Нейротубула


Нейротубулы — это микротрубочки , обнаруженные в нейронах нервных тканей. [1] Наряду с нейрофиламентами и микрофиламентами они образуют цитоскелет нейронов. Нейротубулы представляют собой неделимые полые цилиндры, состоящие из тубулина. белковых полимеров [2] и массивы, параллельные плазматической мембране нейронов. [3] Нейротубулы имеют внешний диаметр около 23 нм и внутренний диаметр, также известный как центральное ядро, около 12 нм. Стенка нейротрубочек имеет ширину около 5 нм. Вокруг нейротрубочки имеется непрозрачная прозрачная зона диаметром около 40 нм. [3] Как и микротрубочки, нейротрубочки очень динамичны, и их длину можно регулировать путем полимеризации и деполимеризации тубулина. [4]
Несмотря на схожие механические свойства, нейротрубочки отличаются от микротрубочек, обнаруженных в других типах клеток, с точки зрения их функций и внутриклеточного расположения. Большинство нейротрубочек не закреплены в организующем центре микротрубочек (MTOC), как это делают обычные микротрубочки. Вместо этого они высвобождаются для транспорта в дендриты и аксоны после их зарождения в центросоме . Следовательно, оба конца нейротрубочек оканчиваются в цитоплазме . [5]
Нейротубулы играют решающую роль в различных клеточных процессах в нейронах. Вместе с нейрофиламентами они помогают поддерживать форму нейрона и обеспечивают механическую поддержку. Нейротубулы также помогают транспортировать органеллы, пузырьки, содержащие нейротрансмиттеры , информационную РНК и другие внутриклеточные молекулы внутри нейрона. [6]
Структура и динамика
[ редактировать ]
Состав
[ редактировать ]Как и микротрубочки, нейротрубочки состоят из белковых полимеров α-тубулина и β-тубулина . α-тубулин и β-тубулин представляют собой тесно связанные глобулярные белки. Они соединяются вместе, образуя димер, называемый тубулином . Нейротубулы обычно состоят из 13 протофиламентов, которые полимеризуются из димеров тубулина. Поскольку димер тубулина состоит из одного α-тубулина и одного β-тубулина, один конец нейротрубочки открыт для α-тубулина, а другой конец для β-тубулина, эти два конца способствуют полярности нейротрубочки – плюсу. (+) конец и минус (-). Субъединица β-тубулина обнажена на плюсовом (+) конце. Два конца различаются по скорости роста: плюс (+) — это быстрорастущий конец, а минус (-) — это медленно растущий конец. Оба конца имеют собственную скорость полимеризации и деполимеризации димеров тубулина, суммарная полимеризация вызывает сборку тубулина, а значит, и длину нейротрубочек. [4]
Динамическая нестабильность
[ редактировать ]Рост нейротрубочек регулируется динамической нестабильностью . [7] Для него характерны четко выраженные фазы роста и быстрая усадка. Переход от роста к быстрому сокращению называется «катастрофой». Обратное называется «спасением».
Поляризованные массивы нейротрубочек
[ редактировать ]Нейроны имеют поляризованную сеть нейротрубочек. [8] Аксоны большинства нейронов содержат нейротрубочки с плюсовым (+) концом, равномерно направленным к окончанию аксона, и минусовым (-) концом, ориентированным к телу клетки, что аналогично общей ориентации микротрубочек в других типах клеток. С другой стороны, дендриты содержат нейротрубочки со смешанной полярностью. Половина из них направлена своим плюсовым (+) концом к вершине дендрита, а другая половина — к телу клетки, что напоминает антипараллельный массив микротрубочек митотического веретена .
Поляризованная сеть нейротрубочек формирует основу для избирательного перемещения грузов в аксоны и дендриты. [9] Например, когда происходят мутации в динеине , моторном белке , который имеет решающее значение для поддержания однородной ориентации аксональных нейротрубочек, полярность нейротрубочек в аксоне становится смешанной. [10] В результате дендритные белки неправильно доставляются в аксоны. [11]
Неполяризованные нейроны содержат 80% нейротрубочек с плюсовым (+) концом, обращенным к терминалю. [ нужна ссылка ]
Аксональный транспорт
[ редактировать ]Нейротубулы отвечают за транспортировку внутриклеточных материалов. Грузы транспортируются моторными белками, которые используют нейротрубочки в качестве «дорожки». Аксональный транспорт можно классифицировать по скорости – быстрый или медленный, а по направлению – антероградный или ретроградный.
Быстрая и медленная аксональная транспортировка
[ редактировать ]Грузы перевозятся быстрым или медленным темпом. Скорость быстрого аксонального транспорта составляет 50–500 мм в день, тогда как скорость медленного аксонального транспорта составляет 0,4 мм в день у золотых рыбок и 1–10 мм в день у нервов млекопитающих. Транспорт нерастворимого белка способствует быстрому перемещению, тогда как медленный транспорт переносит до 40–50% растворимого белка. [12] Скорость перевозки зависит от типа перевозимого груза. Нейротрофины , семейство белков, важных для выживания нейрона, а также органеллы , такие как митохондрии и эндосомы , транспортируются с высокой скоростью. Напротив, структурные белки, такие как субъединицы тубулина и нейрофиламентов, транспортируются с более низкой скоростью. Белкам, которые транспортируются из спинного мозга в стопы, может потребоваться до года, чтобы завершить путешествие. [13]
Антероградный транспорт и ретроградный транспорт
[ редактировать ]
Антероградный транспорт относится к перевозке грузов от минусового (-) конца к плюсовому (+), тогда как ретроградный транспорт - это транспортировка грузов в противоположном направлении. Антероградный транспорт часто представляет собой транспортировку от тела клетки к периферии нейрона, тогда как ретроградный транспорт переносит органеллы и пузырьки от окончания аксона к телу клетки.
Антероградный транспорт регулируется кинезинами , классом моторных белков. Кинезины имеют два домена головы, которые работают вместе, как ноги: один связывается с нейротрубочками, а затем другой связывается, а первый диссоциирует. Связывание АТФ повышает сродство кинезинов к нейротрубочкам. Когда АТФ связывается с одним доменом головки, в головном домене индуцируются конформационные изменения, в результате чего он прочно связывается с нейротрубочкой. Затем другой АТФ связывается с другим головным доменом, в то время как первый АТФ гидролизуется, а головной домен диссоциирует. Процесс повторяется циклически, так что кинезины движутся по нейротрубочкам вместе с органеллами и везикулярными грузами, которые они несут. [14]
Ретроградный транспорт регулируется динеинами , также классом моторных белков. Он имеет схожую структуру с кинезинами, а также механизм транспортировки. Он транспортирует грузы от периферии к телу клетки в нейронах.
Белки, связанные с нейротрубочками
[ редактировать ]
Белки, ассоциированные с микротрубочками (MAP), представляют собой белки, которые взаимодействуют с микротрубочками путем связывания с их субъединицами тубулина и регулирования их стабильности. Состав MAP нейротрубочек заметно отличается от микротрубочек ненейрональных клеток. [15] Например, MAP типа II обнаруживаются исключительно в нейронах, а не в других клетках. К наиболее хорошо изученным относятся MAP2 и тау .
MAPs дифференциально распределены в цитоплазме нейронов. Их распределение также варьируется на разных стадиях развития нейрона. Ювенильная изоформа MAP2 присутствует в нейротрубочках аксонов и дендритах развивающихся нейронов, но ее активность снижается по мере взросления нейронов. Взрослая изоформа MAP2 обогащена нейротрубочками дендритов и практически отсутствует в аксональных нейротрубочках. [16] Напротив, тау отсутствует в нейротрубочках дендритов и его присутствие ограничено аксональными нейротрубочками. Фосфорилирование . тау в определенных участках необходимо для связывания тау с нейротрубочками В здоровом нейроне этот процесс в дендритах в значительной степени не происходит, что приводит к отсутствию тау в дендритных нейротрубочках. Связывание тау разных изоформ и разного уровня фосфорилирования регулирует стабильность нейротрубочек. Установлено, что нейротрубочки нейронов эмбриональной центральной нервной системы содержат более высокофосфорилированный тау, чем у взрослых. [17] Кроме того, тау отвечает за связывание нейротрубочек. [18]
Белки отслеживания плюсовых концов микротрубочек (+TIP) представляют собой MAP, которые накапливаются на плюсовых концах микротрубочек. В нейротрубочках +TIPs контролируют динамику нейротрубочек, направление роста и взаимодействие с компонентами клеточной коры . Они важны для расширения нейритов и роста аксонов . [19]
Многие другие неспецифические для нейронов MAP, такие как MAP1B и MAP6 , обнаружены в нейротрубочках. Более того, взаимодействие между актином и некоторыми MAP обеспечивает потенциальную связь между нейротрубочками и актиновыми нитями . [20]
Неврологические расстройства, связанные с нейротрубочками
[ редактировать ]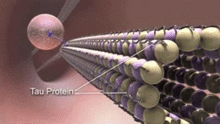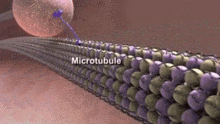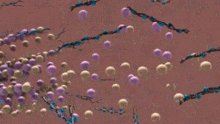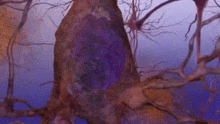
Нарушение целостности и динамики нейротрубочек может нарушать выполняемые ими клеточные функции и вызывать различные неврологические расстройства .
болезнь Альцгеймера
[ редактировать ]При Альцгеймера болезни гиперфосфорилирование тау -белка вызывает диссоциацию тау от нейротрубочек и неправильное сворачивание тау . Агрегация неправильно свернутого тау образует нерастворимые нейрофибриллярные клубки , что является характерным признаком болезни Альцгеймера. [21] Это патологическое изменение называется таупатией . Нейротубулы становятся склонными к распаду белками, разрывающими микротрубочки, когда тау диссоциирует. [22] В результате важные процессы в нейроне, такие как аксональный транспорт и нейронная связь, будут нарушены, что станет основой нейродегенерации . [23] Считается, что распад нейротрубочек происходит по разным механизмам в аксонах и дендритах.
Отслойка тау дестабилизирует нейротрубочки, позволяя катанину избыточно разрывать их , вызывая их распад. Распад нейротрубочек в аксоне нарушает транспорт мРНК и сигнальных молекул к окончанию аксона. [22] Что касается дендритов, новые данные свидетельствуют о том, что аномальная инвазия тау в дендриты вызывает повышенный уровень дендритного TTLL6 (Tubulin-Tyrosine-Ligase-Like-6), что повышает статус полиглутамилирования нейротрубочек в дендритах. [22] Поскольку спастин отдает предпочтение полиглутамилированным микротрубочкам, дендритные нейротрубочки становятся восприимчивыми к дезинтеграции, индуцированной спастином. [22] Утрата сети нейротрубочек в дендритах и аксонах наряду с образованием нейрофибриллярных клубков приводит к нарушению транспортировки важных грузов через клетку, что в конечном итоге может привести к апоптозу . [24]
Лиссэнцефалия
[ редактировать ]
Лиссэнцефалия — редкое врожденное заболевание, при котором головной мозг теряет свои складки ( извилины ) и бороздки ( бороздки ), в результате чего поверхность мозга кажется гладкой. Это вызвано миграцией дефектных нейронов . [25] Неспособность постмитотических нейронов занять правильное положение приводит к образованию дезорганизованного и утолщенного четырехслойного неокортекса вместо нормального шестислойного неокортекса . Тяжесть лиссэнцефалии варьирует от полной утраты складок головного мозга ( агирия ) до общего уменьшения корковых складок ( пахигирия ).
Нейротубула играет центральную роль в механизме миграции нейронов. Нарушенная миграция нейронов у людей, страдающих лиссэнцефалией, вызвана мутациями, связанными с генами, связанными с нейротрубочками, такими как LIS1 и DCX . [26] LIS1 кодирует адаптерный белок Lis1, который отвечает за стабилизацию нейротрубочек во время миграции нейронов путем минимизации катастрофы нейротрубочек. Он также регулирует моторный белок динеин, который имеет решающее значение для транслокации ядра по нейротрубочкам. Это действие продвигает сому нейрона вперед, что является важным шагом в миграции нейронов. [27] Кроме того, , что мутации в LIS1 нарушают единую плюс-концевую-дистальную полярность аксонов на животных моделях, вызывая ошибочный перенос дендритных белков в аксоны. обнаружено [11] С другой стороны, DCX кодирует белок даблкортин , который взаимодействует с Lis1, помимо поддержки структуры 13 протофиламентов нейротрубочек.
Периферическая невропатия, вызванная химиотерапией
[ редактировать ]Периферическая нейропатия, индуцированная химиотерапией, — патологическое изменение нейронов, вызванное нарушением динамики нейротрубочек химиопрепаратами, проявляющееся болью, онемением, ощущением покалывания и мышечной слабостью в конечностях. Это необратимое состояние, от которого страдает около трети пациентов, проходящих химиотерапию. [28] Ингибиторы тубулина ингибируют митоз в раковых клетках, влияя на стабильность и динамику микротрубочек, которые образуют митотическое веретено, ответственное за сегрегацию хромосом во время митоза, подавляя рост опухоли .
Однако те же препараты влияют и на нейротрубочки в нейронах. Винбластин связывается со свободными тубулинами и снижает их способность к полимеризации, способствуя деполимеризации нейротрубочек. С другой стороны, паклитаксел связывается с кепкой нейротрубочек, что предотвращает превращение связанного с тубулином ГТФ в ВВП — процесс, который способствует деполимеризации нейротрубочек. Для нейронов in vitro , обработанных паклитакселом, нарушается паттерн полярности нейротрубочек, что может привести к долгосрочному повреждению нейронов. Кроме того, чрезмерная стабилизация нейротрубочек препятствует их способности выполнять важные клеточные функции в нейронах. [29]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Медицинское определение НЕЙРОТУБУЛЫ» . www.merriam-webster.com . Проверено 25 марта 2019 г.
- ^ Нервная система человека: строение и функции . Нобак, Чарльз Р. (Чарльз Роберт), 1916–2009 гг. (6-е изд.). Тотова, Нью-Джерси: Humana Press. 2005. ISBN 1588290395 . OCLC 222291397 .
{{cite book}}: CS1 maint: другие ( ссылка ) - ^ Перейти обратно: а б Лейн, Нью-Джерси, Трехерн Дж. Э. (июль 1970 г.). «Окрашивание нейротрубочек в аксонах тараканьих ганглиев лантаном». Журнал клеточной науки . 7 (1): 217–31. дои : 10.1242/jcs.7.1.217 . ПМИД 4097712 .
- ^ Перейти обратно: а б Нейрональная специфичность, пластичность и закономерности . Москона, А.А. (Арон Артур), 1922–2009 гг., Монрой, Альберто, Хант, Р. Кевин. Нью-Йорк: Академическая пресса. 1982. ISBN 9780080584409 . OCLC 276661314 .
{{cite book}}: CS1 maint: другие ( ссылка ) - ^ Купер, Джеффри М. (2000). «Микротрубочки» . Клетка: молекулярный подход. 2-е издание .
- ^ Медведь М.Ф., Коннорс Б.В., Парадсо Массачусетс. Нейронаука: исследование мозга (3-е изд.). Липпинкотт Уильямс и Уилкинс. п. 41.
- ^ Киршнер, Марк; Митчисон, Тим (ноябрь 1984 г.). «Динамическая нестабильность роста микротрубочек». Природа . 312 (5991): 237–242. Бибкод : 1984Natur.312..237M . дои : 10.1038/312237a0 . ISSN 1476-4687 . ПМИД 6504138 . S2CID 30079133 .
- ^ Кван AC, Домбек Д.А., Уэбб В.В. (август 2008 г.). «Поляризованные массивы микротрубочек в апикальных дендритах и аксонах» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (32): 11370–5. Бибкод : 2008PNAS..10511370K . дои : 10.1073/pnas.0805199105 . ПМК 2496886 . ПМИД 18682556 .
- ^ ван Бёнинген С.Ф., Хугенраад CC (август 2016 г.). «Нейрональная полярность: ремоделирование организации микротрубочек». Современное мнение в нейробиологии . 39 : 1–7. дои : 10.1016/j.conb.2016.02.003 . ПМИД 26945466 . S2CID 3340102 .
- ^ Конде С., Касерес А (май 2009 г.). «Сборка, организация и динамика микротрубочек в аксонах и дендритах». Обзоры природы. Нейронаука . 10 (5): 319–32. дои : 10.1038/nrn2631 . ПМИД 19377501 . S2CID 205504854 .
- ^ Перейти обратно: а б Дубей Дж., Ратнакаран Н., Кошика С.П. (2015). «Нейродегенерация и динамика микротрубочек: смерть от тысячи порезов» . Границы клеточной нейронауки . 9 : 343. дои : 10.3389/fncel.2015.00343 . ПМЦ 4563776 . ПМИД 26441521 .
- ^ Графштейн Б., МакИвен Б.С., Шелански М.Л. (июль 1970 г.). «Аксональный транспорт белка нейротрубочек». Природа . 227 (5255): 289–90. Бибкод : 1970Natur.227..289G . дои : 10.1038/227289a0 . ПМИД 4193613 . S2CID 4253835 .
- ^ «Являются ли микротрубочки мозгом нейрона» . Джон Лифф, доктор медицинских наук . 29 ноября 2015 г. Проверено 10 апреля 2019 г.
- ^ Берг, Джереми М. (2002). Биохимия . Тимочко, Джон Л., 1948-, Страйер, Люберт., Страйер, Люберт. (5-е изд.). Нью-Йорк: WH Freeman. ISBN 0716730510 . OCLC 48055706 .
- ^ Аль-Бассам Дж., Озер Р.С., Сафер Д., Халпейн С., Миллиган Р.А. (июнь 2002 г.). «MAP2 и тау связываются продольно вдоль внешних гребней протофиламентов микротрубочек». Дж. Клеточная Биол . 157 (7): 1187–96. дои : 10.1083/jcb.200201048. ПМЦ 2173547. ПМИД 12082079.
- ^ Демельт Л., Халпейн С. (2005). «Семейство MAP2/Tau белков, связанных с микротрубочками» . Геномная биология . 6 (1): 204. doi : 10.1186/gb-2004-6-1-204 . ПМК 549057 . ПМИД 15642108 .
- ^ Канемару К., Такио К., Миура Р., Титани К., Ихара Ю. (май 1992 г.). «Фосфорилирование тау по фетальному типу в парных спиральных нитях». Журнал нейрохимии. 58 (5): 1667–75. doi:10.1111/j.1471-4159.1992.tb10039.x. ПМИД 1560225.
- ^ Мандельков Э., Мандельков Э.М. (февраль 1995 г.). «Микротрубочки и белки, связанные с микротрубочками». Курс. Мнение. Клеточная Биол . 7 (1): 72–81. дои : 10.1016/0955-0674(95)80047-6. ПМИД 7755992.
- ^ Ахманова А, Hoogenraad CC (февраль 2005 г.). «Белки, отслеживающие плюс-концы микротрубочек: механизмы и функции». Современное мнение в области клеточной биологии . 17 (1): 47–54. дои : 10.1016/j.ceb.2004.11.001 . ПМИД 15661518 .
- ^ Майерс, Кеннет А.; Баас, Питер В. (2011), Галло, Джанлука; Ланье, Лорен М. (ред.), «Взаимодействие микротрубочек и актина во время развития нейронов», Нейробиология актина: от нейруляции к синаптической функции , Достижения в нейробиологии, том. 5, Springer New York, стр. 73–96, номер документа : 10.1007/978-1-4419-7368-9_5 , ISBN. 9781441973689
- ^ Икбал К., Лю Ф., Гонг CX, Грундке-Икбал I (декабрь 2010 г.). «Тау при болезни Альцгеймера и связанных с ней таупатиях» . Текущие исследования болезни Альцгеймера . 7 (8): 656–64. дои : 10.2174/156720510793611592 . ПМК 3090074 . ПМИД 20678074 .
- ^ Перейти обратно: а б с д Жан Д.К., Баас П.В. (ноябрь 2013 г.). «Это имеет два последствия: потеря микротрубочек при болезни Альцгеймера» . Журнал ЭМБО . 32 (22): 2900–2. дои : 10.1038/emboj.2013.219 . ПМЦ 3831311 . ПМИД 24076651 .
- ^ Ван Ю, Мандельков Э (январь 2016 г.). «Тау в физиологии и патологии». Обзоры природы. Нейронаука . 17 (1): 5–21. дои : 10.1038/nrn.2015.1 . ПМИД 26631930 . S2CID 30614958 .
- ^ Мудхер А., Лавстон С. (январь 2002 г.). «Болезнь Альцгеймера – тауисты и баптисты наконец пожмут друг другу руки?». Тенденции в нейронауках . 25 (1): 22–6. дои : 10.1016/S0166-2236(00)02031-2 . ПМИД 11801334 . S2CID 37380445 .
- ^ Добинс В.Б. (1987). «Аспекты развития лиссэнцефалии и синдромов лиссэнцефалии». Оригинальная серия статей о врожденных дефектах . 23 (1): 225–41. ПМИД 3472611 .
- ^ Мочида Г.Х. (сентябрь 2009 г.). «Генетика и биология микроцефалии и лиссэнцефалии» . Семинары по детской неврологии . 16 (3): 120–6. дои : 10.1016/j.spen.2009.07.001 . ПМК 3565221 . ПМИД 19778709 .
- ^ Танака Т., Сернео ФФ, Хиггинс С., Гамбелло М.Дж., Уиншоу-Борис А., Глисон Дж.Г. (июнь 2004 г.). «Lis1 и даблкортин вместе с динеином обеспечивают соединение ядра с центросомой при миграции нейронов» . Журнал клеточной биологии . 165 (5): 709–21. дои : 10.1083/jcb.200309025 . ПМК 2172383 . ПМИД 15173193 .
- ^ Браун Т.Дж., Седхом Р., Гупта А. (февраль 2019 г.). «Периферическая невропатия, вызванная химиотерапией» . JAMA Онкология . 5 (5): 750. doi : 10.1001/jamaoncol.2018.6771 . ПМИД 30816956 .
- ^ Баас П.В., Ахмад Ф.Дж. (октябрь 2013 г.). «Помимо таксола: лечение заболеваний и повреждений нервной системы на основе микротрубочек» . Мозг . 136 (Часть 10): 2937–51. дои : 10.1093/brain/awt153 . ПМЦ 3784279 . ПМИД 23811322 .