Проект генома шимпанзе
Проект « Геном шимпанзе» был попыткой определить ДНК последовательность шимпанзе генома . Секвенирование началось в 2005 году, и к 2013 году было секвенировано двадцать четыре особи шимпанзе. Этот проект был включен в проект «Геном великой обезьяны» . [1]

В 2013 году были опубликованы последовательности высокого разрешения каждого из четырех признанных [2] [3] подвиды шимпанзе: Центральный шимпанзе , Pan troglodytes troglodytes , 10 последовательностей; Западный шимпанзе , Pan troglodytes verus , 6 последовательностей; Нигерийско-камерунский шимпанзе , Pan troglodytes ellioti , 4 последовательности; и восточный шимпанзе , Pan troglodytes schweinfurthii , 4 последовательности. Все они были секвенированы со средним 25-кратным охватом на человека. [1]
Исследование показало значительное разнообразие генома шимпанзе со многими популяционно-специфичными чертами. Центральные шимпанзе сохраняют наибольшее разнообразие в линии шимпанзе, тогда как другие подвиды демонстрируют признаки узких мест в популяциях . [4]
Фон
[ редактировать ]человека и шимпанзе Хромосомы очень похожи. Основное отличие состоит в том, что у людей на одну пару хромосом меньше, чем у других человекообразных обезьян . У человека 23 пары хромосом, а у других человекообразных обезьян — 24 пары хромосом. В эволюционной линии человека две предковые хромосомы обезьяны слились в своих теломерах , образуя человеческую хромосому 2 . [5] Есть девять других основных хромосомных различий между шимпанзе и людьми: инверсии сегментов хромосом человека 1 , 4 , 5 , 9 , 12 , 15 , 16 , 17 и 18 . После завершения проекта «Геном человека» был начат проект общего генома шимпанзе . В декабре 2003 года предварительный анализ 7600 генов, общих для двух геномов, подтвердил, что некоторые гены, такие как P2 фактор транскрипции , который участвует в развитии речи, различаются в человеческой линии. Также было обнаружено, что несколько генов, участвующих в слухе, изменились в ходе эволюции человека, что позволяет предположить, что отбор включал поведение, связанное с человеческим языком . По оценкам, различия между отдельными людьми и обычными шимпанзе примерно в 10 раз превышают типичную разницу между парами людей. [6]
Другое исследование показало, что закономерности метилирования ДНК, которые являются известным механизмом регуляции экспрессии генов, различаются в префронтальной коре человека и шимпанзе, и это различие привело к эволюционному расхождению двух видов. [7]

Проект последовательности генома обыкновенного шимпанзе
[ редактировать ]Анализ последовательности генома шимпанзе был опубликован в журнале Nature 1 сентября 2005 года в статье, подготовленной Консорциумом по секвенированию и анализу шимпанзе , группой ученых, частично поддерживаемой Национальным институтом исследования генома человека , одним из Национальных исследовательских институтов генома человека. Институты здоровья . Статья ознаменовала завершение проекта секвенирования генома. [6]
В настоящее время существует база данных, содержащая генетические различия между генами человека и шимпанзе, содержащая около тридцати пяти миллионов однонуклеотидных изменений, пять миллионов событий вставки/делеции и различные хромосомные перестройки . [10] Дупликация генов объясняет большую часть различий в последовательностях между людьми и шимпанзе. Замены одной пары оснований вызывают примерно вдвое меньше генетических изменений, чем дупликация генов.
человека и шимпанзе Типичные гомологи белков двум различаются в среднем только по аминокислотам . Около 30 процентов всех белков человека идентичны по последовательности соответствующему белку шимпанзе. Как упоминалось выше, дупликации генов являются основным источником различий между генетическим материалом человека и шимпанзе: около 2,7 процентов генома в настоящее время представляют собой различия, возникшие в результате дупликации или делеции генов в течение примерно 6 миллионов лет. [11] поскольку люди и шимпанзе разошлись от своего общего эволюционного предка. Сопоставимая вариация внутри человеческих популяций составляет 0,5 процента. [12]
Было идентифицировано около 600 генов, которые, возможно, подвергались сильному положительному отбору в линиях человека и шимпанзе; многие из этих генов участвуют в защите иммунной системы от микробных заболеваний (пример: гранулизин защищает от микобактерий туберкулеза). [13] ) или являются мишенными рецепторами патогенных микроорганизмов (пример: гликофорин С и Plasmodium falciparum ). Сравнивая гены человека и шимпанзе с генами других млекопитающих, было обнаружено, что гены, кодирующие факторы транскрипции , такие как forkhead-box P2 ( FOXP2 ), часто развиваются быстрее у человека по сравнению с шимпанзе; относительно небольшие изменения в этих генах могут объяснить морфологические различия между людьми и шимпанзе. Набор из 348 генов транскрипционных факторов кодирует белки, в которых аминокислотных изменений в человеческой линии в среднем примерно на 50 процентов больше, чем в линии шимпанзе.
Было обнаружено шесть хромосомных регионов человека, которые, возможно, находились под особенно сильным и скоординированным отбором в течение последних 250 000 лет. Эти области содержат по крайней мере один маркерный аллель , который кажется уникальным для человеческого происхождения, в то время как вся хромосомная область демонстрирует меньшую, чем обычно, генетическую изменчивость. Эта закономерность предполагает, что один или несколько тщательно отобранных генов в этой хромосомной области могли предотвращать случайное накопление нейтральных изменений в других близлежащих генах. Одна такая область на хромосоме 7 содержит ген FOXP2 (упомянутый выше), и эта область также включает ген регулятора трансмембранной проводимости муковисцидоза (CFTR), который важен для транспорта ионов в таких тканях, как секретирующий соль эпителий потовых желез. Человеческие мутации в гене CFTR могут быть выбраны как способ выжить при холере . [14]
Другая такая область на хромосоме 4 может содержать элементы, регулирующие экспрессию близлежащего гена протокадгерина , который может быть важен для развития и функционирования мозга . Хотя изменения в экспрессии генов, экспрессируемых в головном мозге, в среднем меньше, чем в других органах (например, в печени), изменения в экспрессии генов в мозге были более драматичными у человека, чем у шимпанзе. [15] Это согласуется с резким расхождением уникального паттерна развития человеческого мозга, наблюдаемого в человеческой линии, по сравнению с паттерном развития великих обезьян. Кластер генов протокадгерина-бета на хромосоме 5 также демонстрирует доказательства возможного положительного отбора. [16]
Результаты анализа генома человека и шимпанзе должны помочь в понимании некоторых заболеваний человека. Люди, по-видимому, утратили функциональный ген каспазы 12 , который у других приматов кодирует фермент, который может защитить от болезни Альцгеймера .
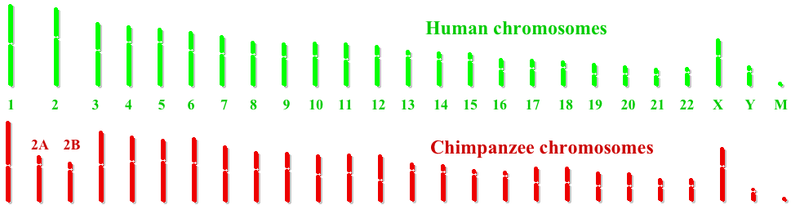
Гены места слияния хромосомы 2
[ редактировать ]
Результаты проекта генома шимпанзе показывают, что когда предковые хромосомы 2A и 2B слились с образованием хромосомы 2 человека, ни один ген не был потерян из слитых концов 2A и 2B. В месте слияния имеется примерно 150 000 пар оснований последовательности, не встречающейся в хромосомах 2А и 2В шимпанзе. Дополнительные связанные копии генов PGML/FOXD/CBWD существуют в других местах генома человека, особенно вблизи p-конца хромосомы 9 . Это предполагает, что копия этих генов могла быть добавлена к концу предкового 2A или 2B до события слияния. Еще предстоит определить, дают ли эти вставленные гены селективное преимущество.
- ПГМ5П4 . Псевдоген фосфоглюкомутазы хромосомы 2 человека. Этот ген неполный и не производит функционального транскрипта. [17]
- ФОКСД4Л1 . Ген , подобный D4, является примером безинтронного гена. Функция этого гена неизвестна, но он может кодировать белок контроля транскрипции.
- CBWD2 . Кобаламинсинтетаза представляет собой бактериальный фермент, вырабатывающий витамин B12 . В далеком прошлом общий предок мышей и обезьян содержал копию гена кобаламинсинтетазы (см.: Горизонтальный перенос генов ). Люди необычны тем, что у них есть несколько копий генов, подобных кобаламинсинтетазе, включая одну на хромосоме 2. Еще предстоит определить, какова функция этих человеческих генов, подобных кобаламинсинтетазе. Если эти гены участвуют в метаболизме витамина B12 , это может иметь отношение к эволюции человека. Основным изменением в развитии человека является больший постнатальный рост мозга, чем у других обезьян. Витамин B 12 важен для развития мозга, а дефицит витамина B 12 в процессе развития мозга приводит к тяжелым неврологическим дефектам у детей.
- ВАС2П . несколько транскриптов неизвестной функции, Было выделено соответствующих этой области. Эта область также присутствует в близкородственной концевой области хромосомы 9p, которая содержит копии генов PGML/FOXD/CBWD.
- РПЛ23АП7 . Многие рибосомного белка L23a псевдогены разбросаны по геному человека.
См. также
[ редактировать ]Дальнейшее чтение
[ редактировать ]- Сунцова, М.В.; Буздин А.А. (10 сентября 2020 г.). «Различия между геномами человека и шимпанзе и их влияние на экспрессию генов, функции белков и биохимические свойства двух видов» . БМК Геномика . 21 (535): 535. doi : 10.1186/s12864-020-06962-8 . ПМЦ 7488140 . ПМИД 32912141 .
Ссылки
[ редактировать ]- ^ Jump up to: а б Прадо-Мартинес, Дж.; и др. (2013). «Генетическое разнообразие и история популяций великих обезьян» . Природа . 499 (7459): 471–475. Бибкод : 2013Natur.499..471P . дои : 10.1038/nature12228 . ПМЦ 3822165 . ПМИД 23823723 .

- ^ Гроувс, Колин П. (2001). Систематика приматов . Вашингтон, округ Колумбия: Издательство Смитсоновского института. стр. 303–307. ISBN 978-1-56098-872-4 .
- ^ Хоф, Дж.; Соммер, В. (2010). Обезьяны, подобные нам: портреты родства . Мангейм: Панорама. п. 114. ИСБН 978-3-89823-435-1 .
- ^ де Мануэль, М.; и др. (2016). «Геномное разнообразие шимпанзе обнаруживает древнюю смесь с бонобо» . Наука . 354 (6311): 477–48. Бибкод : 2016Sci...354..477D . дои : 10.1126/science.aag2602 . ПМЦ 5546212 . ПМИД 27789843 .
- ^ Де Груши Дж. (август 1987 г.). «Хромосомная филогения человека, человекообразных обезьян и обезьян Старого Света». Генетика . 73 (1–2): 37–52. дои : 10.1007/bf00057436 . ПМИД 3333352 . S2CID 1098866 .
- ^ Jump up to: а б Секвенирование шимпанзе; Аналитический консорциум (2005). «Исходная последовательность генома шимпанзе и сравнение с геномом человека» (PDF) . Природа . 437 (7055): 69–87. Бибкод : 2005Natur.437...69. . дои : 10.1038/nature04072 . ПМИД 16136131 .
- ^ Цзэн, Дж.; Конопа, Г.; Хант, Б.Г.; Прейсс, ТМ; Гешвинд, Д.; Йи, СВ (2012). «Дивергентные карты метилирования всего генома мозга человека и шимпанзе раскрывают эпигенетическую основу регуляторной эволюции человека» . Американский журнал генетики человека . 91 (3): 455–465. дои : 10.1016/j.ajhg.2012.07.024 . ПМК 3511995 . ПМИД 22922032 .
- ^ МакКонки Э.Х. (2004). «Ортологическая нумерация хромосом человекообразных обезьян и человека необходима для сравнительной геномики» . Цитогенет. Геном Рез . 105 (1): 157–8. дои : 10.1159/000078022 . ПМИД 15218271 . S2CID 11571357 .
- ^ Спрингер М.С., Мерфи В.Дж., Эйзирик Э., О'Брайен С.Дж. (февраль 2003 г.). «Диверсификация плацентарных млекопитающих и граница мелового и третичного периодов» . Учеб. Натл. акад. наук. США . 100 (3): 1056–61. Бибкод : 2003PNAS..100.1056S . дои : 10.1073/pnas.0334222100 . ПМК 298725 . ПМИД 12552136 .
- ^ «База данных геномов шимпанзе (Genome Data Viewer Pan troglodytes (шимпанзе))» .
- ^ Касвелл Дж.Л., Маллик С., Рихтер Д.Д., Нойбауэр Дж., Ширмер С., Гнерр С., Райх Д. (апрель 2008 г.). «Анализ истории шимпанзе на основе выравнивания последовательностей генома» . ПЛОС Генет . 4 (4): e1000057. дои : 10.1371/journal.pgen.1000057 . ПМК 2278377 . ПМИД 18421364 .
- ^ Ченг З., Вентура М., Ше Х., Хайтович П., Грейвс Т., Осоэгава К. и др. (сентябрь 2005 г.). «Полногеномное сравнение недавних сегментных дупликаций шимпанзе и человека». Природа . 437 (7055): 88–93. Бибкод : 2005Natur.437...88C . дои : 10.1038/nature04000 . ПМИД 16136132 . S2CID 4420359 .
- ^ Стенгер С., Хансон Д.А., Тейтельбаум Р., Деван П., Ниязи К.Р., Фролих С.Дж. и др. (октябрь 1998 г.). «Антимикробная активность цитолитических Т-клеток, опосредованная гранулизином». Наука . 282 (5386): 121–5. Бибкод : 1998Sci...282..121S . дои : 10.1126/science.282.5386.121 . ПМИД 9756476 .
- ^ Гудман Б.Е., Перси WH (июнь 2005 г.). «CFTR при муковисцидозе и холере: от мембранного транспорта к клинической практике». Adv Physiol Educ . 29 (2): 75–82. дои : 10.1152/advan.00035.2004 . ПМИД 15905150 .
- ^ Хайтович П., Хеллманн И., Энард В., Новик К., Лейнвебер М., Франц Х., Вайс Г., Лахманн М., Паабо С. (сентябрь 2005 г.). «Параллельные закономерности эволюции геномов и транскриптомов человека и шимпанзе». Наука . 309 (5742): 1850–4. Бибкод : 2005Sci...309.1850K . дои : 10.1126/science.1108296 . ПМИД 16141373 . S2CID 16674740 .
- ^ Мики Р., Хаттори К., Тагучи Ю., Тада М.Н., Исосака Т., Хидака Ю., Хирабаяши Т., Хашимото Р., Фукудзако Х., Яги Т. (апрель 2005 г.). «Идентификация и характеристика кодирующих однонуклеотидных полиморфизмов в кластерах генов протокадгерина-альфа и -бета человека» . Джин . 349 : 1–14. дои : 10.1016/j.gene.2004.11.044 . ПМИД 15777644 .
- ^ Фан Ю, Ньюман Т., Линардопулу Э., Траск Б.Дж. (ноябрь 2002 г.). «Содержание генов и функция места слияния предковых хромосом в хромосоме человека 2q13-2q14.1 и паралогичных областях» . Геном Рез . 12 (11): 1663–72. дои : 10.1101/гр.338402 . ЧВК 187549 . ПМИД 12421752 .