РНК-термометр
РНК -термометр (или РНК-термосенсор ) представляет собой температуре чувствительную к некодирующую молекулу РНК, которая регулирует экспрессию генов . [ 1 ] Его уникальная особенность заключается в том, что для функционирования ему не нужны белки или метаболиты, а он реагирует только на изменения температуры. [ 2 ] РНК-термометры часто регулируют гены, необходимые во время реакции на тепловой или холодовой шок , но они участвуют и в других регуляторных функциях, таких как патогенность и голодание . [ 1 ]
В общем, РНК-термометры работают, изменяя свою вторичную и третичную структуру. [ 3 ] в ответ на колебания температуры. Этот структурный переход может затем обнажить или закрыть важные области РНК, такие как сайт связывания рибосомы , что затем влияет на скорость трансляции близлежащего гена, кодирующего белок .
РНК-термометры, наряду с рибопереключателями , используются в качестве примеров в поддержку гипотезы мира РНК . Эта теория предполагает, что РНК когда-то была единственной нуклеиновой кислотой, присутствующей в клетках, и была заменена нынешней системой ДНК → РНК → белок . [ 4 ]
Примеры РНК-термометров включают FourU , [ 5 ] элемент Hsp90 цис- регуляторный , [ 6 ] элемент РОЗА , [ 7 ] термометр Лиг РНК , [ 8 ] и термометр Hsp17 . [ 9 ]
Открытие
[ редактировать ]О первом термочувствительном элементе РНК было сообщено в 1989 году. [ 10 ] До этого исследования было обнаружено, что мутации выше места начала транскрипции в лямбда (λ) фага cIII мРНК влияют на уровень трансляции белка cIII. [ 11 ] Этот белок участвует в выборе литического или лизогенного жизненного цикла фага λ, при этом высокие концентрации cIII способствуют лизогении. [ 11 ] Дальнейшее исследование этой вышележащей области РНК выявило две альтернативные вторичные структуры ; Экспериментальное исследование показало, что структуры взаимозаменяемы и зависят как от концентрации ионов магния , так и от температуры. [ 10 ] [ 12 ] Сейчас считается, что этот РНК-термометр способствует входу в литический цикл при тепловом стрессе, что позволяет бактериофагу быстро размножаться и покидать клетку-хозяина. [ 1 ]
Термин «РНК-термометр» был придуман только в 1999 году. [ 13 ] когда его применили к элементу РНК rpoH, идентифицированному в Escherichia coli . [ 14 ] Совсем недавно биоинформатические поиски были использованы для обнаружения нескольких новых потенциальных термометров на основе РНК. [ 15 ] Однако традиционные поиски на основе последовательностей неэффективны, поскольку вторичная структура элемента гораздо более консервативна , чем последовательность нуклеиновой кислоты . [ 15 ]
Биологические реакции и организм чувствительны к температуре для функционирования клеток. РНК-термометры являются эффективным способом реагирования на температуру, поскольку они позволяют клеткам отслеживать и ощущать изменения, чтобы поддерживать клетку живой и стабильной. Механизмы, индуцированные ДНК, РНК или белком, избегают небольших изменений, поскольку, чувствуя любые внешние изменения, [ 16 ]
Бактерии используют термометры РНК, чтобы проникать в своих хозяев и выживать в них, прикрепляясь к хозяину и вызывая колебания его температуры. Бактерии могут быстро реагировать на условия теплового и холодового шока, поскольку РНК-термометры контролируют экспрессию генов на уровне трансляции. [ 16 ]
Первый РНК-термометр, обнаруженный в хлоропласте Chlamydomonas reinhardtii , обнаружен в 5'-UTR мРНК psaA. Его функция была другой, особенно потому, что он считался отсутствующим. Он имеет вторичную структуру типа шпильки, которая защищает последовательность Шайна-Дальгарно при низкой температуре, но как только происходит изменение температуры, он плавится и активирует выработку белка. [ 2 ] Исследование C. Reinhardtii по РНК-термометру — это путь к наблюдению за хлоропластами фотосинтезирующих организмов с целью регуляции генов и к тому, как его можно будет использовать в сельском хозяйстве в какой-то момент в будущем, поскольку он помогает растениям привыкнуть к внешней температуре. [ 2 ]
Распределение
[ редактировать ]Большинство известных РНК-термометров расположены в 5'-нетранслируемой области (UTR) информационной РНК, кодирующей белки теплового шока , хотя предполагается, что этот факт может быть частично обусловлен предвзятостью выборки и присущими трудностями обнаружения коротких, неконсервативных последовательностей РНК. в геномных данных . [ 17 ] [ 18 ]
Хотя потенциальный термометр РНК обнаружен преимущественно у прокариот , он был обнаружен и у млекопитающих, включая человека . [ 19 ] Кандидатная термосенсорная РНК теплового шока РНК-1 (HSR1) активирует фактор транскрипции теплового шока 1 (HSF1) и индуцирует защитные белки, когда температура клеток превышает 37 ° C ( температура тела ), тем самым предотвращая перегрев клеток. [ 19 ]
Первый РНК-термометр, обнаруженный в хлоропласте Chlamydomonas reinhardtii, обнаружен в 5'-UTR мРНК psaA. Его функция была другой, особенно потому, что он считался отсутствующим. Он имеет вторичную структуру типа шпильки, которая защищает последовательность Шайна-Дальгарно при низкой температуре, но как только происходит изменение температуры, он плавится и активирует выработку белка. [ 2 ] Исследование C. Reinhardtii по РНК-термометру — это первый шаг к наблюдению за хлоропластами фотосинтезирующих организмов с целью регуляции генов и к тому, как его можно будет использовать в сельском хозяйстве в какой-то момент в будущем, поскольку он помогает растениям привыкнуть к внешней температуре. [ 2 ]
Элементы ROSE представляют собой класс бактериальных РНК-термометров, который регулирует активацию генов, содержащих небольшие белки теплового шока. Он тает на умеренном уровне параллельно с повышением температуры окружающей среды. Как только он полностью плавится при высокой температуре ~ 42 ° C, он приступает к высвобождению Шайна-Дальгарно и стартового кодона AUG. РНК-термометры также можно найти у некоторых растительных симбиотов или патогенов. Симбиоты и патогены используют РНК-термометры для регулирования экспрессии генов растения. [ 3 ] Хорошо изученными бактериями-симбиотами является семейство Rhizobiaceae. У большинства видов ризобий были видны элементы ROSE (цис-действующие), контролирующие гены теплового шока. [ 3 ]
Структура
[ редактировать ]
РНК-термометры структурно просты и могут быть изготовлены из коротких последовательностей РНК; наименьший из них состоит всего из 44 нуклеотидов и обнаружен в мРНК белка теплового шока hsp17 у Synechocystis видов PCC 6803 . [ 9 ] Обычно эти элементы РНК имеют длину от 60 до 110 нуклеотидов. [ 21 ] и они обычно содержат шпильку с небольшим количеством несовпадающих пар оснований, которые снижают стабильность структуры, тем самым обеспечивая более легкое развертывание в ответ на повышение температуры. [ 17 ]
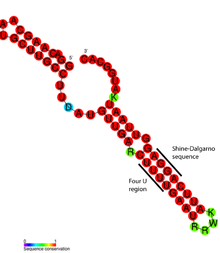
Детальный структурный анализ РНК-термометра ROSE показал, что несовпадающие основания на самом деле участвуют в нестандартном спаривании оснований, которое сохраняет спиральную структуру РНК (см. Рисунок). Необычные пары оснований состоят из пар GG, UU и UC-U. Поскольку эти неканонические пары оснований относительно нестабильны, повышение температуры вызывает локальное плавление структуры РНК в этой области, обнажая последовательность Шайна-Дальгарно. [ 20 ]
Некоторые РНК-термометры значительно сложнее, чем одна шпилька, как в случае с областью мРНК CspA , которая, как полагают, содержит псевдоузел , а также несколько шпилек. [ 22 ] [ 23 ]
Синтетические РНК-термометры имеют простую структуру с одной шпилькой. [ 24 ] Однако вторичная структура таких коротких РНК-термометров может быть чувствительной к мутациям, поскольку изменение одного основания может сделать шпильку неактивной in vivo . [ 25 ]
Механизм
[ редактировать ]
РНК-термометры находятся в 5'- UTR матричной РНК, выше гена, кодирующего белок. [ 1 ] Здесь они способны закупорить сайт связывания рибосомы (RBS) и предотвратить трансляцию мРНК в белок. [ 17 ] При повышении температуры шпилечная структура может «расплавиться» и обнажить последовательность RBS или Шайна-Дальгарно , чтобы обеспечить связывание малой рибосомальной субъединицы ( 30S ), которая затем собирает другой механизм трансляции. [ 1 ] Старт -кодон обычно находится на 8 нуклеотидов ниже последовательности Шайна-Дальгарно. [ 17 ] сигнализирует о начале кодирования белка геном, который затем транслируется в пептидный продукт рибосомой . В дополнение к этому цис -действующему механизму единственный пример транс -действующего РНК-термометра был обнаружен в мРНК RpoS , где, как полагают, она участвует в реакции голодания. [ 1 ]
Конкретным примером мотива термометра РНК является термометр FourU, обнаруженный у Salmonella enterica . [ 5 ] При воздействии температур выше 45 ° C стебель-петля , которая образует пару оснований напротив последовательности Шайна-Дальгарно, становится неспаренной и позволяет мРНК проникнуть в рибосому для осуществления трансляции. [ 25 ] мг 2+ Также было показано, что концентрация ионов влияет на стабильность FourU. [ 26 ] Наиболее хорошо изученный РНК-термометр обнаружен в гене rpoH у Escherichia coli . [ 27 ] Этот термосенсор активирует белки теплового шока при высоких температурах посредством σ 32 , специализированный сигма-фактор теплового шока . [ 13 ]
Хотя РНК-термометры обычно связаны с экспрессией белков, вызванной нагреванием, они также могут регулировать белки холодового шока. [ 22 ] экспрессия двух белков массой 7 кДа регулируется с помощью РНК-термометра. Например, у термофильной бактерии Thermus thermophilus [ 28 ] аналогичный механизм был идентифицирован у Enterobacteriales . [ 23 ]
РНК-термометры, чувствительные к температуре 37 °C, могут использоваться патогенами для активации генов, специфичных для инфекции. [ 17 ] Например, активация prfA с , кодирующего ключевой регулятор транскрипции вирулентности генов Listeria monocytogenes , была продемонстрирована путем слияния 5'-ДНК prfA геном зеленого флуоресцентного белка ; затем слитый ген транскрибировали с промотора Т7 в E. coli , и флуоресценцию наблюдали при 37 ° C, но не при 30 ° C. [ 29 ]
Последствия для гипотезы мира РНК
[ редактировать ]Гипотеза мира РНК утверждает, что когда-то РНК была одновременно носителем наследственной информации и ферментативно активной , причем различные последовательности действовали как биокатализаторы, регуляторы и сенсоры. [ 30 ] Затем гипотеза предполагает, что современная жизнь на основе ДНК, РНК и белков эволюционировала, и отбор заменил большую часть ролей РНК другими биомолекулами . [ 4 ]
Считается, что РНК-термометры и рибопереключатели являются эволюционно древними из-за их широкомасштабного распространения в отдаленно родственных организмах. [ 31 ] Было высказано предположение, что в мире РНК термосенсоры РНК должны были отвечать за температурно-зависимую регуляцию других молекул РНК. [ 4 ] [ 32 ] РНК-термометры в современных организмах могут представлять собой молекулярные окаменелости , что может указывать на ранее более широкое значение в мире РНК. [ 4 ]
Другие примеры
[ редактировать ]- Цис-регуляторный элемент Hsp90 регулирует hsp90 у дрозофилы , увеличивая скорость трансляции белка теплового шока при высоких температурах. [ 6 ]
- ibpAB E. Предполагается , что оперон coli будет содержать два взаимодействующих РНК-термометра: элемент ROSE и термометр IbpB . [ 33 ]
- ROSE 1 и ROSE AT2 обнаружены у Hyphomicrobiales Bradyrhizobium japonicum и Agrobacterium tumefaciens соответственно. Они существуют в 5'-UTR мРНК HspA и подавляют трансляцию белка теплового шока при физиологических температурах. [ 7 ] [ 34 ]
- Цианобактериальные РНК-термометры
- Межгенный РНК-термометр
- РНК-термометры Neisseria
- Lig РНК термометр
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г Нарберхаус Ф, Вальдмингхаус Т, Чоудхури С (январь 2006 г.). «РНК-термометры». Обзоры микробиологии FEMS . 30 (1): 3–16. дои : 10.1111/j.1574-6976.2005.004.x . ПМИД 16438677 .
- ^ Перейти обратно: а б с д и Раза А., Сиддик К.Х., Ху З. (февраль 2024 г.). «Контроль генов хлоропластов: раскрытие механизмов термометра РНК в фотосинтетических системах». Тенденции в науке о растениях . doi : 10.1016/j.tplants.2024.01.005 . ПМИД 38311501 .
- ^ Перейти обратно: а б с Томас С.Е., Бальцерович М., Чанг БАЙ (17 августа 2022 г.). «Терморегуляция, опосредованная структурой РНК: чему мы можем научиться у растений?» . Границы в науке о растениях . 13 : 938570. doi : 10.3389/fpls.2022.938570 . ПМЦ 9450479 . ПМИД 36092413 .
- ^ Перейти обратно: а б с д Аткинс Дж. Ф., Гестеланд РФ, Чех Т (2006). Мир РНК: природа современной РНК предполагает пребиотический мир РНК . Плейнвью, Нью-Йорк: Лабораторное издательство Колд-Спринг-Харбор. ISBN 978-0-87969-739-6 .
- ^ Перейти обратно: а б Вальдмингхаус Т., Гейдрих Н., Брантл С., Нарберхаус Ф (июль 2007 г.). «FourU: новый тип РНК-термометра для сальмонелл» . Молекулярная микробиология . 65 (2): 413–424. дои : 10.1111/j.1365-2958.2007.05794.x . ПМИД 17630972 .
- ^ Перейти обратно: а б Ахмед Р., Дункан Р.Ф. (ноябрь 2004 г.). «Регуляция трансляции мРНК Hsp90. Элементы AUG-проксимальной 5'-нетранслируемой области, необходимые для преимущественной трансляции теплового шока» . Журнал биологической химии . 279 (48): 49919–49930. дои : 10.1074/jbc.M404681200 . ПМИД 15347681 .
- ^ Перейти обратно: а б Нокер А., Хаушерр Т., Балсигер С., Крстулович Н.П., Хеннеке Х., Нарберхаус Ф. (декабрь 2001 г.). «Термосенсор на основе мРНК контролирует экспрессию ризобиальных генов теплового шока» . Исследования нуклеиновых кислот . 29 (23): 4800–4807. дои : 10.1093/нар/29.23.4800 . ПМК 96696 . ПМИД 11726689 .
- ^ Мацунага Дж., Шлакс П.Дж., Хааке Д.А. (ноябрь 2013 г.). «Роль последовательностей цис-действующей РНК в зависимой от температуры экспрессии мультиадгезивных белков lig у Leptospira interrogans» . Журнал бактериологии . 195 (22): 5092–5101. дои : 10.1128/jb.00663-13 . ПМЦ 3811586 . ПМИД 24013626 .
- ^ Перейти обратно: а б Кортманн Дж, Шодрок С, Ринненталь Дж, Швальбе Х, Нарберхаус Ф (апрель 2011 г.). «Перевод по требованию простым термосенсором на основе РНК» . Исследования нуклеиновых кислот . 39 (7): 2855–2868. дои : 10.1093/нар/gkq1252 . ПМК 3074152 . ПМИД 21131278 .
- ^ Перейти обратно: а б Алтувиа С., Корницер Д., Тефф Д., Оппенгейм А.Б. (ноябрь 1989 г.). «Альтернативные структуры мРНК гена cIII бактериофага лямбда определяют скорость инициации его трансляции». Журнал молекулярной биологии . 210 (2): 265–280. дои : 10.1016/0022-2836(89)90329-X . ПМИД 2532257 .
- ^ Перейти обратно: а б Алтувия С, Оппенгейм, AB (июль 1986 г.). «Регуляторные сигналы трансляции в кодирующей области гена бактериофага лямбда cIII» . Журнал бактериологии . 167 (1): 415–419. дои : 10.1128/jb.167.1.415-419.1986 . ПМК 212897 . ПМИД 2941413 .
- ^ Алтувиа С., Корнитцер Д., Коби С., Оппенгейм А.Б. (апрель 1991 г.). «Функциональные и структурные элементы мРНК гена cIII бактериофага лямбда». Журнал молекулярной биологии . 218 (4): 723–733. дои : 10.1016/0022-2836(91)90261-4 . ПМИД 1827163 .
- ^ Перейти обратно: а б Шторц Г. (март 1999 г.). «РНК-термометр» . Гены и развитие . 13 (6): 633–636. дои : 10.1101/gad.13.6.633 . ПМИД 10090718 .
- ^ Морита М.Т., Танака Ю., Кодама Т.С., Кёгоку Ю., Янаги Х., Юра Т. (март 1999 г.). «Трансляционная индукция фактора транскрипции теплового шока сигма32: свидетельства наличия встроенного термосенсора РНК» . Гены и развитие . 13 (6): 655–665. дои : 10.1101/gad.13.6.655 . ПМК 316556 . ПМИД 10090722 .
- ^ Перейти обратно: а б Вальдмингхаус Т., Гаубиг Л.К., Нарберхаус Ф (ноябрь 2007 г.). «Пологеномное биоинформатическое предсказание и экспериментальная оценка потенциальных термометров РНК». Молекулярная генетика и геномика . 278 (5): 555–564. дои : 10.1007/s00438-007-0272-7 . ПМИД 17647020 . S2CID 24747327 .
- ^ Перейти обратно: а б Шарма П., Мондал К., Кумар С., Таманг С., Наджар И.Н., Дас С. и др. (октябрь 2022 г.). «РНК-термометры у бактерий: роль в терморегуляции». Биохимия и биофизика Acta (BBA) - Механизмы регуляции генов . 1865 (7):194871 doi : 10.1016/j.bbagrm.2022.194871 . ПМИД 36041664 .
- ^ Перейти обратно: а б с д и Нарберхаус Ф (2010). «Трансляционный контроль генов бактериального теплового шока и вирулентности с помощью чувствительных к температуре мРНК» . Биология РНК . 7 (1): 84–89. дои : 10.4161/rna.7.1.10501 . ПМИД 20009504 .
- ^ Йоханссон Дж (2009). «РНК-термосенсоры в бактериальных патогенах». Бактериальное зондирование и передача сигналов . Вклад в микробиологию. Том. 16. Базель: С. Каргер АГ. стр. 150–160. дои : 10.1159/000219378 . ISBN 978-3-8055-9132-4 . ПМИД 19494584 .
- ^ Перейти обратно: а б Шамовский И., Иванников М., Кандель Е.С., Гершон Д., Нудлер Э. (март 2006 г.). «РНК-опосредованный ответ на тепловой шок в клетках млекопитающих». Природа . 440 (7083): 556–560. Бибкод : 2006Natur.440..556S . дои : 10.1038/nature04518 . ПМИД 16554823 . S2CID 4311262 .
- ^ Перейти обратно: а б Чоудхури С., Марис С., Аллен Ф.Х., Нарберхаус Ф. (июнь 2006 г.). «Молекулярные основы измерения температуры РНК-термометром» . Журнал ЭМБО . 25 (11): 2487–2497. дои : 10.1038/sj.emboj.7601128 . ПМК 1478195 . ПМИД 16710302 .
- ^ Вальдмингхаус Т., Фиппингер А., Альфсманн Дж., Нарберхаус Ф. (декабрь 2005 г.). «РНК-термометры распространены у альфа- и гамма-протеобактерий». Биологическая химия . 386 (12): 1279–1286. дои : 10.1515/BC.2005.145 . ПМИД 16336122 . S2CID 84557068 .
- ^ Перейти обратно: а б Breaker RR (январь 2010 г.). «РНК отключается на холоде» . Молекулярная клетка . 37 (1): 1–2. doi : 10.1016/j.molcel.2009.12.032 . ПМЦ 5315359 . ПМИД 20129048 .
- ^ Перейти обратно: а б Джулиодори А.М., Ди Пьетро Ф., Марци С., Маскида Б., Вагнер Р., Ромби П. и др. (январь 2010 г.). «МРНК cspA представляет собой термосенсор, который модулирует трансляцию белка холодового шока CspA» . Молекулярная клетка . 37 (1): 21–33. doi : 10.1016/j.molcel.2009.11.033 . ПМИД 20129052 .
- ^ Нойперт Дж., Керхер Д., Бок Р. (ноябрь 2008 г.). «Разработка простых синтетических РНК-термометров для контролируемой температуры экспрессии генов в Escherichia coli» . Исследования нуклеиновых кислот . 36 (19): е124. дои : 10.1093/нар/gkn545 . ПМК 2577334 . ПМИД 18753148 .
- ^ Перейти обратно: а б Николова Е.Н., Аль-Хашими Х.М. (сентябрь 2010 г.). «Термодинамика плавления РНК по одной паре оснований» . РНК . 16 (9): 1687–1691. дои : 10.1261/rna.2235010 . ПМЦ 2924531 . ПМИД 20660079 .
- ^ Ринненталь Дж., Клинкерт Б., Нарберхаус Ф., Швальбе Х. (октябрь 2011 г.). «Модуляция стабильности РНК-термометра Salmonella fourU-типа» . Исследования нуклеиновых кислот . 39 (18): 8258–8270. дои : 10.1093/nar/gkr314 . ПМК 3185406 . ПМИД 21727085 .
- ^ Шах П., Гилкрист, Массачусетс (июль 2010 г.). Спирин А.С. (ред.). «Уникальны ли термочувствительные свойства РНК-термометров?» . ПЛОС ОДИН . 5 (7): е11308. Бибкод : 2010PLoSO...511308S . дои : 10.1371/journal.pone.0011308 . ПМЦ 2896394 . ПМИД 20625392 .
- ^ Мега Р., Мандзоку М., Синкай А., Накагава Н., Курамицу С., Масуи Р. (август 2010 г.). «Очень быстрая индукция белка холодового шока за счет понижения температуры у Thermus thermophilus». Связь с биохимическими и биофизическими исследованиями . 399 (3): 336–340. дои : 10.1016/j.bbrc.2010.07.065 . ПМИД 20655297 .
- ^ Йоханссон Дж., Мандин П., Ренцони А., Кьяруттини С., Спрингер М., Коссарт П. (сентябрь 2002 г.). «Термосенсор РНК контролирует экспрессию генов вирулентности у Listeria monocytogenes» . Клетка . 110 (5): 551–561. дои : 10.1016/S0092-8674(02)00905-4 . ПМИД 12230973 .
- ^ Гилберт В. (февраль 1986 г.). «Мир РНК» . Природа . 319 (6055): 618. Бибкод : 1986Natur.319..618G . дои : 10.1038/319618a0 .
- ^ Серганов А, Патель DJ (октябрь 2007 г.). «Рибозимы, рибопереключатели и не только: регуляция экспрессии генов без белков» . Обзоры природы. Генетика . 8 (10): 776–790. дои : 10.1038/nrg2172 . ПМЦ 4689321 . ПМИД 17846637 .
- ^ Бокобза С.Е., Ахарони А. (октябрь 2008 г.). «Включение света на растительных рибопереключателях». Тенденции в науке о растениях . 13 (10): 526–533. doi : 10.1016/j.tplants.2008.07.004 . ПМИД 18778966 .
- ^ Гаубиг Л.К., Вальдмингхаус Т, Нарберхаус Ф (январь 2011 г.). «Множественные уровни контроля управляют экспрессией оперона теплового шока ibpAB Escherichia coli» . Микробиология . 157 (Часть 1): 66–76. дои : 10.1099/mic.0.043802-0 . ПМИД 20864473 .
- ^ Балсигер С., Рагаз С., Барон С., Нарберхаус Ф. (октябрь 2004 г.). «Репликон-специфическая регуляция малых генов теплового шока у Agrobacterium tumefaciens» . Журнал бактериологии . 186 (20): 6824–6829. дои : 10.1128/JB.186.20.6824-6829.2004 . ПМК 522190 . ПМИД 15466035 .