Митохондриальный противовирусный сигнальный белок
| МАВС | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | MAVS , CARDIF, IPS-1, IPS1, VISA, митохондриальный противовирусный сигнальный белок | ||||||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | Опустить : 609676 ; МГИ : 2444773 ; Гомологен : 17004 ; Генные карты : МАВС ; ОМА : МАВС – ортологи | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||





Митохондриальный противовирусный сигнальный белок ( MAVS ) — это белок , который необходим для врожденного противовирусного иммунитета . MAVS расположен на внешней мембране митохондрий, пероксисомах и мембране митохондриально-ассоциированного эндоплазматического ретикулума (МАМ). [5] [6] При вирусной инфекции группа цитозольных белков обнаруживает присутствие вируса и связывается с MAVS, тем самым активируя MAVS. Активация MAVS приводит к тому, что инфицированная вирусом клетка начинает секретировать цитокины . Это вызывает иммунный ответ , который убивает инфицированные вирусом клетки хозяина, что приводит к уничтожению вируса.
Структура
[ редактировать ]
MAVS также известен как стимулятор промотора IFN-β I (IPS-1), адаптер домена рекрутирования активации каспазы, индуцирующий IFN-β (CARDIF), или вирус-индуцируемый сигнальный адаптер (VISA). [7] MAVS кодируется MAVS геном . [7] [8] MAVS представляет собой белок из 540 аминокислот, который состоит из трех компонентов: домена рекрутирования N-концевой активации каспазы (CARD), богатого пролином домена и трансмембранного C-концевого домена (TM). [7]
После того, как ген MAVS транскрибируется в РНК, рибосомы могут транслировать белок MAVS с двух разных сайтов. [7] Начальный сайт трансляции генерирует полноразмерный белок MAVS. Альтернативный сайт трансляции генерирует более короткий белок, называемый «miniMAVS» или short-MAVS (sMAVS). [7] sMAVS представляет собой белок MAVS из 398 аминокислот, в котором отсутствует домен CARD. Это важно, поскольку в домене CARD два цитозольных белка связываются, активируя MAVS, сигнализируя о присутствии вируса в клетке. [7]
Функция
[ редактировать ]Вирусы с двухцепочечной РНК распознаются либо трансмембранным toll-подобным рецептором 3 ( TLR3 ), либо одним из двух цитозольных белков: рецепторами, индуцируемыми ретиноевой кислотой гена I ( RIG-I ), и геном 5, связанным с дифференцировкой меланомы ( MDA5 ). . [7] [8] [9] [10] RIG-I и MDA5 различаются по вирусной РНК, которую они распознают, но у них много общих структурных особенностей, включая N-концевой CARD, который позволяет им связываться с MAVS. [7] Активация MAVS приводит к повышению уровня провоспалительных цитокинов посредством активации факторов транскрипции, ядерного фактора kB ( NF-κB ), регуляторного фактора интерферона 1 ( IRF1 ) и регуляторного фактора интерферона 3 ( IRF3 ). [7] [8] [9] NF-κB, IRF1 и IRF3 являются факторами транскрипции и играют решающую роль в выработке цитокинов.
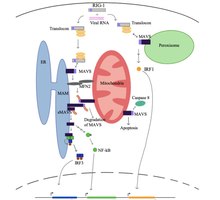
в состоянии покоя клетки белок митофузин 2 (MFN2) взаимодействует с MAVS, предотвращая связывание MAVS с цитозольными белками, такими как RIG-I и MDA5. Известно, что [7] [8] После распознавания вируса в цитозоле мембраны ER, связанные с митохондриями (MAM), и митохондрии становятся физически связанными с помощью MFN2, а RIG-I связывается со вторым белком RIG-I, образуя белковый комплекс. [7] [8] [9] Этот комплекс связывается с TRIM25 и молекулярным шапероном 14-3-3e, образуя комплекс, называемый «транслокон». [7] [8] [9] [10] Транслокон перемещается в митохондрии, где связывается с регионом CARD на MAVS, что приводит к активации MAVS. [7] [8] [9] [10] Впоследствии белки MAVS связываются друг с другом через домены CARD и TM, рекрутируя несколько нижестоящих сигнальных факторов для формирования сигнального комплекса MAVS. [7] [8] Формированию этого сигнального комплекса MAVS способствует повышенный уровень митохондриальных активных форм кислорода (mROS) , независимо от восприятия РНК. [8] [9] Сигнальный комплекс MAVS взаимодействует с TANK-связывающей киназой 1 и/или протеинкиназами IKKA (CHUK) и IKKB (IKBKB), что приводит к фосфорилированию и ядерной транслокации IRF3. [7] Хотя передача и регуляция сигнала MAVS до конца не изучены, активированные белки MAVS в митохондриях, ЭР и пероксисомах необходимы для максимизации противовирусного врожденного иммунного ответа.
Белок MAVS индуцирует апоптоз в инфицированных вирусом клетках хозяина путем взаимодействия с протеазой, называемой каспазой 8 . [7] Активация апоптоза каспазой 8 не зависит от апоптотического пути Bax/Bak, основного пути апоптоза в клетках. [7]
Вирусное уклонение
[ редактировать ]Некоторые вирусы, такие как цитомегаловирус человека (ЦМВ) и гепатит С (ВГС), адаптировались к подавлению функции MAVS в противовирусном врожденном иммунном ответе, способствуя репликации вируса. [7] [11] HCMV ослабляет MAVS через вирусный митохондриальный ингибитор белка апоптоза (vMIA), тем самым снижая провоспалительную цитокиновую реакцию. [11] vMIA также локализуется в пероксисоме, где взаимодействует с цитоплазматическим белком-шапероном Pex19, отключая транспортный механизм пероксисомальных мембранных белков. [11] Штамм HCV NS3-NS4A инактивирует передачу сигналов MAVS путем расщепления белка MAVS непосредственно перед доменом, нацеленным на мембрану MAVS, в MAM и пероксисоме, предотвращая передачу сигналов ниже по ходу MAVS. [7]
Регулирование
[ редактировать ]Экспрессия и функция MAVS регулируются на транскрипционном, посттранскрипционном и посттрансляционном уровне. На уровне транскрипции активные формы кислорода (АФК), образующиеся во время противовирусного ответа, действуют как негативный регулятор . [7] [8] [9] Кроме того, MAVS кодирует ряд вариантов сплайсинга, которые, как предполагается, регулируют MAVS. На посттранскрипционном уровне на MAVS присутствуют два сайта трансляции, которые могут генерировать два белка MAVS. Альтернативный сайт трансляции находится выше по течению, что приводит к экспрессии sMAVS. [7] [8] [9] На уровне трансляции белки, такие как семейство убиквитин E3-лигазы, регулируют активность MAVS. [7] [8] [9]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с GRCh38: Версия Ensembl 89: ENSG00000088888 – Ensembl , май 2017 г.
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000037523 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Васкес С., Beachboard DC, Хорнер С.М. (2017). «Методы визуализации субклеточной локализации MAVS». Врожденный противовирусный иммунитет . Методы молекулярной биологии. Том. 1656. стр. 131–142. дои : 10.1007/978-1-4939-7237-1_7 . ISBN 978-1-4939-7236-4 . ПМК 6103534 . ПМИД 28808966 .
- ^ Сет Р.Б., Сунь Л., Эа К.К., Чен З.Дж. (сентябрь 2005 г.). «Идентификация и характеристика MAVS, митохондриального противовирусного сигнального белка, который активирует NF-kappaB и IRF 3» . Клетка . 122 (5): 669–682. дои : 10.1016/j.cell.2005.08.012 . ПМИД 16125763 . S2CID 11104354 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v Васкес С., Хорнер С.М. (июль 2015 г.). «Координация противовирусного врожденного иммунитета MAVS» . Журнал вирусологии . 89 (14): 6974–6977. дои : 10.1128/JVI.01918-14 . ПМЦ 4473567 . ПМИД 25948741 .
- ^ Перейти обратно: а б с д и ж г час я дж к л Моханти А., Тивари-Пандей Р., Панди Н.Р. (сентябрь 2019 г.). «Митохондрии: незаменимые участники врожденного иммунитета и хранители воспалительной реакции» . Журнал сотовой связи и сигнализации . 13 (3): 303–318. дои : 10.1007/s12079-019-00507-9 . ПМК 6732146 . ПМИД 30719617 .
- ^ Перейти обратно: а б с д и ж г час я Джейкобс Дж.Л., Койн CB (декабрь 2013 г.). «Механизмы регуляции MAVS на митохондриальной мембране» . Журнал молекулярной биологии . 425 (24): 5009–5019. дои : 10.1016/j.jmb.2013.10.007 . ПМЦ 4562275 . ПМИД 24120683 .
- ^ Перейти обратно: а б с Цзян QX (2019). «Структурная изменчивость пути RLR-MAVS и чувствительное обнаружение вирусных РНК» . Медицинская химия . 15 (5): 443–458. дои : 10.2174/1573406415666181219101613 . ПМК 6858087 . ПМИД 30569868 .
- ^ Перейти обратно: а б с Эшли КЛ, Абендрот А, МакШарри Б.П., Слобедман Б (2019). «Интерферон-независимые врожденные ответы на цитомегаловирус» . Границы в иммунологии . 10 : 2751. дои : 10.3389/fimmu.2019.02751 . ПМЦ 6917592 . ПМИД 31921100 .
Дальнейшее чтение
[ редактировать ]- Нагасе Т., Исикава К., Кикуно Р., Хиросава М., Номура Н., Охара О. (октябрь 1999 г.). «Прогнозирование кодирующих последовательностей неопознанных генов человека. XV. Полные последовательности 100 новых клонов кДНК головного мозга, которые кодируют большие белки in vitro» . Исследование ДНК . 6 (5): 337–345. дои : 10.1093/dnares/6.5.337 . ПМИД 10574462 .
- Мацуда А., Сузуки Ю., Хонда Г., Мурамацу С., Мацузаки О., Нагано Ю. и др. (май 2003 г.). «Крупномасштабная идентификация и характеристика генов человека, которые активируют сигнальные пути NF-kappaB и MAPK». Онкоген . 22 (21): 3307–3318. дои : 10.1038/sj.onc.1206406 . ПМИД 12761501 . S2CID 38880905 .
- Каваи Т., Такахаши К., Сато С., Кобан С., Кумар Х., Като Х. и др. (октябрь 2005 г.). «IPS-1, адаптер, вызывающий индукцию интерферона I типа, опосредованную RIG-I и Mda5». Природная иммунология . 6 (10): 981–988. дои : 10.1038/ni1243 . ПМИД 16127453 . S2CID 31479259 .
- Мейлан Э., Карран Дж., Хофманн К., Морадпур Д., Биндер М., Бартеншлагер Р., Чопп Дж. (октябрь 2005 г.). «Кардиф представляет собой адаптерный белок противовирусного пути RIG-I, на который воздействует вирус гепатита С» (PDF) . Природа . 437 (7062): 1167–1172. Бибкод : 2005Natur.437.1167M . дои : 10.1038/nature04193 . ПМИД 16177806 . S2CID 4391603 .
- Ли XD, Сунь Л., Сет Р.Б., Пинеда Г., Чен З.Дж. (декабрь 2005 г.). «Протеаза вируса гепатита С NS3/4A отщепляет митохондриальный противовирусный сигнальный белок от митохондрий, чтобы избежать врожденного иммунитета» . Труды Национальной академии наук Соединенных Штатов Америки . 102 (49): 17717–17722. дои : 10.1073/pnas.0508531102 . ПМЦ 1308909 . ПМИД 16301520 .
- О Дж.Х., Ян Джо, Хан Ю, Ким М.Р., Бён С.С., Чон Ю.Дж. и др. (декабрь 2005 г.). «Транскриптомный анализ рака желудка человека». Геном млекопитающих . 16 (12): 942–954. дои : 10.1007/s00335-005-0075-2 . ПМИД 16341674 . S2CID 69278 .
- Лу Ю.М., Оуэн Д.М., Ли К., Эриксон А.К., Джонсон К.Л., Фиш П.М. и др. (апрель 2006 г.). «Вирусный и терапевтический контроль стимулятора промотора IFN-бета 1 при инфекции вируса гепатита С» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (15): 6001–6006. Бибкод : 2006PNAS..103.6001L . дои : 10.1073/pnas.0601523103 . ПМЦ 1458687 . ПМИД 16585524 .
- Ченг Дж., Чжун Дж., Чисари Ф.В. (май 2006 г.). «Ингибирование передачи сигналов, индуцированной дцРНК, в клетках, инфицированных вирусом гепатита С, с помощью протеаз-зависимых и независимых механизмов NS3» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (22): 8499–8504. Бибкод : 2006PNAS..103.8499C . дои : 10.1073/pnas.0602957103 . ПМЦ 1482521 . ПМИД 16707574 .
- Лин Р., Лакост Дж., Нахаи П., Сунь К., Ян Л., Пис С. и др. (июнь 2006 г.). «Диссоциация молекулярного комплекса MAVS/IPS-1/VISA/Cardif-IKKepsilon от внешней мембраны митохондрий путем протеолитического расщепления NS3-4A вируса гепатита С» . Журнал вирусологии . 80 (12): 6072–6 дои : 10.1128/JVI.02495-05 . ПМЦ 1472616 . ПМИД 16731946 .
- Саха С.К., Пьетрас Э.М., Хе Дж.К., Канг Дж.Р., Лю С.Ю., Оганесян Г. и др. (июль 2006 г.). «Регуляция противовирусных ответов путем прямого и специфического взаимодействия между TRAF3 и Cardif» . Журнал ЭМБО . 25 (14): 3257–3263. дои : 10.1038/sj.emboj.7601220 . ПМЦ 1523175 . ПМИД 16858409 .
- Босолей С.А., Виллен Дж., Гербер С.А., Раш Дж., Гиги С.П. (октябрь 2006 г.). «Вероятностный подход к высокопроизводительному анализу фосфорилирования белков и локализации сайтов». Природная биотехнология . 24 (10): 1285–1292. дои : 10.1038/nbt1240 . ПМИД 16964243 . S2CID 14294292 .
- Опиц Б., Винцинг М., ван Лаак В., Шмек Б., Хейне Г., Гюнтер С. и др. (ноябрь 2006 г.). «Legionella pneumophila индуцирует IFNbeta в эпителиальных клетках легких посредством IPS-1 и IRF3, которые также контролируют репликацию бактерий» . Журнал биологической химии . 281 (47): 36173–36179. дои : 10.1074/jbc.M604638200 . ПМИД 16984921 .
- Чен З., Бенюро Ю., Рейнбранд Р., Йи Дж., Ван Т., Вартер Л. и др. (январь 2007 г.). «Вирус GB B нарушает передачу сигналов RIG-I путем NS3/4A-опосредованного расщепления адаптерного белка MAVS» . Журнал вирусологии . 81 (2): 964–976. дои : 10.1128/JVI.02076-06 . ПМК 1797450 . ПМИД 17093192 .
- Хирата Ю., Броке А.Х., Менчен Л., Кагнофф М.Ф. (октябрь 2007 г.). «Активация механизмов врожденной иммунной защиты путем передачи сигналов через RIG-I/IPS-1 в эпителиальных клетках кишечника» . Журнал иммунологии . 179 (8): 5425–5432. дои : 10.4049/jimmunol.179.8.5425 . ПМИД 17911629 .
- Цзэн В., Сюй М., Лю С., Сунь Л., Чэнь З.Дж. (октябрь 2009 г.). «Ключевая роль полиубиквитинирования Ubc5 и лизина-63 в вирусной активации IRF3» . Молекулярная клетка . 36 (2): 315–325. doi : 10.1016/j.molcel.2009.09.037 . ПМЦ 2779157 . ПМИД 19854139 .
- Лю С., Чен Дж., Цай Икс, Ву Дж., Чен Икс, Ву Ю.Т. и др. (август 2013 г.). «MAVS рекрутирует несколько убиквитин-лигаз Е3 для активации противовирусных сигнальных каскадов» . электронная жизнь . 2 (е00785): е00785. doi : 10.7554/eLife.00785 . ПМЦ 3743401 . ПМИД 23951545 .
- Лю С., Цай Икс, Ву Дж, Конг Ц, Чен Икс, Ли Т и др. (март 2015 г.). «Фосфорилирование врожденных иммунных адаптерных белков MAVS, STING и TRIF индуцирует активацию IRF3» . Наука . 347 (6227): ааа2630. дои : 10.1126/science.aaa2630 . ПМИД 25636800 .
Эта статья включает текст из Национальной медицинской библиотеки США , который находится в свободном доступе .