Первичная моторная кора
| Первичная моторная кора | |
|---|---|
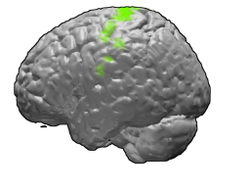 Четвертая зона Бродмана человеческого мозга. | |
 Первичная моторная кора показана зеленым цветом. | |
| Подробности | |
| Часть | Прецентральная извилина |
| Артерия | Передняя мозговая Средний мозговой |
| Идентификаторы | |
| латинский | первая моторная кора |
| Нейроимена | 1910 |
| НейроЛекс ID | nlx_143555 |
| ФМА | 224854 |
| Анатомические термины нейроанатомии | |

Первичная моторная кора ( поле Бродмана 4 ) — это область мозга , которая у человека расположена в дорсальной части лобной доли . Это основная область , двигательной системы которая работает совместно с другими двигательными областями, включая премоторную кору , дополнительную двигательную область , заднюю теменную кору и несколько подкорковых областей мозга, для планирования и выполнения произвольных движений. Первичная моторная кора анатомически определяется как область коры, содержащая крупные нейроны, известные как клетки Бетца , которые, наряду с другими кортикальными нейронами, посылают длинные аксоны вниз по спинному мозгу для синапсов в цепи промежуточных нейронов спинного мозга, а также непосредственно в альфа-мотонейроны спинного мозга, которые соединяются с мышцами.
В первичной моторной коре двигательные представительства упорядоченно расположены (перевернутым образом) от пальца ноги (вверху полушария головного мозга) до рта (внизу) вдоль складки коры, называемой центральной бороздой . Однако некоторые части тела могут контролироваться частично перекрывающимися областями коры. Каждое полушарие головного мозга первичной моторной коры содержит двигательное представительство только противоположной (контралатеральной) стороны тела. Количество первичной моторной коры, отведенной той или иной части тела, пропорционально не абсолютному размеру поверхности тела, а относительной плотности кожных моторных рецепторов на этой части тела. Плотность кожных двигательных рецепторов на той или иной части тела обычно указывает на необходимую степень точности движений, требуемую на этой части тела. По этой причине человеческие руки и лицо имеют гораздо большее изображение, чем ноги.
Информацию об открытии первичной моторной коры и ее связи с другими областями моторной коры см. в основной статье о моторной коре .
Структура
[ редактировать ]Первичная моторная кора человека расположена на передней стенке центральной борозды. Он также простирается вперед из борозды частично на предцентральную извилину. Спереди первичная моторная кора граничит с набором областей, которые лежат на прецентральной извилине и обычно считаются составляющими латеральную премоторную кору. Сзади первичная моторная кора граничит с первичной соматосенсорной корой, лежащей на задней стенке центральной борозды. Вентрально первичная моторная кора граничит с островковой корой в латеральной борозде. Первичная моторная кора простирается дорсально до верхней части полушария, а затем переходит на медиальную стенку полушария.
Расположение первичной моторной коры наиболее очевидно при гистологическом исследовании из-за присутствия характерных клеток Бетца . Слой V первичной моторной коры содержит гигантские (70–100 мкм ) пирамидные нейроны — клетки Бетца. Эти нейроны посылают длинные аксоны к контрлатеральным двигательным ядрам черепных нервов и к нижним мотонейронам передних рогов спинного мозга . Эти аксоны составляют часть кортикоспинального тракта . Клетки Бетца составляют лишь небольшой процент кортикоспинального тракта. По некоторым оценкам, они составляют около 10% нейронов первичной моторной коры, проецирующихся в спинной мозг. [1] или около 2-3% от общей кортикальной проекции спинного мозга. [2] Хотя клетки Бетца не составляют всю двигательную активность коры, они, тем не менее, служат четким маркером первичной моторной коры. Эту область коры, характеризующуюся наличием клеток Бетца, Бродман назвал областью 4.
Клеточные компоненты
[ редактировать ]Было показано, что только первичная моторная кора содержит до 116 различных типов клеток, дифференцированных по своей морфологии, электрофизиологическим свойствам (включая характер импульсов) и профилю экспрессии генов (например, по типу высвобождаемого нейромедиатора (ГАМК, глутамат и т. д.) . [3]
Путь
[ редактировать ]По мере того, как первичные двигательные аксоны проходят через белое вещество головного мозга , они сближаются и образуют часть задней ножки внутренней капсулы .
Они продолжаются вниз в ствол мозга , где некоторые из них, перейдя на контрлатеральную сторону, распределяются по двигательным ядрам черепных нервов . ( Примечание : несколько двигательных волокон образуют синапсы с нижними мотонейронами на той же стороне ствола мозга ).
После перехода на контрлатеральную сторону в продолговатом мозге ( пирамидный перекрест ) аксоны перемещаются вниз по спинному мозгу по латеральному кортикоспинальному пути .
Волокна, которые не перекрещиваются в стволе мозга, проходят вниз по отдельному вентральному кортикоспинальному тракту , и большинство из них переходят на контрлатеральную сторону спинного мозга , незадолго до достижения нижних мотонейронов . Помимо основного кортикоспинального пути, моторная кора проецируется на другие области коры и подкорки, включая полосатое тело, гипоталамус, средний и задний мозг, а также таламус, базальные ганглии, средний и продолговатый мозг. [4]
Кортикомоторные нейроны
[ редактировать ]Кортикомоторнейроны — это нейроны первичной коры головного мозга, которые проецируются непосредственно на мотонейроны вентральных рогов спинного мозга. [5] [6] Аксоны кортикомоторных нейронов заканчиваются на спинальных мотонейронах многих мышц, а также на спинальных интернейронах. [5] [6] Они уникальны для приматов, и было высказано предположение, что их функция заключается в адаптивном контроле над дистальными конечностями (например, руками), включая относительно независимый контроль над отдельными пальцами. [6] Кортикомоторнейроны до сих пор были обнаружены только в первичной моторной коре, а не во вторичных моторных областях. [6]
Кровоснабжение
[ редактировать ]Ветви средней мозговой артерии обеспечивают большую часть артериального кровоснабжения первичной двигательной коры .
Медиальная часть (области ног) кровоснабжается ветвями передней мозговой артерии .
Функция
[ редактировать ]Гомункул
[ редактировать ]В первичной моторной коре имеется широкое представительство различных частей тела в структуре, называемой моторным гомункулом (лат. « маленький человек» ). [7] Область голени расположена близко к средней линии, во внутренних отделах двигательной области сходится в медиальную продольную щель . Латеральная выпуклая сторона первичной моторной коры расположена сверху вниз в областях, соответствующих ягодицам, туловищу, плечу, локтю, запястью, пальцам, большому пальцу, векам, губам и челюсти. Двигательная зона рук и кистей является самой большой и занимает часть прецентральной извилины между ногой и областью лица.
Эти области не пропорциональны своим размерам на теле: губы, части лица и руки представлены особенно большими областями из-за сравнительного обогащения и плотности моторных рецепторов в этих областях. После ампутации или паралича двигательные области могут смещаться, приспосабливаясь к новым частям тела.
Нейронный сигнал от таламуса
[ редактировать ]Первичная моторная кора получает сигналы таламуса от разных ядер таламуса. Среди прочего:
- Вентрально-латеральное ядро афферентов мозжечка.
- Вентральное переднее ядро афферентов базальных ганглиев
Альтернативные карты
[ редактировать ]
Сообщалось по крайней мере о двух модификациях классического соматотопического порядка частей тела в первичной моторной коре приматов.
Во-первых, изображение руки может быть организовано по центральному и объемному принципу. В коре головного мозга обезьяны пальцы руки представлены в центральной области на заднем крае первичной моторной коры. Эта основная область окружена с трех сторон (с дорсальной, передней и вентральной сторон) изображениями более проксимальных частей руки, включая локоть и плечо. [8] [9] У людей изображение пальцев окружено изображением запястья сверху, спереди и снизу. [10]
Вторая модификация классической соматотопической упорядоченности частей тела — двойное представление пальцев и запястья, изучаемое преимущественно в моторной коре человека. Одно представительство находится в задней области, называемой областью 4p, а другое — в передней области, называемой областью 4a. Задняя область может активироваться вниманием без какой-либо сенсорной обратной связи и, как предполагается, важна для начала движений, тогда как передняя область зависит от сенсорной обратной связи. [11] Его также можно активировать воображаемыми движениями пальцев. [12] и слушать речь, не делая реальных движений. Было высказано предположение, что эта передняя область представительства важна для выполнения движений, включающих сложные сенсомоторные взаимодействия. [13] Возможно, что область 4а у человека соответствует некоторым частям каудальной премоторной коры, как описано в коре головного мозга обезьяны.
В 2009 году сообщалось, что существует две эволюционно различные области: более старая на внешней поверхности и новая, обнаруженная в расщелине. Старший соединяется со спинальными мотонейронами через интернейроны спинного мозга. Более новый, обнаруженный только у обезьян и человекообразных обезьян, соединяется непосредственно со спинальными мотонейронами. [14] Прямые связи формируются после рождения, доминируют над непрямыми связями и более гибки в схемах, которые они могут развивать, что позволяет постнатальному обучению сложным навыкам мелкой моторики. «Поэтому появление «новой» области M1 в ходе эволюции линии приматов, вероятно, имело важное значение для повышения ловкости человеческой руки». [15]
Распространенные заблуждения
[ редактировать ]Определенные заблуждения о первичной моторной коре распространены во вторичных обзорах, учебниках и популярных материалах. Здесь перечислены три наиболее распространенных заблуждения.
Разделенная карта тела
[ редактировать ]Одно из наиболее распространенных заблуждений относительно первичной моторной коры состоит в том, что карта тела четко разделена. Однако это не карта отдельных мышц или даже отдельных частей тела. Карта содержит значительные совпадения. Это перекрытие увеличивается в более передних областях первичной моторной коры. Одной из главных целей в истории исследований моторной коры было определение того, насколько различные части тела перекрываются или разделяются в моторной коре. Исследователи, занимавшиеся этой проблемой, обнаружили, что карта кисти, руки и плеча сильно перекрывается. [7] [9] [10] [16] [17] [18] [19] [20] Исследования, которые отображают точную функциональную связь корковых нейронов с мышцами, показывают, что даже один нейрон первичной моторной коры может влиять на активность многих мышц, связанных со многими суставами. [16] В экспериментах на кошках и обезьянах, когда животные учатся сложным, скоординированным движениям, карты первичной моторной коры становятся все более перекрывающимися, очевидно, учатся интегрировать контроль над многими мышцами. [21] [22] У обезьян, когда электрическая стимуляция применяется к моторной коре головного мозга в поведенческом временном масштабе, она вызывает сложные, высокоинтегрированные движения, такие как протягивание руки, предназначенной для хватания, или поднесение руки ко рту и открытие рта. [23] [24] Данные этого типа позволяют предположить, что первичная моторная кора, хотя и содержит приблизительную карту тела, может участвовать в полноценной интеграции мышц, а не в разделении контроля над отдельными группами мышц. Было высказано предположение, что более глубоким принципом организации может быть карта статистических корреляций в поведенческом репертуаре, а не карта частей тела. [24] [25] Поскольку репертуар движений частично распадается на действия отдельных частей тела, карта содержит грубое и перекрывающееся расположение тел.
М1 и первичная моторная кора
[ редактировать ]Термин «М1» и термин «первичная моторная кора» часто используются как синонимы. Однако они происходят из разных исторических традиций и относятся к разным отделам коры головного мозга. Некоторые ученые предположили, что моторную кору можно разделить на первичную моторную полосу, расположенную ближе кзади, и латеральную премоторную полосу, которая расположена ближе кпереди. Среди первых исследователей, которые первоначально предложили эту точку зрения, были Кэмпбелл, [26] Фогт и Фогт, [27] Ферстер, [28] и Фултон. [29] Другие предположили, что моторную кору нельзя разделить таким образом. Вместо этого, на этом втором взгляде, так называемые первичные моторные и латеральные премоторные полосы вместе составляют единую кортикальную область, называемую M1. Вторая двигательная область на медиальной стенке полушария была названа М2 или дополнительной двигательной областью . Среди сторонников этой точки зрения были Пенфилд. [7] и Вулси. [30] Сегодня общепринято различие между первичной моторной корой и латеральной премоторной корой. Однако термин M1 иногда ошибочно используется для обозначения первичной моторной коры. Строго говоря, M1 относится к единой карте, которая, по мнению некоторых предыдущих исследователей, охватывала как первичную моторную, так и латеральную премоторную кору.
Клетки Бетца как последний общий путь
[ редактировать ]Клетки Бетца , или гигантские пирамидные клетки первичной моторной коры, иногда ошибочно принимают за единственный или основной выходной сигнал из коры в спинной мозг. Эта ошибка старая, она восходит, по крайней мере, к Кэмпбеллу в 1905 году. [26] Тем не менее, клетки Бетца составляют лишь около 2-3% нейронов, которые передаются из коры в спинной мозг. [2] и только около 10% нейронов проецируются именно из первичной моторной коры в спинной мозг. [1] Ряд областей коры, включая премоторную кору , дополнительную двигательную область и даже первичную соматосенсорную кору, проецируются на спинной мозг. Даже когда клетки Бетца повреждены, кора все еще может связываться с подкорковыми двигательными структурами и контролировать движение. Если первичная моторная кора с клетками Бетца повреждена, возникает временный паралич, и другие области коры, очевидно, могут взять на себя часть утраченных функций.
Клиническое значение
[ редактировать ]Поражение прецентральной извилины приводит к параличу контрлатеральной стороны тела ( паралич лицевого нерва рук/ног , монопарез , гемипарез ) — см. верхний мотонейрон .
Кодирование движения
[ редактировать ]Эвартс [31] предположил, что каждый нейрон моторной коры вносит свой вклад в силу в мышце. Когда нейрон становится активным, он посылает сигнал в спинной мозг, сигнал передается мотонейрону, мотонейрон посылает сигнал мышце, и мышца сокращается. Чем больше активность нейрона двигательной коры, тем больше мышечная сила.
Георгопулос и коллеги [32] [33] [34] предположил, что мышечная сила сама по себе является слишком простым описанием. Они обучали обезьян тянуться в разных направлениях и следили за активностью нейронов моторной коры. Они обнаружили, что каждый нейрон моторной коры был максимально активен в определенном направлении и хуже реагировал на соседние направления. На этом основании они предположили, что нейроны моторной коры путем «голосования» или объединения своих влияний в « популяционный код » могут точно определять направление действия.
Предположение о том, что нейроны моторной коры кодируют направление движения, стало спорным. Скотт и Каласка [35] показали, что каждый нейрон моторной коры лучше коррелирует с деталями движения суставов и мышечной силой, чем с направлением вытягивания рук. Шварц и коллеги [36] показали, что нейроны моторной коры хорошо коррелируют со скоростью руки. Стрик и коллеги [37] обнаружили, что некоторые нейроны в моторной коре активны в связи с мышечной силой, а некоторые — с пространственным направлением движения. Тодоров [38] предположил, что множество различных корреляций являются результатом работы мышечного контроллера, в котором многие параметры движения коррелируют с мышечной силой.
Код, с помощью которого нейроны моторной коры приматов контролируют спинной мозг и, следовательно, движения, остается дискуссионным.
Некоторый конкретный прогресс в понимании того, как моторная кора вызывает движение, также был достигнут на модели грызунов. Моторная кора грызунов, как и моторная кора обезьяны, может содержать субрегионы, которые подчеркивают различные общие типы действий. [39] [40] Например, в одной области подчеркивается ритмический контроль взбивания . [39] [41] [42] Нейроны в этой области проецируются на определенное подкорковое ядро, в котором генератор паттернов координирует циклический ритм усов. Затем это ядро проецируется на мышцы, контролирующие усы.
Дополнительные изображения
[ редактировать ]

 Двигательный тракт.
Двигательный тракт.



См. также
[ редактировать ]- Кортикоспинальный тракт
- Моторная кора
- Кортикальный гомункул
- Верхний двигательный нейрон
- район Бродмана
- Список регионов человеческого мозга
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Ривара CB, Шервуд CC, Бурас C, Хоф PR (2003). «Стереологическая характеристика и закономерности пространственного распределения клеток Бетца в первичной моторной коре человека» . Анатомические записи, часть A: открытия в молекулярной, клеточной и эволюционной биологии . 270 (2): 137–151. дои : 10.1002/ar.a.10015 . ПМИД 12524689 .
- ^ Перейти обратно: а б Лассек, AM (1941). «Пирамидный тракт обезьяны». Дж. Комп. Нейрол . 74 (2): 193–202. дои : 10.1002/cne.900740202 . S2CID 83536088 .
- ^ Каллауэй, Эдвард М.; Донг, Хун-Вэй; Экер, Джозеф Р.; Гаврилич, Майкл Дж.; Хуанг, З. Джош; Лейн, Эд С.; Нгай, Джон; Остен, Павел; Рен, Бинг; Толиас, Андреас Савас; Уайт, Оуэн (октябрь 2021 г.). «Мультимодальная перепись клеток и атлас первичной моторной коры млекопитающих» . Природа . 598 (7879): 86–102. дои : 10.1038/s41586-021-03950-0 . ISSN 1476-4687 . ПМЦ 8494634 . ПМИД 34616075 .
- ^ «Просмотр разделов» . www.brainarchitecture.org . Проверено 20 ноября 2020 г.
- ^ Перейти обратно: а б Сигельбаум, Стивен А.; Хадспет, Эй Джей (2013). Принципы нейронауки . Кандел, Эрик Р. (5-е изд.). Нью-Йорк. ISBN 9780071390118 . OCLC 795553723 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Перейти обратно: а б с д Лемон, Роджер Н. (4 апреля 2008 г.). «Нисходящие пути в управлении двигателем». Ежегодный обзор неврологии . 31 (1): 195–218. дои : 10.1146/annurev.neuro.31.060407.125547 . ISSN 0147-006X . ПМИД 18558853 .
- ^ Перейти обратно: а б с Пенфилд, В. и Болдри, Э. (1937). «Соматическое моторное и сенсорное представление в коре головного мозга человека, изученное с помощью электрической стимуляции». Мозг . 60 (4): 389–443. дои : 10.1093/мозг/60.4.389 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Кван ХК, Маккей В.А., Мерфи Дж.Т., Вонг Ю.К. (1978). «Пространственная организация прецентральной коры бодрствующих приматов. II. Двигательная активность». Дж. Нейрофизиология . 41 (5): 1120–1131. дои : 10.1152/jn.1978.41.5.1120 . ПМИД 100584 .
- ^ Перейти обратно: а б Парк, М.С., Белхадж-Саиф, А., Гордон, М. и Чейни, П.Д. (2001). «Последовательные особенности представления первичной моторной коры передних конечностей у макак-резус» . Дж. Нейроски . 21 (8): 2784–2792. doi : 10.1523/JNEUROSCI.21-08-02784.2001 . ПМК 6762507 . ПМИД 11306630 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б Мейер, Дж.Д., Афлало, Т.Н., Кастнер, С. и Грациано, MSA (2008). «Сложная организация первичной моторной коры человека: исследование фМРТ высокого разрешения» . Дж. Нейрофизиология . 100 (4): 1800–1812. дои : 10.1152/jn.90531.2008 . ПМК 2576195 . ПМИД 18684903 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Бинькофски Ф., Финк Г.Р., Гейер С., Буччино Г., Грубер О., Шах Н.Дж., Тейлор Дж.Г., Зейтц Р.Дж., Зиллес К., Фрейнд Х.Дж. (2002). «Нервная активность в областях 4a и 4b первичной моторной коры человека по-разному модулируется вниманием к действию». Дж. Нейрофизиология . 88 (1): 514–519. дои : 10.1152/jn.2002.88.1.514 . hdl : 11858/00-001M-0000-0010-CA2B-E . ПМИД 12091573 . S2CID 6459190 .
- ^ Нихил Шарма; PS Джонс; Т. А. Карпентер; Жан-Клод Барон (2008). «Картирование участия BA 4a и 4p в двигательном воображении». НейроИмидж . 41 (1): 92–99. doi : 10.1016/j.neuroimage.2008.02.009 . ПМИД 18358742 . S2CID 8673179 .
- ^ Терумицу М., Икеда К., Кви И.Л. и Накада Т. (2009). «Участие области 4а первичной моторной коры в сложной сенсорной обработке: исследование фМРТ 3,0 Т». НейроОтчет . 20 (7): 679–683. дои : 10.1097/WNR.0b013e32832a1820 . ПМИД 19339906 . S2CID 23674509 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ратло, Ж.-А.; Стрик, Польша (20 января 2009 г.). «Подразделения первичной моторной коры на основе кортико-мотонейрональных клеток» . Учеб. Натл. акад. Наука . 106 (3): 918–923. Бибкод : 2009PNAS..106..918R . дои : 10.1073/pnas.0808362106 . ПМК 2621250 . ПМИД 19139417 .
- ^ Костанди, Мо (2009). «Эволюция ловкости рук» . Труды Национальной академии наук . 106 (3): 918–923. дои : 10.1073/pnas.0808362106 . ПМК 2621250 . ПМИД 19139417 . Проверено 29 ноября 2015 г.
- ^ Перейти обратно: а б Чейни, П.Д. и Фетц, Э.Э. (1985). «Сравнимые закономерности мышечного облегчения, вызываемые отдельными кортикомотонейрональными (CM) клетками и отдельными внутрикортикальными микростимулами у приматов: данные о функциональных группах клеток CM». Дж. Нейрофизиология . 53 (3): 786–804. дои : 10.1152/jn.1985.53.3.786 . ПМИД 2984354 .
- ^ Шибер, М.Х. и Хиббард, Л.С. (1993). «Насколько соматотопична область моторной коры рук?». Наука . 261 (5120): 489–492. Бибкод : 1993Sci...261..489S . дои : 10.1126/science.8332915 . ПМИД 8332915 .
- ^ Рателот, Дж. А. и Стрик, П. Л. (2006). «Представление мышц в моторной коре макака: анатомический взгляд» . Учеб. Натл. акад. наук. США . 103 (21): 8257–8262. Бибкод : 2006PNAS..103.8257R . дои : 10.1073/pnas.0602933103 . ПМЦ 1461407 . ПМИД 16702556 .
- ^ Санес Дж. Н., Донохью Дж. П., Тангарадж В., Эдельман Р. Р. и Варах С. (1995). «Общие нейронные субстраты, контролирующие движения рук в моторной коре человека». Наука . 268 (5218): 1775–1777. Бибкод : 1995Sci...268.1775S . дои : 10.1126/science.7792606 . ПМИД 7792606 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Донохью, Дж. П., Лейбович, С. и Санес, Дж. Н. (1992). «Организация области передних конечностей в моторной коре белой обезьяны: представление мышц пальцев, запястья и локтя». Эксп. Мозговой Рес . 89 (1): 1–10. дои : 10.1007/bf00228996 . ПМИД 1601087 . S2CID 1398462 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Нудо, Р.Дж., Милликен, Г.В., Дженкинс, В.М. и Мерцених, М.М. (1996). «Зависимые от использования изменения представлений о движении в первичной моторной коре взрослых обезьян» . Дж. Нейроски . 16 (2): 785–807. doi : 10.1523/JNEUROSCI.16-02-00785.1996 . ПМК 6578638 . ПМИД 8551360 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Мартин Дж. Х., Энгбер Д. и Мэн З. (2005). «Влияние использования передних конечностей на постнатальное развитие двигательного представительства передних конечностей в первичной моторной коре кошки». Дж. Нейрофизиология . 93 (5): 2822–2831. дои : 10.1152/jn.01060.2004 . ПМИД 15574795 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Грациано, MSA , Тейлор, CSR и Мур, Т. (2002). «Сложные движения, вызываемые микростимуляцией прецентральной коры» . Нейрон . 34 (5): 841–851. дои : 10.1016/S0896-6273(02)00698-0 . ПМИД 12062029 . S2CID 3069873 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б Грациано, MSA (2008). Интеллектуальная машина движения . Оксфорд, Великобритания: Издательство Оксфордского университета.
- ^ Грациано, MSA и Афлало, Теннесси (2007). «Отображение поведенческого репертуара в коре головного мозга» . Нейрон . 56 (2): 239–251. дои : 10.1016/j.neuron.2007.09.013 . ПМИД 17964243 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б Кэмпбелл, AW (1905). Гистологические исследования локализации функций головного мозга . Кембридж, Массачусетс: Издательство Кембриджского университета.
- ^ Фогт К. и Фогт О. (1919). «Результаты нашего исследования мозга». Журнал психологии и неврологии . 25 :277-462.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Ферстер, О (1936). «Моторная кора человека в свете учения Хьюлингса Джексона». Мозг . 59 (2): 135–159. дои : 10.1093/мозг/59.2.135 .
- ^ Фултон, Дж (1935). «Заметка об определении «двигательной» и «премоторной» областей». Мозг . 58 (2): 311–316. дои : 10.1093/мозг/58.2.311 .
- ^ Вулси, К.Н., Сеттлейдж, П.Х., Мейер, Д.Р., Сенсер, В., Хамуи, Т.П. и Трэвис, А.М. (1952). «Схема локализации в прецентральных и «дополнительных» двигательных областях и их отношение к понятию премоторной области». Ассоциация исследований нервных и психических заболеваний . 30 . Нью-Йорк, штат Нью-Йорк: Raven Press: 238–264.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Эвартс, Э.В. (1968). «Связь активности пирамидного тракта с силой, прилагаемой во время произвольного движения». Дж. Нейрофизиология . 31 (1): 14–27. дои : 10.1152/jn.1968.31.1.14 . ПМИД 4966614 .
- ^ Георгопулос А.П., Каласка Дж.Ф., Каминити Р. и Мэсси Дж.Т. (1982). «О связи между направлением двумерных движений рук и разрядом клеток в моторной коре приматов» . Дж. Нейроски . 2 (11): 1527–1537. doi : 10.1523/JNEUROSCI.02-11-01527.1982 . ПМК 6564361 . ПМИД 7143039 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Георгопулос А.П., Кеттнер Р.Э. и Шварц А.Б. (1988). «Моторная кора приматов и свободные движения рук к зрительным целям в трехмерном пространстве. II. Кодирование направления движения популяцией нейронов» . Дж. Нейроски . 8 (8): 2928–2937. doi : 10.1523/JNEUROSCI.08-08-02928.1988 . ПМК 6569382 . ПМИД 3411362 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Георгопулос А.П., Шварц А.Б. и Кеттнер Р.Э. (1986). «Нейрональная популяция, кодирующая направление движения». Наука . 233 (4771): 1416–1419. Бибкод : 1986Sci...233.1416G . дои : 10.1126/science.3749885 . ПМИД 3749885 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Скотт, С.Х. и Каласка, Дж.Ф. (1995). «Изменения активности моторной коры во время движений рук с одинаковыми движениями, но с разными положениями рук». Дж. Нейрофизиология . 73 (6): 2563–2567. дои : 10.1152/jn.1995.73.6.2563 . ПМИД 7666162 .
- ^ Моран, Д.В. и Шварц, AB (1999). «Моторное корковое представление скорости и направления во время достижения». Дж. Нейрофизиология . 82 (5): 2676–2692. дои : 10.1152/jn.1999.82.5.2676 . ПМИД 10561437 . S2CID 9789197 .
- ^ Какей С., Хоффман Д. и Стрик П. (1999). «Представления мышц и движений в первичной моторной коре». Наука . 285 (5436): 2136–2139. CiteSeerX 10.1.1.137.8610 . дои : 10.1126/science.285.5436.2136 . ПМИД 10497133 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Тодоров, Э (2000). «Прямой кортикальный контроль активации мышц при произвольных движениях рук: модель». Нат Нейроски . 3 (4): 391–398. дои : 10.1038/73964 . ПМИД 10725930 . S2CID 13996279 .
- ^ Перейти обратно: а б Хейсс Ф. и Шварц К. (2005). «Пространственное разделение различных способов управления движением в представлении усов первичной моторной коры крысы» . Дж. Нейроски . 25 (6): 1579–1587. doi : 10.1523/JNEUROSCI.3760-04.2005 . ПМК 6726007 . ПМИД 15703412 .
- ^ Раманатан Д., Коннер Дж. М. и Тушински М. Х. (2006). «Форма моторной кортикальной пластичности, которая коррелирует с восстановлением функций после травмы головного мозга» . Учеб. Натл. акад. наук. США . 103 (30): 11370–11375. Бибкод : 2006PNAS..10311370R . дои : 10.1073/pnas.0601065103 . ПМК 1544093 . ПМИД 16837575 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Брехт М., Шнайдер М., Сакманн Б. и Маргри Т.В. (2004). «Движения усов, вызванные стимуляцией одиночных пирамидных клеток в моторной коре крыс». Природа . 427 (6976): 704–710. Бибкод : 2004Natur.427..704B . дои : 10.1038/nature02266 . ПМИД 14973477 . S2CID 1105868 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Крамер, Н.П. и Келлер, А. (2006). «Кортикальный контроль взбивающего центрального генератора паттернов» . Дж. Нейрофизиология . 96 (1): 209–217. дои : 10.1152/jn.00071.2006 . ПМК 1764853 . ПМИД 16641387 .