМК 218710 . ПМИД 14523234 .
- ^ Фулчер, Калифорния, «Химерный путь MetaCyc: суперпуть биосинтеза сиаловой кислоты и CMP-сиаловой кислоты» , «MetaCyc, март 2009 г.»
- ^ Уоррен, Леонард; Фельзенфельд, Герберт (1962). «Биосинтез сиаловых кислот» (PDF) . Журнал биологической химии . 237 (5): 1421. doi : 10.1016/S0021-9258(19)83718-3 .
- ^ Хай Ю; Харшал Чокхавала; Шэншу Хуан и Си Чен (2006). «Трёхферментный химиоферментативный подход к синтезу сиалозидов, содержащих природные и неприродные функциональные группы», в одном горшке . Протоколы природы . 1 (5): 2485–2492. дои : 10.1038/nprot.2006.401 . ПМЦ 2586341 . ПМИД 17406495 .
- ^ Фустер, Марк М.; Эско, Джеффри Д. (2005). «Кисло-сладкое рака: гликаны как новые терапевтические мишени». Обзоры природы Рак . 5 (7): 526–42. дои : 10.1038/nrc1649 . ПМИД 16069816 . S2CID 10330140 .
- ^ Дасгупта, Дебаян; Палли, Дхарма; Шайни, Дипак; Бхат, Рамрей; Гош, Амбариш (2020). «Наномоторы чувствуют локальные физико-химические неоднородности в микроокружении опухоли» . Ангеванде Хеми . 59 (52): 23690–23696. дои : 10.1002/anie.202008681 . ПМЦ 7756332 . ПМИД 32918839 .
- ^ Коэн, Мириам (2013). «Грипп А проникает в слизь хозяина, расщепляя сиаловые кислоты нейраминидазой» . Вирусологический журнал . 10 :321. дои : 10.1186/1743-422X-10-321 . ПМЦ 3842836 . ПМИД 24261589 .
- ^ Юрген Сандов; Эккехард Шайфеле; Майкл Харинг; Гюнтер Нееф; Клаус Прежевовский; Ульрих Стахе (2007), «Гормоны», Энциклопедия промышленной химии Ульмана (7-е изд.), Wiley, стр. 1–81, номер домена : 10.1002/14356007.a13_089 , ISBN. 978-3527306732
- ^ Бейкер, Александр Т.; Манди, Рози М.; Дэвис, Джеймс А.; Ризкалла, Пьер Ж.; Паркер, Алан Л. (сентябрь 2019 г.). «Аденовирус человека типа 26 использует гликаны, несущие сиаловую кислоту, в качестве первичного рецептора входа в клетку» . Достижения науки . 5 (9): eaax3567. Бибкод : 2019SciA....5.3567B . дои : 10.1126/sciadv.aax3567 . ПМК 6726447 . ПМИД 31517055 .
- ^ Перейти обратно: а б Варки А.; Ганье П. (2012). «Многообразная роль сиаловых кислот в иммунитете» . Энн, Нью-Йоркская академия наук . 1253 (1): 16–36. Бибкод : 2012NYASA1253...16V . дои : 10.1111/j.1749-6632.2012.06517.x . ПМК 3357316 . ПМИД 22524423 .
- ^ Трэвинг, К.; Шауэр, Р. (1998). «Структура, функции и метаболизм сиаловых кислот» . Клеточные и молекулярные науки о жизни . 54 (12): 1330–1349. дои : 10.1007/s000180050258 . ПМК 7082800 . ПМИД 9893709 .
- ^ Оливерос Э., Васкес Э., Барранко А., Рамирес М., Груарт А., Дельгадо-Гарсия Х.М., Бак Р., Руэда Р., Мартин М.Дж. (2018). «Прием сиаловой кислоты и сиалилированных олигосахаридов во время лактации улучшает обучение и память у крыс» . Питательные вещества . 10 (10): Е1519. дои : 10.3390/nu10101519 . ПМК 6212975 . ПМИД 30332832 .
- ^ Ван Б. (2012). «Молекулярный механизм, лежащий в основе сиаловой кислоты как важного питательного вещества для развития мозга и познания» . Адв. Нутр . 3 (3): 465С–472С. дои : 10.3945/ан.112.001875 . ПМЦ 3649484 . ПМИД 22585926 .
- ^ ван Карнебек, Клара Д.М.; Бонафе, Луиза; Йен, Сяо-Янь; Тарайло-Граовац, Майя; Бальзано, Сара; Ройе-Бертран, Берил; Ашиков, Ангел; Гаравелли, Ливия; Мама, Изабелла; Туролла, Ликия; Брин, Кэтрин (июль 2016 г.). «NANS-опосредованный синтез сиаловой кислоты необходим для развития мозга и скелета» . Природная генетика . 48 (7): 777–784. дои : 10.1038/ng.3578 . hdl : 2066/174072 . ISSN 1546-1718 . ПМИД 27213289 . S2CID 24953080 .
- ^ Тран С, Туролла Л, Бальхаузен Д, Бурос СК, Тив Т, Галларт-Аяла Х, Иванишевич Дж, Фаузи М, Лефебер Д.Д., Ивановский И, Джанджиоббе С., Караффи С.Г., Гаравелли Л., Суперти-Фурга А (2021). «Судьба перорально принимаемой сиаловой кислоты: первые сведения от пациентов с дефицитом синтазы N-ацетилнейраминовой кислоты и контрольных субъектов» . Представитель Мол Генет Метаб . 28 : 100777. doi : 10.1016/j.ymgmr.2021.100777 . ПМЦ 8251509 . ПМИД 34258226 .
- ^ Перейти обратно: а б с «Болезнь Салла | Информационный центр по генетическим и редким заболеваниям (GARD) – программа NCATS» .
- ^ «Болезнь накопления свободной сиаловой кислоты» . Сирота . Проверено 21 февраля 2019 г.
- ^ Понсо, Г. (2007). «Болезни отложения свободной сиаловой кислоты: болезнь Салла (включая ее тяжелую инфантильную форму) и сиалурия». EMC - Педиатрия (на испанском языке). 42 :1–3. дои : 10.1016/S1245-1789(07)70257-3 .
- ^ «Болезни отложения свободной сиаловой кислоты: болезнь Саллы (включая ее тяжелую инфантильную форму) и сиалурия» (на испанском языке).
- ^ Перейти обратно: а б с Иванова Е.А., Мясоедова В.А., Мельниченко А.А., Гречко А.В., Орехов АН (2017). «Маленькие плотные липопротеины низкой плотности как биомаркер атеросклеротических заболеваний» . Окислительная медицина и клеточное долголетие . 2017 (10): 1273042. doi : 10.1155/2017/1273042 . ПМЦ 5441126 . ПМИД 28572872 .
- ^ Раканиелло, Винсент (5 мая 2009 г.). «Прикрепление вируса гриппа к клеткам: роль различных сиаловых кислот» . Блог вирусологии . Проверено 10 апреля 2019 г.
- ^ Аджит, Варки (2015). «Сиаловые кислоты и другие ненулозвуковые кислоты». Сиаловые кислоты и другие неулозоновые кислоты, Основы гликобиологии (3-е изд.). Глава 15.: Колд-Спринг-Харбор (Нью-Йорк): Лабораторное издательство Колд-Спринг-Харбор. стр. 2015–2017 гг. doi : 10.1101/glycobiology.3e.015 (неактивен 31 января 2024 г.). ПМИД 28876847 .
{{cite book}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) CS1 maint: местоположение ( ссылка ) - ^ Севери Э.; Капот ДВ; Томас Г.Х. (2007). «Утилизация сиаловой кислоты бактериальными патогенами» . Микробиология . 153 (9): 2817–2822. дои : 10.1099/mic.0.2007/009480-0 . ПМИД 17768226 .
- ^ Аманда, Льюис (2009). «Инновации в биосинтезе сиаловой кислоты у хозяина и микробов, выявленные путем филогеномного предсказания структуры неулозвуковой кислоты» . Труды Национальной академии наук . 106 (32): 13552–13557. Бибкод : 2009PNAS..10613552L . дои : 10.1073/pnas.0902431106 . ПМК 2726416 . ПМИД 19666579 .
- ^ Клейкамп, Хьюго (2020). «Решение вопроса о химическом разнообразии микробных нонулозвуковых кислот – универсальный подход к крупномасштабным исследованиям» . Химическая наука . 11 (11): 3074–3080. дои : 10.1039/C9SC06406K . ПМЦ 8157484 . ПМИД 34122812 .
- ^ Болей, Марисса (31 марта 2020 г.). «Украшение дома Анаммокса: сиаловые кислоты и сульфатированные гликозаминогликаны во внеклеточных полимерных веществах гранулированного ила Анаммокса» . Окружающая среда. наук. Технол . 54 (8): 5218–5226. Бибкод : 2020EnST...54.5218B . дои : 10.1021/acs.est.9b07207 . ПМЦ 7181257 . ПМИД 32227885 .
- ^ Пабст, Мартин; Гроуздев Денис С.; Лоусон, Кристофер Э.; Клейкамп, Хьюго, Британская Колумбия; де Рам, Кэрол; Лоуэн, Рожье; Линь, Юэ Мэй; Люкер, Себастьян; ван Лоосдрехт, Марк CM; Лаурени, Микеле (2 августа 2021 г.). «Общий подход к изучению гликозилирования прокариотических белков обнаруживает уникальную модуляцию поверхностного слоя анаммокс-бактерии» . Журнал ISME . 16 (2): 346–357. дои : 10.1038/s41396-021-01073-y . ISSN 1751-7370 . ПМЦ 8776859 . ПМИД 34341504 .
- ^ Ван, Цзе; Ченг, Бо; Ли, Цзе; Чжан, Чжаоюэ; Хун, Вейяо; Чен, Син; Чен, Пэн Р. (2015). «Химическое ремоделирование сиаловых кислот на клеточной поверхности посредством реакции биоортогонального элиминирования, запускаемой палладием» . Angewandte Chemie, международное издание . 54 (18): 5364–5368. дои : 10.1002/anie.201409145 . ISSN 1521-3773 . ПМИД 25765364 .
Внешние ссылки
[ редактировать ]| Базы данных органов управления : Национальные |
|---|